
Abstract
Interleukin-21 (IL-21) is a T-cell-derived cytokine that modulates T-cell, B-cell, and natural killer cell responses. It is not known if it could be used as an adjuvant for HIV DNA vaccination. In our study, we investigated if a DNA construct expressing IL-21 (designated as pVAX-IL-21) as a molecule adjuvant could enhance antigen-specific immune responses to an HIV DNA vaccine (pGX-EnvC). We found that a higher level of antigen-specific cytotoxic responses was induced in BALB/C mice immunized with pGX-EnvC with the pVAX-IL-21 via electroporation. The increased response was associated with higher expression of IFN-γ in CD8+ T cells. In contrast, the administration of pVAX-IL-21 inhibited the antibody responses to HIV induced by the pGX-EnvC. The plasma cell inhibitory transcription factors B-cell lymphoma 6 protein (Bcl-6) and Pax-5 were increased in B cells from mice that had been immunized by HIV DNA vaccine plus pVAX-IL-21, suggesting that the expressed IL-21 may inhibit the differentiation from B cells to plasma cells. These results indicate that IL-21 could enhance CD8+ T-cell immunity, but inhibit humoral responses during HIV DNA vaccination.
Introduction
IL-21 is produced mainly by activated CD4+ T cells, natural killer T cells, and follicular helper T cells, and acts upon cells from a range of lymphohematopoietic lineages. IL-21 serves a critical role in terminal B-cell differentiation and immunoglobulin production. It also acts as a T-cell co-mitogen to drive the expansion of CD8+ T cells, and plays an essential role in the differentiation of Th-17 cells (7,8). Although most of the effects of IL-21 on lymphoid cells are stimulatory, involving enhanced proliferation or effector function, the effects of IL-21 on dendritic cells (DCs) are inhibitory for DC maturation and function (9,10). In addition, IL-21 can be potently pro-apoptotic for B cells in a context-dependent manner (10 –12). Therefore, whether IL-21 has adjuvant effects to enhance HIV DNA vaccine responses remains unclear.
In our study, we directly examined whether IL-21 can be used as an adjuvant to enhance immune responses to a DNA vaccine encoding the outer membrane proteins of HIV. Although it did enhance antigen-specific cytotoxic responses, we found that co-inoculation with an IL-21 expression construct with the HIV DNA vaccine also inhibited the humoral response by decreasing the number of plasma cells and increasing the expression of B-cell lymphoma 6 protein (Bcl-6) and Pax-5 in B cells.
Materials and Methods
Reagents and animals
HIV envelope peptides representing T-cell epitopes mTh1 (VYYGVPVWKEAKTTLFC), mTh2 (TSAITQACPKVSFDPIPIHYCAPAG), mCTL1 (RYLKDQQLL), mCTL2 (WYIKIFIMI), mCTL3/Th3 (TSAITQACPKVSFDPIPIHYCAPAG), and neutralizing antibody epitopes mB1 (PNNNTRKSIRIGPGQTFYAT), mB2 (QKVYALFYRLD), and mB3 (ENFDMWKNDM) were synthesized by Science Peptide Co., Ltd. (Shanghai, China). FITC-, APC-, and PE-conjugated antibodies (Abs) used for flow cytometry analysis were purchased from BD Pharmingen (San Diego, CA). Adult female BALB/C mice at 8–10 wk of age were purchased from the Animal Institute of the Chinese Medical Academy (Beijing, China), and supplied with pathogen-free food and water in a 12-h light-dark cycle.
Plasmid construction and preparation
The pGX-EnvC plasmid was made in Dr. Weiner's lab at University of Pennsylvania. Seven pairs of mouse IL-21-specific primers were designed as listed in Table 1. Then the coding sequence for mouse IL-21 was obtained by the overlapping PCR method, and subcloned into the pVAX vector to create pVAX-IL-21. Plasmids were maxi-prepared by the alkaline method, subsequently purified with a Qiagen Maxi prep kit (Qiagen Inc., Duesseldorf, Germany), and diluted in saline.
