
Abstract
Human papillomavirus (HPV) is associated with premalignant lesions such as high-grade cervical intraepithelial neoplasia (CIN-III) with potential progression to cervical carcinoma. There are now preventive vaccines against HPV. However, no effective therapeutic vaccine or immunological treatment exists for individuals already infected or for the 470,000 women that develop high-grade dysplasia, carcinoma in situ, and cervical cancer each year. More than half of these women die from cervical cancer. Relative non-immunogenicity of HPV infection is one of the main reasons for the difficulty in designing a comprehensive therapeutic vaccine against HPV-induced premalignant lesions and cervical carcinoma. HPV E6 and E7 proteins, the major HPV oncogenes, are highly immunogenic but fail to induce cross-reactive and protective immune responses against heterologous strains. We designed and synthesized a therapeutic peptide vaccine comprised of multivalent peptide mixtures called hypervariable epitope constructs (HECs) that represent the major epitope variants of the oncogenic E7 structural protein, and assessed their immunogenicity and in vivo efficacy in mice. Our results show that this peptide vaccine can induce strong, HPV-specific, T-helper cell and CTL responses. More significantly, we have demonstrated that the vaccine is efficacious as a therapeutic agent in a mouse HPV tumor model. Therefore, the HPV HEC vaccine approach described herein can potentially prevent progression of HPV-associated premalignant lesions, and may also be therapeutic against tumors associated with HPV.
Introduction
We report here the immunogenicity, prophylactic, and therapeutic effects of synthetic E7 peptide constructs in an HPV-induced murine transplantable tumor model. These peptide constructs are unique in that they are synthesized to represent the maximum intra-genotype variability of the E7 oncoprotein, and include several epitopes known to be recognized by human helper T cells. Our results show that such hypervariable epitope constructs (HECs), given with adjuvant, induce robust cellular and humoral immune responses that protect mice against HPV tumor cell challenge, and that are also able to temporarily reduce established HPV tumors. Furthermore, we demonstrate that lymphocytes from cervical cancer patients recognize the E7 HEC peptide constructs in vitro, suggesting that these epitopes are presented as tumor antigens in vivo.
Materials and Methods
Animals and tumor cell line
Female C57BL/6 mice, 6–8 wk of age, were purchased from Charles River Laboratories. The Animal Care and Use Committee of the University of California at Davis approved these animal experiments. Mice were housed under specific pathogen-free conditions. The TC-1 tumor cell line was a gift from Dr. T.C. Wu (John Hopkins Medical Institutions, Baltimore, MD). TC-1 cells are derived from mouse lung epithelial cells transformed to express HPV16 E6/E7 and activated c-Ha-ras oncogene. TC-1 cells were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum, 2 mM L-glutamine, and 50 μM 2-mercaptoethanol. Cells were grown at 37°C and 5% CO2.
HPV hypervariable epitope construct
The sequences of the HPV HECs are shown in Table 1. The procedure of synthesizing HECs has been described elsewhere (26,31). Briefly, the possible amino acids for each position along a variable epitope are determined from published sequence information of HPVs present globally. Subsequently, selected amino acid coupling steps in the synthesis of the epitope are performed with two amino acids chosen to represent the antigenic diversity determined from sequence data. Thus, in a single synthesis, an HEC consisting of a mixture of peptides representing the observed in vivo variants of the epitope is produced. HPV HECs were synthesized by 9-fluorenylmethyloxycarbonyl (Fmoc) chemistry, cleaved from the resin, and deprotected using trifluoroacetic acid (TFA) with EDT as scavenger, followed by extensive dialysis and lyophilization. Amino acid analysis was performed to ensure the appropriate amino acid contents, and the purity was analyzed by reverse-phase HPLC. Peptide stocks were prepared in 1×PBS for immunization experiments, and for ELISA and in RPMI 1640 media for in vitro cell assays.
Each HEC is based on an epitope in the E7 protein, recognized as human T-helper cell epitope, and expected to contain 64 variants. HPV HEC-1 also represents a murine cytotoxic T-cell epitope based on an H-2Kb background. Letters below each sequence indicate positions where two amino acids were added, and numbers indicate the percentage of each amino acid.
Immunogenicity studies
C57BL/6 mice (n=6) were vaccinated intradermally (ID) at the base of the tail with 100 μg of all three HPV HECs (equimolar mixture) in Montanide ISA-51 (Seppic Inc., Fairfield, NJ) four times on days 0, 12, 28, and 45. One week after the last immunization, the mice were euthanized and cellular immune assays were performed with cells harvested from the spleen and lymph nodes. The number of mice per experiment was chosen based on those used for several other HPV vaccines tested in animal models (13).
Therapeutic vaccination
Mice (n=12) were injected SC in the back with 2×105 TC-1 tumor cells in 200 μL of 1×PBS. Twelve days later, mice with tumors of palpable size (∼ 5 mm) were selected and randomly divided into two groups (n=6). Mice from group 1 were left unvaccinated (controls). Mice from group 2 were vaccinated ID with 100 μg of all three HPV HECs (equimolar mixture) in Montanide ISA-51 (Seppic Inc.), followed by boosters 12 and 28d later. Tumor development was monitored at least twice a week for 130 d.
Prophylactic vaccination
One week after the fourth immunization, mice (n=6) were challenged subcutaneously (SC) in the shaved back with 1×106 TC-1 tumor cells in 200 μL of 1×PBS, and tumor growth was monitored at least twice a week for a period of 150 d. TC-1 tumor cells were cultured in vitro, trypsinized, and washed three times with 1×PBS prior to being injected into mice.
HPV HEC-specific antibody response by ELISA
One week after the third and fourth immunizations, blood was collected and tested for HPV HEC-specific antibodies by ELISA. Ninety-six-well microtiter plates were coated with 10 μg/mL HPV HEC peptides per well in carbonate buffer (pH 9.6) overnight at 37°C. Non-specific binding sites were blocked with PBS/0.5% v/v Tween 20 (PBST) containing 3% milk powder. Sera were diluted at 1:1000, 1:5000, and 1:10000 in PBST containing 0.5% BSA, added to antigen-coated plates, and incubated 2 h at 37°C. After washing, HPV HEC-bound antibodies were detected using horseradish peroxidase-conjugated secondary antibody (rat anti-mouse IgG; Southern Biotechnology Inc., Birmingham, AL) at 1:1000 dilution in PBST containing 0.5% BSA. Color development was determined following the addition of SureBlue TMB microwell substrate (KPL). Absorbance was measured spectrophotometrically at 605 nm with an automatic plate reader (VERSAmax; Molecular Devices, Sunnyvale, CA) according to the manufacturer's instructions (SOFTmax PRO; Molecular Devices). Sera obtained before immunization, termed pre-bleed, was used as a negative control and compared with sera obtained after immunization. Two HPV-unrelated peptides derived from hepatitis C virus (HCV) and influenza viruses, as well as recombinant survivin, were used as negative controls. Sera were also tested against the buffer solution used to dissolve the peptides to exclude the possibility that the responses were caused by an artifact.
Ex vivo T-cell proliferation
Seven days after the final immunization, spleen and lymph nodes (inguinal, axillary, and mesenteric) were harvested. Then 3×105 cells from spleen and lymph nodes were cultured in round-bottom 96-well plates with HPV HECs at a final concentration of 1, 5, or 10 μg/mL in RPMI media supplemented with 10% FBS. HPV-unrelated peptides from influenza or hepatitis C virus were used as negative controls and ConA as positive control. After 48 h of incubation with peptide, 1 μCi [3H] thymidine (Amersham Biosciences, Piscataway, NJ) was added to each well. Following 18 h of culture, the plates were harvested using a PHD cell harvester, and the amount of tritiated thymidine incorporated into proliferated cells was assessed with a Beckman LS 6000IC scintillation counter. Results were expressed as counts per minute (CPM).
Cytotoxicity assay
Cytotoxic T cell (CTL) activity was determined using the JAM test that measures fragmentation of DNA associated with apoptosis (25). Each experiment was repeated three times. One week after the last vaccination, splenocytes and lymph nodes from vaccinated and control groups were harvested, pooled, and cultured with an equimolar mixture of the three HPV HEC peptides at a final concentration of 10 μg/mL and 100 IU/mL of recombinant murine IL-2 (R&D Systems, Minneapolis, MN) in 24-well tissue culture plates (4×106 cells per well) for 6 d to generate effector cells. [3H]-thymidine-labeled target cells were mixed with effector cells at various effector:target ratios. After 5 h of incubation at 37°C, cells were harvested using a PHD cell harvester, and radioactivity associated with fragmented DNA was assessed with a Beckman LS 6000IC scintillation counter. Results were expressed as CPM. Spontaneous release was determined by incubating target cells in medium alone. In this assay, reduction of counts when compared with TC-1 indicates a positive response. The amount of [3H]-thymidine in labeled TC-1 cells was used as an indicator of total CPM that remained in the target cells before incubation with effector cells. TC-1 tumor cells lysed by incubation with 5% Triton X-100 were used to determine maximum CPM.
ELISPOT for mouse cells
96-well MultiScreen HA sterile plates (Fisher Scientific, Pittsburgh, PA) were equilibrated with 1×PBS and coated with 10 μg/mL of anti-mouse IFN-γ capture antibody (eBioscience, San Diego, CA) in 100 μL of 1×PBS overnight at 4°C. Controls were processed in the absence of antibody. After blocking for 2 h with 100 μL of medium (RPMI and 10% FCS) at 37°C, freshly isolated splenocytes were added at 2.5×105 cells per well in triplicate, followed by serial dilutions in medium containing 10 μg/mL of an equimolar mixture of the three HPV HECs. HPV-irrelevant peptides were used as negative controls, and positive control wells received PMA/ionomycin (Sigma-Aldrich, St. Louis, MO), or mouse anti-CD3 (e-Bioscience) in 200 μL of complete medium. Cultures were incubated at 37°C in 5% CO2 for 24 h. Cells were removed by washing the plates six times with 1×PBS containing 0.05% Tween 20 and once with 1×PBS alone. Following the addition of 100 μL of 1×PBS containing 5 μg/mL of biotinylated anti-mouse IFN-γ antibody (eBioscience) per well, plates were incubated 2 h at 37°C. Wells were washed three times with 1×PBS and filled with 100 μL of avidin-horseradish peroxidase (1:1000 dilution; e-Bioscience) in 1×PBS. After standing 2 h at room temperature, the wells were washed three times with 1×PBS and developed for 30 to 60 min with 100 μL of freshly prepared 3-amino-9-ethyl carbazole (AEC) substrate (Sigma-Aldrich). The reaction was stopped by rinsing the wells with ice-cold water. Spots were quantified in an ELISPOT reader. Wells with media instead of splenocytes were run in parallel as background controls. Counts of negative control wells were subtracted from the samples. A sample was scored positive if it reached a value that was more than twice that of the control.
ELISPOT for human cells
Ninety-six well MultiScreen HA sterile plates were equilibrated with 1×PBS and coated with 10 μg/mL of anti-human IFN-γ capture antibody (eBioscience) in 100 μL of 1×PBS overnight at 4°C. Controls were processed in the absence of antibody. After blocking for 2 h with 100 μL of medium (RPMI and 10% FCS) at 37°C, human PBMCs previously thawed and conditioned were added at 500,000 cells per well in triplicate, followed by serial dilutions in medium containing 10 μg/mL of an equimolar mixture of the three HPV HECs. HPV-irrelevant peptides were used as negative controls, and positive control wells received PMA/ionomycin (Sigma-Aldrich) or human anti-CD3 (e-Bioscience) in 200 μL of complete medium. Cultures were incubated at 37°C in 5% CO2 for 48 h. Cells were removed by washing the plates six times with 1×PBS containing 0.05% Tween 20 and once with PBS alone. Following the addition of 100 μL of PBS containing 5 μg/mL of biotinylated anti-human IFN-γ antibody (eBioscience) per well, plates were incubated 2 h at 37°C. The wells were washed three times with 1×PBS and filled with 100 μL of avidin-horseradish peroxidase (1:1000 dilution; e-Bioscience) in PBS. After standing 2 h at room temperature, the wells were washed three times with 1×PBS and developed for 30 to 60 min with 100 μL of freshly prepared AEC substrate (Sigma-Aldrich). The reaction was stopped by rinsing the wells with ice-cold water. Spots were quantified in an ELISPOT reader (Zeiss-Vision Technologies, Hebron, KY). Wells with media instead of PBMCs were run in parallel as background controls. Counts of negative control wells were subtracted from the samples. A sample was scored positive if it reached a value that was more than twice that of the control.
Clinical samples
The clinical samples studied were from HPV-infected patients, cervical cancer patients, ovarian cancer patients, and young healthy individuals with no detectable HPV infection by PCR. The protocol was approved by the ethics committee of the Health Department of The State of Sonora, Mexico (protocol number CARES/81/09/05-CARES/SSP/01-04-09).
HPV genotyping
HPV testing was performed in samples from vaginal swabs by PCR using HPV-specific primers for HPV-16, -18, -31, and -45.
Statistical analysis
The significance of differences between groups was determined using a paired Student's t-test (with a confidence level of 95%) with GraphPad Prism software, version 5.01. Two-tailed p values of <0.05 were considered statistically significant.