Transfection of the BHK cell lines
The purified plasmids pVAX-IL-21 were transfected into BHK cells with lipofectamine according to the manufacturer's instructions (Invitrogen, Carlsbad, CA). The transfected cells were harvested after 48 h and blocked with Fc-Block (BD Pharmingen) in PBS for 30 min at 4°C, before being fixed with 4% paraformaldehyde and permeabilized with saponin. The cells were intracellularly stained with anti-IL-21-PE (BD Pharmingen) for 1 h at 4°C, and analyzed with a FACSCalibur using Cell Quest ProSoftware (BD Biosciences, San Jose, CA).
Immunization
The BALB/C mice were randomly divided into five groups (6 per group), and immunized intramuscularly with 30 μg plasmid or 30 μL PBS as control via electroporation (Inovio Inc., Blue Bell, PA) on days 0, 14, and 28, as listed in Table 2.
Detection of anti-Env-specific antibody
The detection of anti-EnvC-specific antibodies in the sera was carried out by quantitative ELISA assays. The wells were coated with 2 μg/mL of HIV-1 envelope peptide at 4°C overnight at 100 μL per well. These wells were subsequently blocked with 5% of BSA-PBST for 1 h at 37°C, and reacted with serial dilutions of mice sera (in 10-fold increments). After three washes, 1:1000 diluted goat anti-mouse IgG, IgG1, and IgG2a conjugated with horseradish peroxidase was added to the wells, and the plates were incubated at 37°C for 1 h. To develop the ELISA result, a 10-mg TMB tablet was dissolved in 0.025 M phosphate-citrate buffer, and 50 μL of the resulting solution was added to each well. The reaction was stopped by the addition of 2 M of H2SO4, and the resulting optical density (OD) at 450/620 nm was analyzed with a plate reader (Bio-Rad, Hercules, CA). The OD ratios of the same dilutions in immunized mice and naive mice were calculated. The titer of anti-EnvC antibody from the immunized mice was determined by the maximum dilution with a ratio greater than or equal to 2. The serum samples from immunized mice were tested individually on day 7 after the third immunization, and the antibody concentrations were calculated as the means of three independent assays.
Cell proliferation
Single-lymphocyte suspensions were obtained from spleens of the mice on day 7 after the third immunization. Cells in RPMI-1640 medium (Gibco, Eggenstein, Germany)/10% FBS were used to assay antigen-specific cell proliferation by the MTT method after HIV envelope peptide stimulation in vitro for 48 h as previously described (13). The OD values were read at 490 nm with a plate reader (Magellan; Tecan, Männedorf, Switzerland). Data were expressed as stimulation index (SI), calculated as the mean reading of triplicate wells stimulated with an antigen, divided by the mean reading of triplicate wells stimulated with medium alone.
In vivo cytotoxicity assay
Cytotoxicity in vivo was assayed as described previously (14). Splenocytes from naive BALB/C mice were pulsed with 10−6M EnvC peptide and labeled with a high concentration of CFSE (15 μm, CFSE high cells) as target cells. A portion of the same splenocytes was labeled with a low concentration of CFSE (0.5 μM, CFSE low cells) without peptide pulse as a nontarget control. The target and control cells were mixed in a 1:1 ratio, and injected into immunized mice at 2×107 total cells per mouse via the tail vein on day 7 after the third immunization. Four hours later, the spleens of injected mice were removed and the target and control cells were analyzed by their differential CFSE fluorescence intensities using a FACSCalibur. Specific lysis was calculated using the following formula: ratio=percentage CFSE low/percentage CFSE high. Percentage specific lysis=[1 – (ratio unprimed/ratio primed)×100].
Flow cytometric analysis
Splenic cells were isolated on day 7 after the third immunization. T cells or CD8+ T cells from splenocytes of BALB/C mice were isolated by MACS sorting (R&D Systems, Inc., Huntingdon Valley, PA), adjusted to 0.5×106 cells/20 μL, and stimulated in 96-well plates with EnvC peptide (5 μg/mL), and anti-CD28 (5 μg/mL) mAbs for 6 h at 37°C and 5% CO2. Monensin (5 μg/mL) was added for the last 4 h, and the cells were washed three times with PBS/10% FCS. The cells were blocked with Fc-Block (BD Phamingen) in PBS for 30 min at 4°C before being fixed with 4% paraformaldehyde and permeabilized with saponin, then were immunostained with isotype controls, or double-stained with anti-CD8-FITC and anti-IFN-γ-PE, or anti-CD4-FITC and anti-IFN-γ-PE, or anti-CD4-FITC and anti-IL-4-PE, for 1 h at 4°C. The cells were washed and analyzed with a FACSCalibur using Cell Quest Pro Software.