Results
Design and construction of the three HPV HECs
Three HEC constructs representing three distinct regions of the E7 protein of HPV-16 were synthesized (Table 1). The sequences were selected based on previous studies showing that they contain T-helper and CTL epitopes.
Immunization with HPV HECs induces antigen-specific antibodies
Before performing survival trials of mice challenged with tumor cells before and after vaccination with HPV HECs, we studied the in vivo immunogenicity of HPV HECs in mice to determine whether they could elicit strong, antigen-specific humoral and cellular immune responses. HPV HEC-specific antibodies in mice immunized with HPV HECs were detected by ELISA (Fig. 1). Sera were collected after the third immunization with all three HPV HECs and tested individually against each HPV HEC to determine the HPV HEC-specific antibody response. Results showed that mice developed robust HPV HEC-specific antibody responses. Sera from mice immunized with HPV HECs showed a strong antibody response to the individual HPV HEC with which they were immunized, but not to a peptide unrelated to HPV HECs (FLU HEC). Interestingly, sera from mice immunized with HPV HECs also showed a strong antibody response to recombinant E7 protein. Pre-bleed sera obtained prior to immunization did not show antibody response to HPV HECs.

Induction of HPV HEC-specific antibodies. Sera were collected from mice immunized with an equimolar mixture of all three HPV HECs 7 d after the third immunization; 10 μg/mL of individual HPV HEC-1, -2, or -3, were used as coating antigens to detect HPV HEC-specific antibodies. Pre-bleed sera obtained from the same mice prior to immunization, and 1 μg/mL of recombinant E7 protein corresponding to HPV-16 (rE7) and 1 μg/mL of recombinant survivin were used as controls. The HPV-unrelated peptide, influenza HEC peptide (FLU-HEC), was also used as a negative control to demonstrate that the antibody responses elicited after immunization were HPV HEC-specific. Sera were diluted to 1:1000, 1:5000, and 1:10,000 and the plates were read for optical density (OD) at 605 nm. The results were expressed as mean OD values and standard deviation of triplicate wells.
HPV HECs induce antigen-specific T-cell proliferative responses
We also assessed whether HPV HECs could induce antigen-specific lymphoproliferation (Fig. 2A and B). The results show that all six individual mice immunized with HPV HECs developed strong HPV HEC-specific proliferative T-cell responses. Lymphocytes from spleens and lymph nodes in mice immunized with HPV HECs proliferated, reacting to individual HPV HEC, but there were no proliferative responses detected with HPV-unrelated peptides, such as FLU-HEC and HCV-HEC. Interestingly, the proliferative response to HPV HEC-2 was stronger than that to HPV HEC-1 or -3, a result similar to that observed in the detection of HPV HEC-specific antibody response in Fig. 1. Overall, these results suggest that the proliferative responses were HPV HEC-specific.
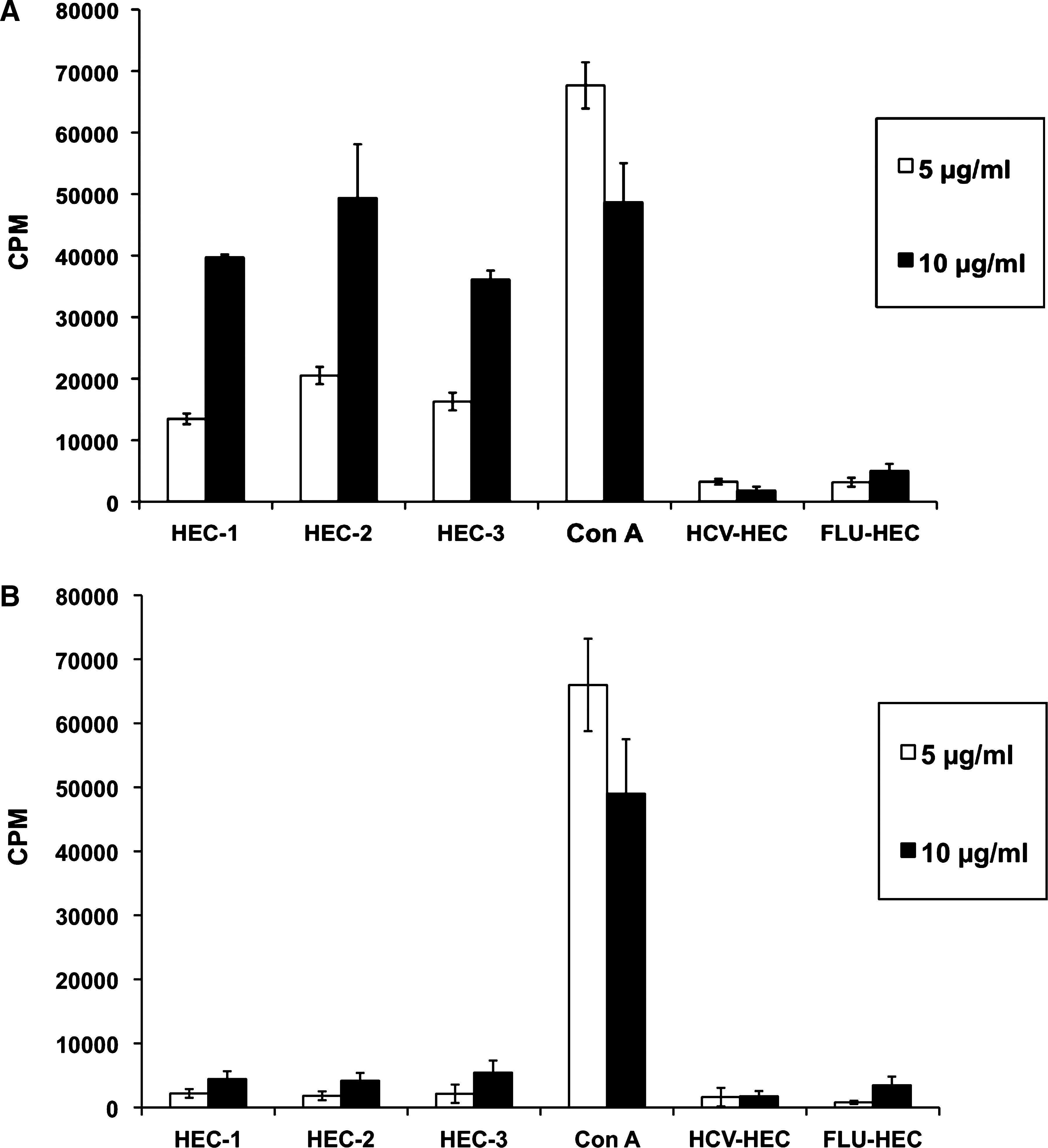
HPV HEC-specific proliferative T-cell responses. One week after the last immunization, spleens and lymph nodes from mice immunized with an equimolar mixture of all three HPV HECs were harvested. HPV-unrelated HECs based on a variable epitope present in the hemagglutinin of influenza virus (FLU-HEC) and hepatitis C virus (FLU-HEC) were used as negative controls, and ConA was used as positive control. 3×105 cells were re-stimulated with individual HPV HEC or HPV-unrelated peptides in a final concentration of 5 μg/mL (white bars) or 10 μg/mL (black bars). After 48 h of incubation, 1 μCi of [3H]-thymidine was added, and the cells were incubated further for 18 h. The amount of incorporated [3H]-thymidine into proliferating cells was expressed as counts per minute (CPM). Data shown in this bar graph represent the results from six individual mice with standard deviation (SD). Results with immunized (
HPV HECs induce specific T cells that secrete IFN-γ
We also assessed the production of interferon-γ (IFN-γ) by HPV HEC-specific memory T cells from mice immunized with HPV HECs by ELISPOT analysis (Fig. 3A and B). Splenocytes from mice immunized with HPV HECs were stimulated individually with HPV HEC-1, -2, or -3. In addition, splenocytes were also incubated with irradiated TC-1 tumor cells to assess whether HPV HEC-primed T cells could recognize HPV-16 E7 oncoprotein on TC-1 tumor cells, and secrete tumor-specific IFN-γ. Medium alone or HPV-unrelated HCV HEC peptides were used as negative controls.