Follicular helper T cells and plasma cell analysis
T cells or B cells were isolated from spleen on day 7 after the third immunization, and adjusted to 0.5×106 cells/20 μL. Cells were blocked with Fc-Block (BD Phamingen) in PBS for 30 min at 4°C, immunostained with isotype controls, or double-stained with anti-CD4-FITC and anti-CXCR5-APC, or anti-CD138-PE and anti-B220-APC, for 30 min at 4°C. The cells were washed and analyzed with a FACSCalibur using Cell Quest Pro Software. The percentage of follicular helper T cells (TFH) and plasma cells was set as the percentage of CD4 and CXCR5, or B220 and CD138 double-positive cells.
Quantitative PCR
Seven days after the third immunization, B cells from splenocytes of BALB/C mice were isolated by MACS sorting. Total RNA was extracted from sorted B cells, and then was reverse transcribed. The sequences of the primers are listed in Table 3. Quantitative PCR was performed to amplify the Pax-5 and Bcl-6 genes using a SYBR® PrimeScript® RT-PCR Kit (Takara Inc., Otsu, Japan). The reaction was run on an ABI 7500, and data analysis was performed using 7500 software v 2.0.
Statistical analysis
Results are presented as means±standard error of the mean (SEM). Student's t-test analysis was used for data analysis. A value of p<0.05 was considered statistically significant.
Results
Cloning of IL-21 and expression in BHK cells
In order to construct the IL-21 expression plasmid pVAX-IL-21, we designed seven pairs of mouse IL-21 primers, and the full-length (441 bp) IL-21 coding sequence was cloned by overlap PCR, verified by sequencing, and then subcloned into the pVAX vector for eukaryotic expression (Fig. 1A). To validate the plasmid, we digested it with HindIII and XbaI to reveal the IL-21 fragment (Fig. 1B). To confirm pVAX-IL-21 protein expression, we transfected BHK cells, and these cells were stained intracellularly with anti-IL-21-PE 48 h after transfection. Expression was observed as shown in Fig. 1C.

Cloning and expression of IL-21. (
IL-21 as a molecular adjuvant enhances cytotoxicity responses
Since the antigen-specific cytotoxic response had been previously demonstrated to be important for protection against or eradication of viruses or other intracellular pathogens, we first tested this cellular response in vivo. BALB/C mice were immunized intramuscularly with pVAX-IL-21 or with vector alone, and the specific cytotoxic response was detected in vivo on day 7 after the third immunization. As shown in Fig. 2A, and the mean of three experiments shown in Fig. 2B, mice immunized with pGX-EnvC plus pVAX-IL-21 had an augmented HIV-EnvC-specific cytotoxic response compared to pGX-EnvC plus pVAX. The result demonstrated that IL-21 could act as an adjuvant and enhance the HIV-EnvC-specific cytotoxic response to HIV DNA vaccine.

Effects of IL-21 on in vivo cytotoxic responses. (
Effects of IL-21 on expression of cytokines in T cells
To determine which subset of T cells was involved in enhancing the cytotoxic response, we analyzed the expression of cytokines in T cells. Total splenic cells were isolated on day 7 after the third immunization, and re-stimulated in culture with the HIV-EnvC peptide. These cells were then analyzed by the FACS with gates set for CD4+ and CD8+ T cells. From the results shown in Fig. 3A, and the mean of three independent experiments shown in Fig. 3B, we observe that pGX-EnvC plus pVAX-IL-21 induced a higher level of IFN-γ expression by antigen-specific CD8+ T cells than did pGX-EnvC alone, whereas the percentage of CD4+ T cells stained for IFN-γ was not changed compared with pGX-EnvC alone. These results suggest that IL-21 can stimulate the response of CD8+ effector cells.

Effects of IL-21 on the expression of cytokines in T cells. (
IL-21 as adjuvant inhibits humoral responses
To examine the effect of adjuvant on the humoral responses in mice, serum was collected on day 7 after the third immunization, serum total IgG, IgG2a, and IgG1 antibodies against HIV-EnvC were determined by quantitative ELISA. Contrary to our expectations, lower levels of total IgG, IgG2a, and IgG1, were elicited in mice vaccinated with pGX-EnvC plus pVAX-IL-21, compared to the control group immunized with pGX-EnvC alone (Fig. 4A, B, and C). Hence, IL-21 as adjuvant could inhibit the humoral response.
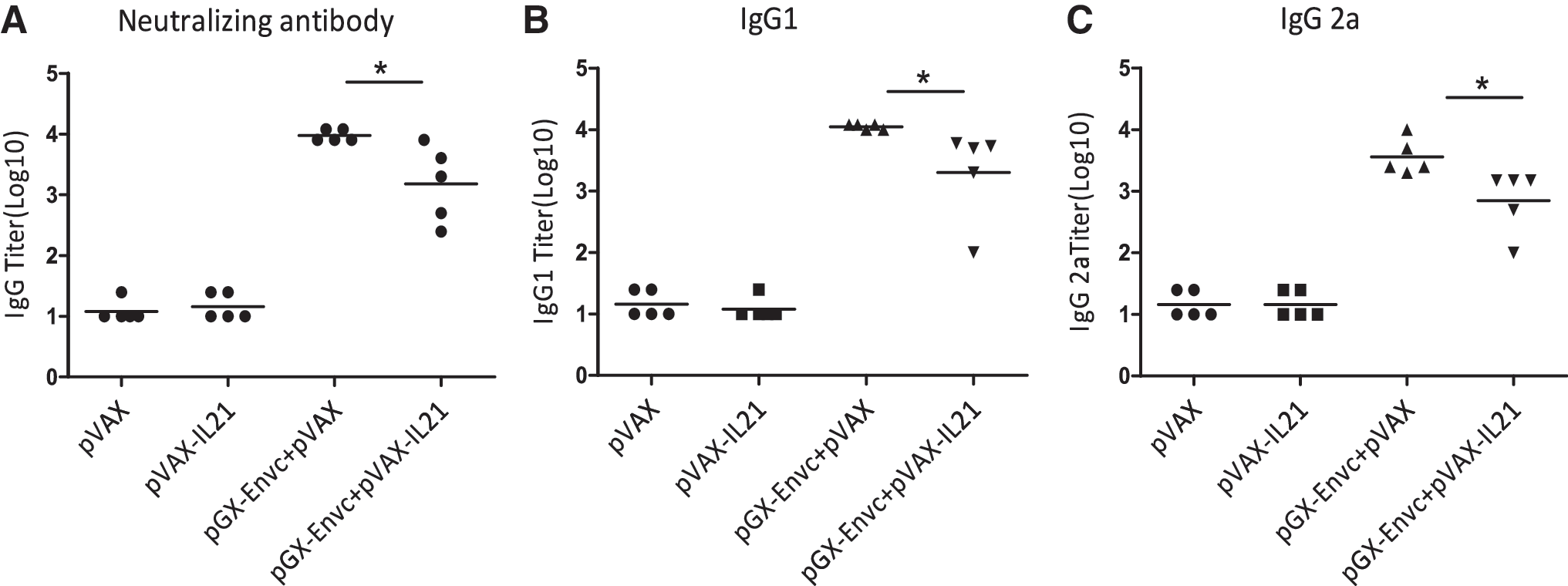
IL-21 as adjuvant inhibits the humoral response. Serum samples from five mice per immunized group were analyzed for antibody titers of total IgG (
IL-21 abates the number of plasma cells
Both IL-4-producing CD4+ T cells and plasma cells could influence the production of antibody (15). We measured the IL-4 level on day 7 after the third immunization. A single-cell suspension of lymphocytes was prepared and re-stimulated in culture with HIV-EnvC peptide as a specific antigen, then analyzed by FACS. The results showed no difference in IL-4 among the four groups (Fig. 5). It is known that TFH cells provide a helper function to B cells (16,17), and that IL-21 plays a critical role in TFH differentiation and function (18 –20). Hence IL-21 could affect B-cell development into plasma cells, and functions (12) through its effects on TFH cells. We examined the effect of IL-21 as an adjuvant on the number of plasma cells and TFH cells on day 7 after the third immunization. From Fig. 7, we can see that the number of plasma cells was decreased by pGX-EnvC plus pVAX-IL-21, whereas there was no difference in the number of TFH cells between these groups (Fig. 6).