ELISPOT detection of IFN-γ production by HPV-specific T cells. Spleen cells were harvested from C57BL/6 mice (8 per group) immunized four times with HPV HEC-1, -2, and -3 (an equimolar mixture of the 3 HECs). One week after the final immunization, 10 μg/mL of HPV HEC-1, -2, and -3, and irradiated TC-1 cells were added in triplicate to each well pre-coated with anti-mouse IFN-γ, and incubated for 24 h. Cells containing medium only were used as background, HCV HEC as negative control, and PMA/ionomycin as positive control. Results with immunized (
Our results showed that HPV HECs elicited IFN-γ-secreting, HPV HEC-specific T cells in mice immunized with HPV HECs. Moreover, splenocytes from mice immunized with HPV HECs also secreted IFN-γ upon culture with TC-1 tumor cells. The number of IFN-γ spots produced by splenocytes stimulated with TC-1 tumor cells was comparable to that from cells stimulated with HPV HEC-1 or -3. Furthermore, the number of cells secreting IFN-γ in response to HPV HEC-2 was higher than that to other HPV HECs or TC-1 cells, and was comparable to the PMA/ionomycin used as positive control.
HPV HECs induce CTLs that kill tumor cells
Next we assessed whether HPV HECs could induce CTLs that were cytotoxic to tumor cells. When cells from mice immunized with an equimolar mixture of HPV HECs were tested against TC-1 tumor cells, generation of HPV HEC-specific CTLs was demonstrated, as shown in Fig. 4. CTLs from mice immunized with HPV HECs recognized TC-1 cells as targets, and had significant cytotoxicity against these tumor cells. When the E:T ratio was 50:1, the cytotoxic responses of HPV HEC-specific CTLs were as strong as that of the positive control. Moreover, about 25% of killing activity of CTLs induced by HPV HECs was observed at the lowest E:T ratio of 6:1. These data suggest that HPV HECs induce CTLs in mice that are HPV-specific and have strong cytolytic activity against TC-1 tumor target cells. The experiment was performed three times.
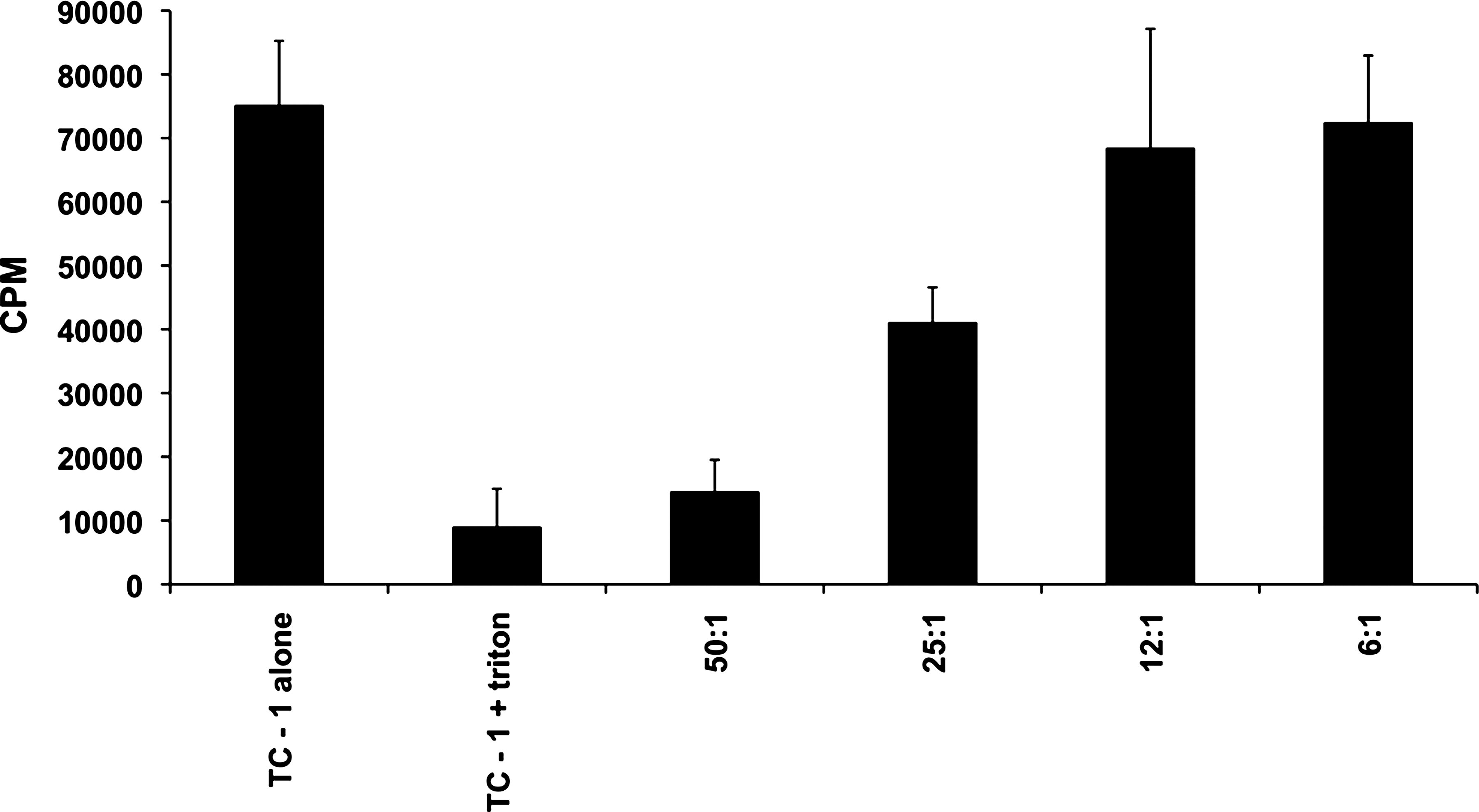
CTL responses in HPV HEC-immunized mice. One week after the last immunization, splenocytes and lymph nodes were harvested and a CTL assay was performed in triplicate using the JAM test. Results were derived from pooled lymph node and spleen cells cultured at a final concentration of 10 μg/mL of a mixture of three HPV HECs and IL-2 (500 IU/mL) for 6 d to generate effector killer cells. HPV HEC-specific effector cells were added to [3H]-thymidine-labeled (5 μCi/mL) TC-1 tumor cells at different ratios of effectors to targets: 50:1, 25:1, 12:1, and 6:1. The amount of [3H]-thymidine from live target cells that were not killed by CTLs was measured by the JAM test to determine the cytotoxicity of HPV HEC-specific CTLs. The amount of [3H]-thymidine in labeled TC-1 cells alone was used as an indicator of the total CPM that remained in target cells before incubation with effector cells. TC-1 tumor cells incubated with 5% Triton X-100 were used as positive control for maximum lysis.