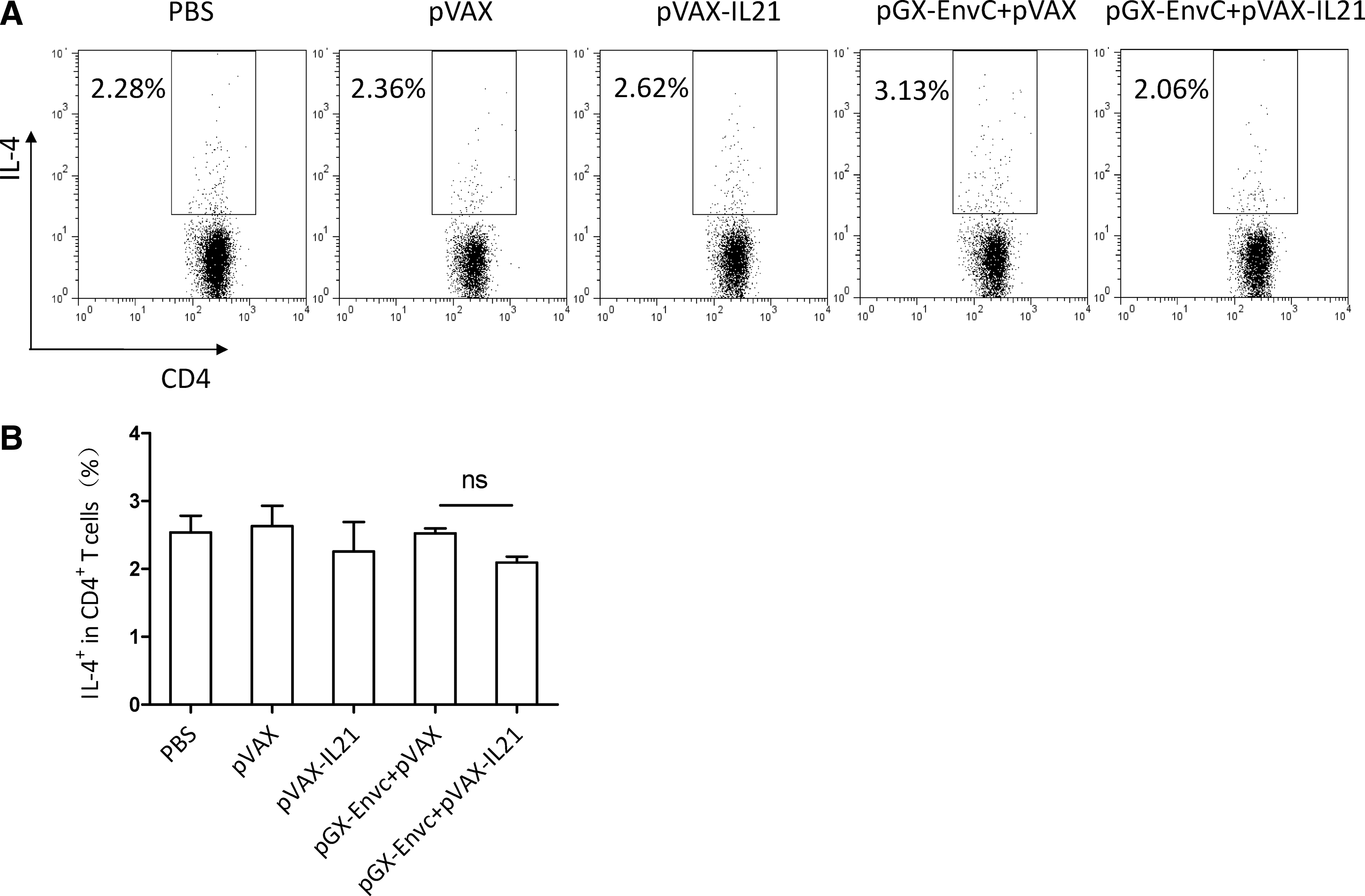
Analysis of IL-4 production in CD4+ T cells by FACS. (
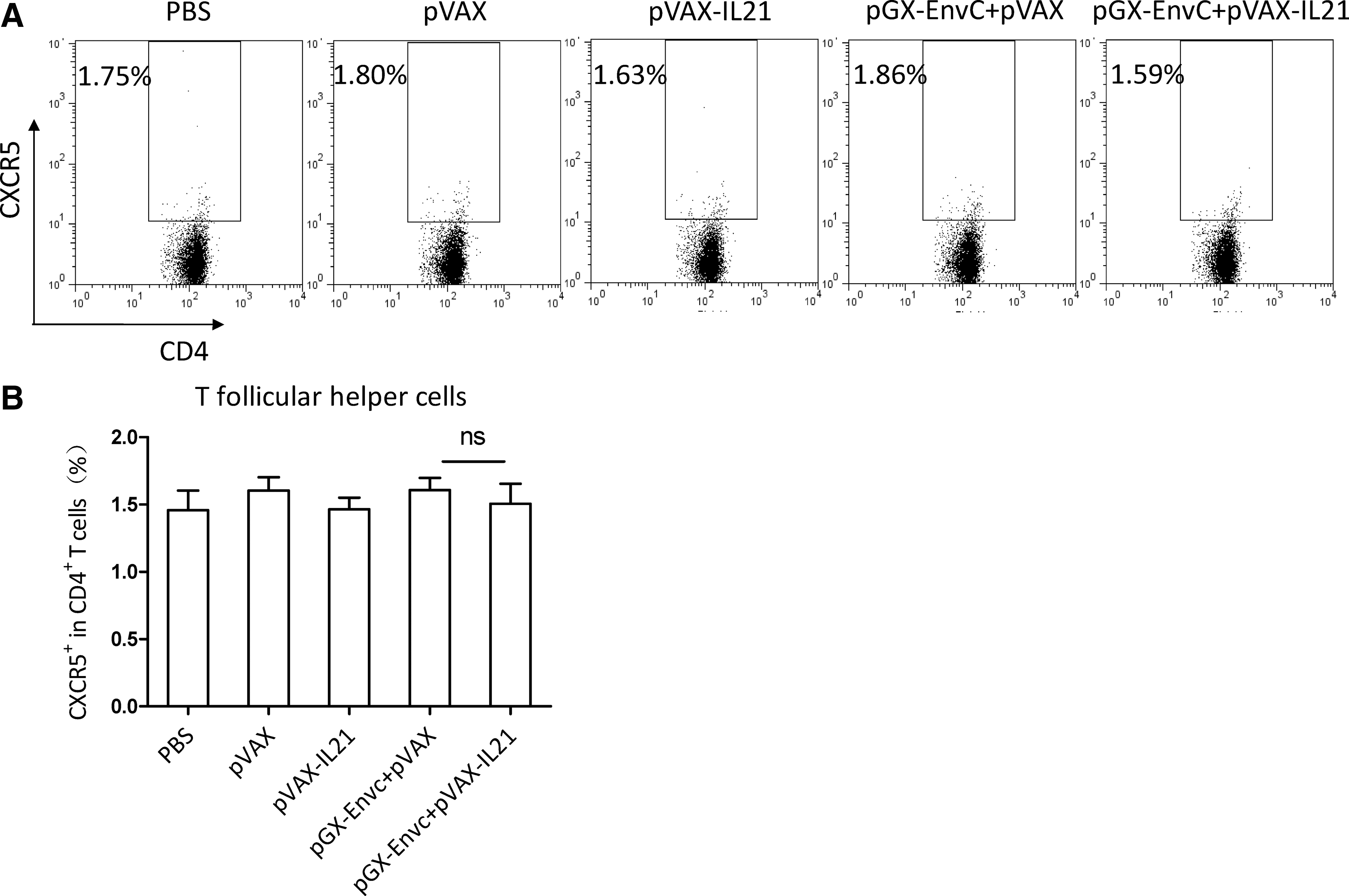
Analysis of follicular helper T cells (TFH) by FACS. (
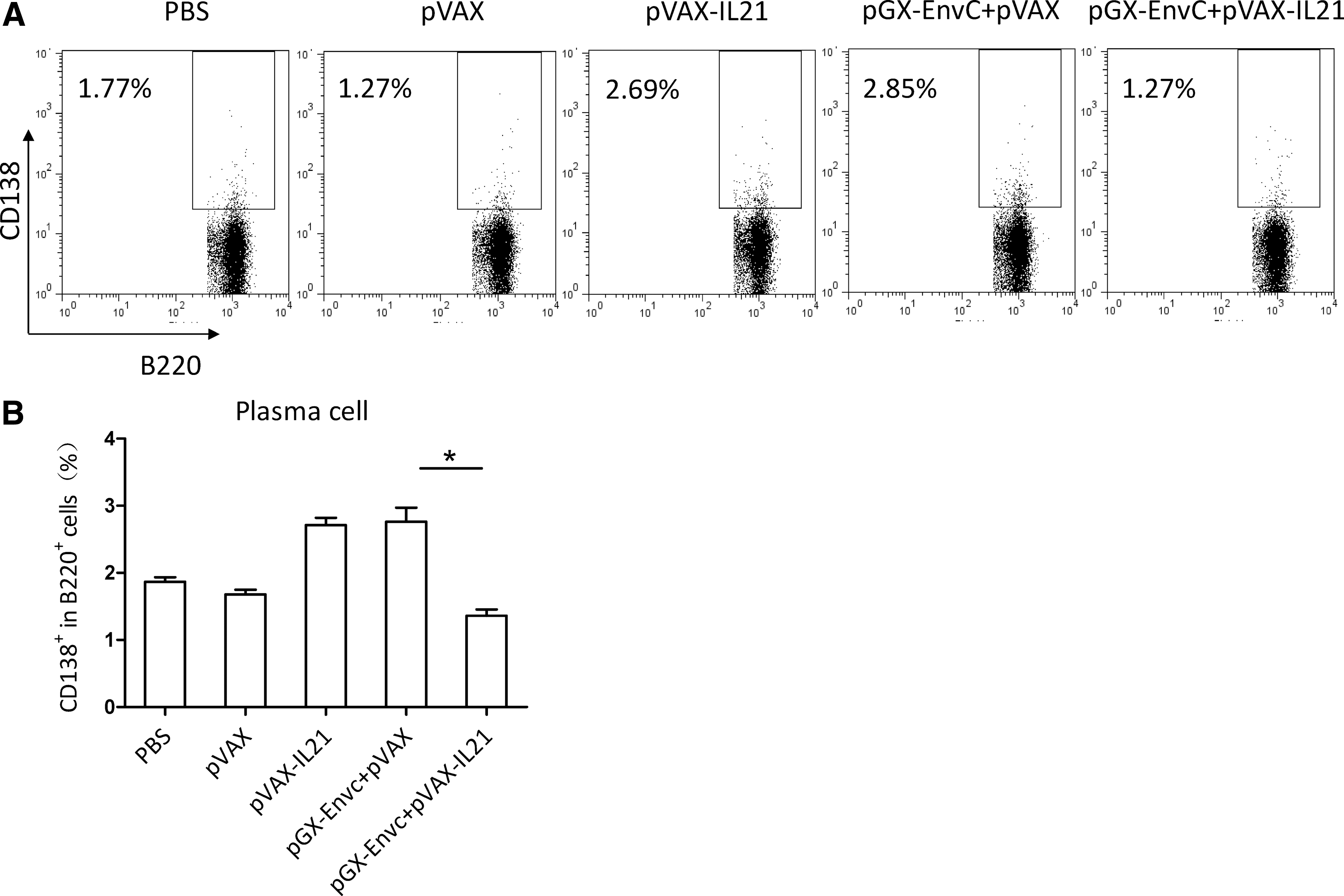
Analysis of plasma cells by FACS. (
IL-21 enhances the mRNA level of Bcl-6 and Pax-5
As the transcription factor Pax-5 and Bcl-6 are known as the main inhibitors of plasma cell differentiation and B-cell activation (21 –24), we examined the effect of IL-21 as adjuvant on these transcription factors in B cells that were purified on day 7 after the third immunization. As depicted in Fig. 8, the levels of mRNA for Bcl-6 and Pax-5 were higher after pGX-EnvC plus pVAX-IL-21 immunization than after pGX-EnvC alone, indicating that IL-21 may decrease the number of plasma cells via enhancing the level of the inhibitory transcription factors.