Tumor-bearing mice can recognize HPV HECs as antigens
Our in vivo immunogenicity study showed that HPV HECs are strong immunogens that can elicit HPV HEC-specific humoral and cellular immune responses. Next we wanted to determine whether natural tumor-specific immune responses generated in tumor-bearing mice could recognize HPV HECs. C57BL/6 mice (n=6) were injected with 1×106 TC-1 tumor cells. After 21 d, all the mice developed a tumor larger than 1.5 cm in diameter. The spleens were harvested, and total splenocytes (3×105 cells per well) were cultured in vitro with 10 μg/mL HPV HEC or a mixture of HPV HEC-1, -2, and -3. Media alone or HPV HEC-unrelated peptides (HCV-HEC) were used as negative controls.
The proliferative responses of lymphocytes from HPV-16 E7-positive tumor-bearing mice were specific for HPV HECs, showing negative responses to HPV HEC-unrelated peptide (HCV-HEC), and to media only. This was an interesting result, in that immune responses generated against TC-1 tumors had reactivity against HPV HECs in vitro. These results suggest that even under immunosuppressive conditions, there were spontaneous T cells able to react with the synthetic HPV constructs. This may represent spontaneous but irrelevant immune responses due to the limitations of the model, and/or that the large number of tumor cells transplanted did not stop tumor growth. It is possible that these spontaneous T cells potentially represent the capability of the peptides to be presented in vivo. The data suggest that the HPV HECs represented the epitopes in the tumor. In addition, the lymphoproliferative response to HPV HEC-2 from TC-1 tumor-bearing mice was the strongest of the three HPV HECs tested (Fig. 5). These results are consistent with the experiments showing that the in vivo immunogenicity of HPV HEC-2 was the strongest in terms of antibody response, lymphoproliferative response, and IFN-γ detection by HPV HEC-specific T cells. These results suggest that HPV HEC-2 could be the strongest immunogenic CD4+ T-cell epitope.

Splenocytes from tumor-bearing mice recognize the HPV HECs in vitro. Mice (n=6) were injected with 1×106 TC-1 tumor cells. Spleens were harvested from mice that developed a tumor larger than 1.5 cm in diameter after 21 d. 3×105 splenocytes were incubated in vitro with individual HPV HECs or an equimolar mixture of all three HPV HECs at a final concentration of 10 μg/mL. Proliferative responses specific to HPV HECs were assessed by measuring the amount of [3H]-thymidine incorporated by cells during proliferation. The results were expressed as mean counts per minutes (CPM) with standard deviation in triplicate.
Lymphocytes from patients with cervical carcinoma in situ and CIN 2/3 recognize the HPV HECs
We also examined a repository of PBMC samples from HPV-positive patients with pre-malignant lesions (CIN 2/3) and cervical carcinoma to determine whether HPV HECs could be recognized by cells from this group of patients (Fig. 6). The samples were collected just before initiation of standard of care treatment. Repository samples from HPV-negative patients, and from patients with ovarian carcinoma (HPV-negative), were used as controls. HPV HEC-2, the most immunogenic peptide based on our in vivo immunogenicity study of HPV HECs in mice, was tested in vitro to assess whether HPV HEC-2 stimulation could elicit IFN-γ secretion by lymphocytes from these patients. Our results show that lymphocytes from patients with pre-malignant lesions (CIN 2/3) and cervical carcinoma secreted IFN-γ upon stimulation with HPV HEC-2, suggesting that HPV-positive patients from both groups can recognize HPV HEC-2 as antigen presented in vivo.
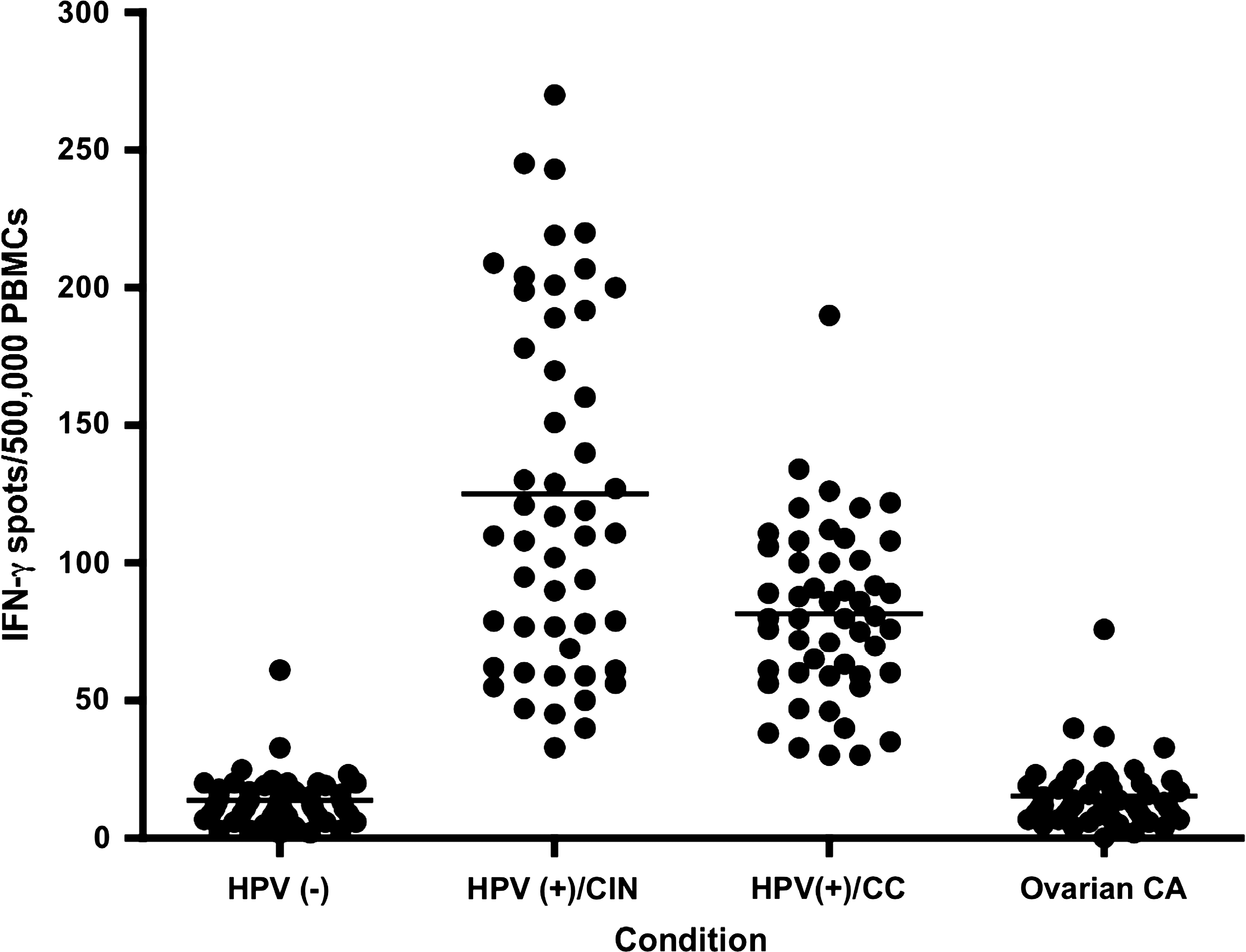
IFN-γ secretion by lymphocytes from HPV-infected patients incubated with HPV HEC-2. Cells collected just before initiation of standard of care treatment from HPV-16/HPV-18-positive patients with CIN 2/3 [HPV(+)/CIN], or cervical carcinoma [HPV(+)/CC] recognize the most immunogenic HPV HEC peptide, HEC-2. Cells from 50 patients were tested by ELISPOT. The ovarian cancer (Ovarian CA) patients selected were negative for HPV.
HPV HECs can be effective as a therapeutic vaccine
To determine the in vivo efficacy of HPV HECs as a therapeutic vaccine, the transplantable TC-1 tumor cell line expressing HPV E7 was used to challenge mice before vaccination (Fig. 7). Mice started developing tumors that were palpable 12 d after injection. Mice were randomly divided into two groups (6 mice per group). In the group of mice that was vaccinated after tumor challenge, tumor growth was stabilized and slowed down, never growing more than 0.5 cm in size. Furthermore, starting around day 60 post-inoculation, tumor regression was observed, and mice that received the therapeutic HPV HEC vaccine became tumor-free at about 105 d after challenge. In contrast to mice that received therapeutic HPV HEC vaccination, unvaccinated mice had to be sacrificed 20 d after challenge with tumor cells due to large tumor growth.
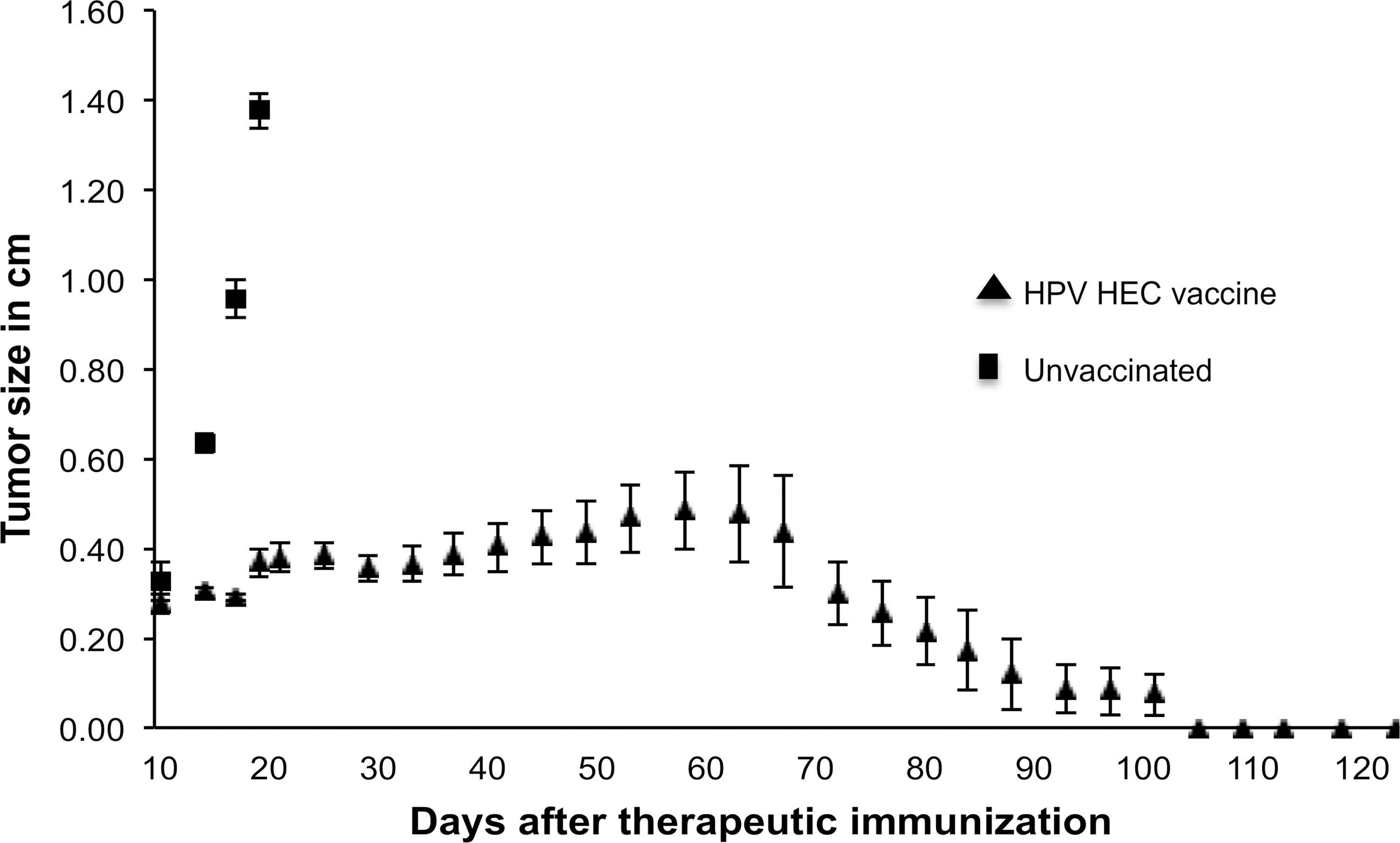
Therapeutic effect of the HPV HEC vaccine. Eight- to 10-wk-old female mice (n=12) were injected SC with 2×105 TC-1 tumor cells. Palpable tumors were detected 12 d later, and mice were divided into two groups. Mice in the experimental group (triangles) were vaccinated with 100 μg of an equimolar mixture of all three HPV HECs, followed by boosters with the same amount 12 and 28 d later. Control group mice (squares) were left unvaccinated. Tumor growth of mice from tumor challenge were monitored for up to 130 d. Data are presented as the mean of 6 mice per group.
HPV HECs can be effective as a prophylactic vaccine
After demonstrating the therapeutic activity of the HPV HEC construct, we determined the efficacy of HPV HEC as a prophylactic vaccine (Fig. 8). Seven days after the last immunization, mice were challenged subcutaneously with 1×106 TC-1 tumor cells. Tumor growth and mouse survival were monitored for 150 d. Unvaccinated mice of the same age as the vaccinated group were used as controls. All mice in the control group developed progressive tumors and were eventually sacrificed before 30 d due to tumor growth beyond the size restriction of the approved protocol. Remarkably, all the animals vaccinated with HPV HECs were protected from tumors up to 65 d, remaining tumor-free. Although some mice eventually developed delayed tumor formation starting at 70 d after challenge, this was only seen in 25% of the vaccinated animals. Moreover, more than 60% of mice remained tumor-free until the end of the prophylactic study (150 d).
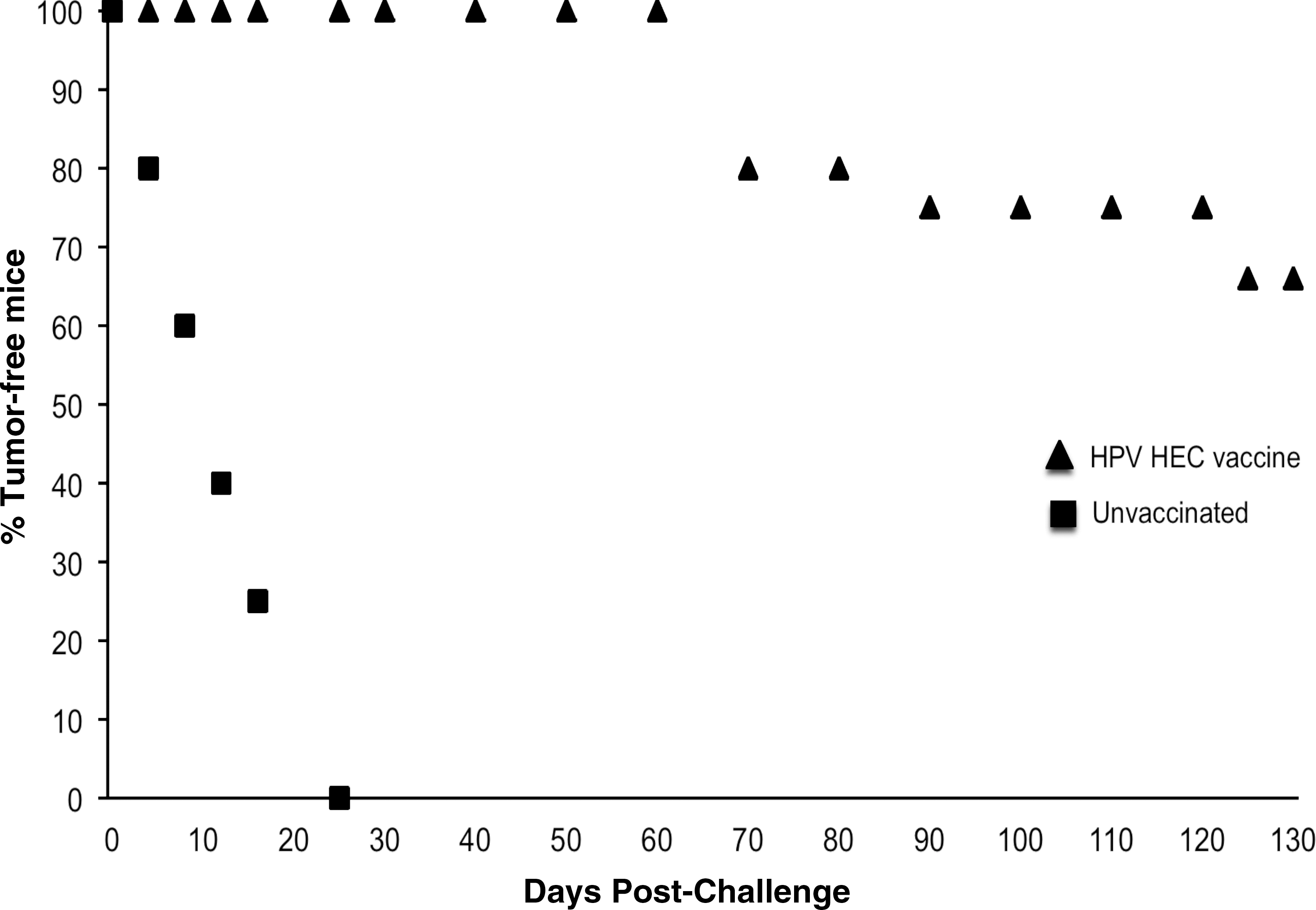
Preventive immunization induces tumor stabilization and regression. Mice were vaccinated ID with 100 μg of an equimolar mixture of all three HPV HECs four times at days 0, 12, 28, and 45. Seven days after the last immunization, the mice were challenged SC with 1×106 TC-1 tumor cells. Tumor growth and survival of mice were monitored for 150 d. Unvaccinated mice of the same age (squares) were also challenged with the same number of tumor cells and used as a control group. A significant number of the mice remained tumor free for more than 130 d. In 25% of the animals, tumors re-appeared after 70 days (triangles). Data are presented as the mean of 6 mice per group.
Discussion
In this report, we have shown that an immunogen based on three antigenically variable epitopes of the E7 protein of HPV can be used both therapeutically and preventively against challenge with tumor cells expressing E7. These results demonstrate that HPV HECs are strong immunogens inducing antibodies and T cells that are specific to tumors expressing HPV E7 protein, and importantly that these peptides are recognized by PBMCs from cervical carcinoma patients.
Multiple HPV immunotherapy clinical trials have involved immunization with virus-like particles (VLPs), DNA, peptides, and adoptive therapy with dendritic cells (DCs) pulsed with peptides (14,15,19,21,37,38,40). Most have been Phase I trials showing safety of the vaccine formulation (12,37,38,40,41). Knowledge gained from clinical trials performed to date indicates that neutralizing antibodies and/or E6/E7-specific CTLs can be produced in healthy subjects, and to an appreciable extent in patients with cervical dysplasia (9,16,34,39).
The role of cellular immune responses mediated by CTLs in protection against progression of CIN2/3 to carcinoma in situ has been extensively examined (16,17,20). It is clear that HPV-16-specific CTLs can be detected in the blood of these patients (32,34,40,41).
Relatively low immunogenicity and intra-strain variability complicate the development of a broadly protective and effective therapeutic vaccine against pre-malignant lesions and cervical carcinoma associated with the main types and subtypes of HPV (35,37). A hallmark of the HPV virus is its extensive intra-strain antigenic variation (2,3,6). Although HPV antigenic epitopes show sequence variation, we have observed that the sequence diversity of these epitopes appears to be limited and partially predictable (16,18,21). To exploit this pattern of diversity, we have developed a procedure that uses computer-aided epitope variation analysis and peptide synthesis to produce a peptide mixture representing the known amino acid sequences of an antigenic epitope (26,31). This approach of representing the known antigenic variant sequences of an epitope was developed at our laboratory, and the synthetic immunogen was named hypervariable epitope construct (HEC) (26). These immunogens are then validated for antigenicity with a collection of sera obtained from affected individuals, such as those bearing CIN 2/3, carcinoma in situ, and cervical carcinoma (31).
The experiments reported here included an immunogenicity study of HPV HECs, anti-tumor T-cell recognition of tumor cells expressing E7, in vivo efficacy of HPV HECs for prevention of tumor establishment, and as therapy of existing tumors and the assessment of the spontaneous immune response in cervical cancer patients.
The immunogenicity studies demonstrated that HPV HECs are capable of inducing HPV HEC-specific humoral and cellular immune responses. Antibodies generated in mice strongly bound only to HPV HECs and recombinant E7 and did not show cross-reactivity with other peptides and proteins unrelated to HPV (Fig. 1). The data suggest that HPV HECs induce B cells that produce abundant antibodies capable of recognizing HPV HECs and the recombinant HPV-16 E7 protein with high affinity. In a future study, it would be interesting to learn how long HPV HEC-specific antibodies would persist after the final immunization.
The cellular immune responses induced by HPV HEC in mice were quite interesting. Proliferation responses and IFN-γ secretion of T cells were significantly strong and specific to HPV HECs and recombinant E7 protein (Fig. 2). The results presented here suggest that robust proliferative T-cell responses can also be induced in mice immunized with HPV HECs. As this lymphoproliferation assay was performed with unfractionated lymphocytes, it is also possible that antigen-presenting DCs that process HPV HECs could cross-present to CD8+ T cells and induce HPV HEC-specific CD8+ T-cell proliferative responses. Further studies are required to explore this possibility.
It was surprising to detect IFN-γ secretion when T cells from HPV HEC-immunized mice were incubated with tumor cells (Fig. 3A). In addition, mice injected with tumor cells developed strong spontaneous proliferative splenocyte responses to HPV HECs. These results suggest that although the natural immune response only represents the pool of T cells generated against the tumor cells, there were also memory cells able to recognize the HEC peptides (Fig. 5). In fact, there are several reports concerning the study of the natural/spontaneous immune response in cancer-bearing hosts, even under the high tumor burdens seen when the pool of cells that were generated against the tumor eventually became irrelevant due the multiple immunosuppressive networks (29,36).
These responses were specific for HPV HECs, and no significant responses to HPV-irrelevant peptides or proteins were observed. These results could indicate that the peptides from HPV HECs were intracellularly processed, and that they include sequences that are very similar to the naturally-processed epitopes derived from tumor cells. These data suggest that HPV HECs represent important antigenic epitopes of E7 found in HPV tumors, and are strong immunogens that induce antigen-specific cellular responses.
It was significant to determine that the ex vivo-generated effector killer cells from mice immunized with HPV HECs had strong cytotoxic activity against TC-1 tumor target cells (Fig. 4). Results from the JAM test suggest that HPV HECs can induce robust CTLs against TC-1 tumor cells in vivo in mice.
Based on these positive results, we wanted to determine if tumor-specific naturally-occurring immune responses generated in humans with CIN 2/3 and cervical carcinoma and in HPV tumor-bearing mice could recognize the most immunogenic HPV HEC (HEC-2).
Significantly, lymphocytes from cervical cancer patients and CIN 2/3 patients secrete IFN-γ when incubated with these immunogens (Fig. 6). These data may suggest that some of these peptides are naturally processed in vivo, and may represent good candidates for future human studies using HEC peptides as potential therapeutic vaccines.
The proliferative response to HPV HEC-2 in tumor-bearing mice was the strongest among the three HPV HECs tested, and this correlates with our findings with the clinical samples. This is consistent with the results obtained when studying the immunogenicity of HPV HECs, showing that proliferative responses to HPV HEC-2 were stronger than to the other two HECs. It is reasonable to conclude that HPV HEC-2 represents the strongest immunogenic peptide. These results suggest that the sequences represented by HPV HECs are processed in vivo and presented to the immune system as natural epitopes.
Mice were protected from tumors when they were vaccinated with HPV HECs before (Fig. 7) and after tumor challenge (Fig. 8). Not only were mice able to survive tumor challenge, but the size of established tumors was also reduced.
Collectively, these data suggest that HPV HECs, a mixture of peptides designed by our laboratory based on the antigenic variation of the E7 oncoprotein, turned out to be both an effective preventive and therapeutic vaccine, with the potential to act against pre-malignant lesions and tumors. The vaccine is inexpensive to produce and very stable, and can be stored in dry powder form (lyophilized) at room temperature for use in developing countries without the need for cold chain storage and distribution.
Footnotes
Acknowledgments
We thank Carmelo R. Peraza and Dylan Dodd for expert technical work.
Author Disclosure Statement
No competing financial interests exist.