Expression of inhibitory factor genes in purified B cells by quantitative PCR. Total RNA was isolated from the splenic B cells of mice immunized on day 7 after the final boost. The expression levels of Bcl-6 (
Discussion
Our study demonstrates that IL-21 as molecular adjuvant can induce a much higher antigen-specific cytotoxicity response, and a significantly higher level of IFN-γ in CD8+ T cells during immunization with pGX-EnvC plus pVAX-IL-21. However, IL-21 inhibited the humoral immune responses, which may be due to the high expression levels of inhibitory transcription factor genes of Bcl-6 and Pax-5 induced in B cells by this adjuvant.
This is the first study carried out to determine what immune responses are induced when IL-21 is used as a molecular adjuvant. IL-21 is a novel cytokine produced by activated peripheral blood T cells (25), and acts on a range of lymphohematopoietic cell lineages that are involved in both innate and adaptive immune responses. IL-21 has been shown to be associated with the development and function of TH17, TFH, and B cells (12). Several studies have demonstrated that IL-21 has potent antitumor activity, but it is also associated with the development of autoimmune disease (26,27). In our study, we observed that IL-21, when expressed as a molecular adjuvant from transduced DNA, could increase the cytotoxicity response, but inhibit the humoral response to a DNA vaccine.
Cellular cytotoxic immune responses play a key role in the defense against viral infections. Several subsets of lymphocytes are involved in this process. IFN-γ-producing CD8+ T cells (Tc1) comprise one subset of these cells that is found in the lung following primary challenge with influenza A, and protects against lethal secondary influenza challenge (28,29). In our study we found high levels of expression of IFN-γ in CD8+ T cells, suggesting that IL-21 might increase cytotoxic responses through upregulation of IFN-γ.
The humoral response is another critical aspect in the protection against viruses and other pathogens. Once a B cell encounters its cognate antigen and receives an additional signal from a T-helper cell, it can further differentiate into plasma B cells and memory B cells, and then antigen-specific antibody is produced by plasma cells. Pax-5 and Bcl-6 are the main inhibitors of plasma cell differentiation, and differentiation is inhibited if the two factors are overexpressed (30). In our study, we observed that the IL-21-induced decrease in the number of plasma cells was accompanied by increased expression of Pax-5 and Bcl-6, suggesting mediation by these inhibitory transcription factors.
It is worth mentioning that IL-21 was able to increase the expression of IFN-γ in CD8+ T cells, but decrease antibody level in B cells. IL-21 has been demonstrated to upregulate the expression of genes associated with innate immunity, including the IFN-γ, T-bet, IL-12Rβ2, and IL-18R genes, along with STAT3 activation (31,32). Perhaps IL-21 might induce the expression of IFN-γ in CD8+ T cells via binding to IL-21R through inducing STAT3 and JAK1 and JAK3 tyrosine phosphorylation in our study. To date, the function of IL-21 signaling in B cells has not been extensively characterized. Initial studies demonstrated that IL-21 co-stimulates human B-cell proliferation induced by anti-CD40 Ab, whereas it inhibits proliferation induced by anti-IgM Ab and IL-4 (25). Recently it has been shown IL-21 can induce the apoptosis of primary B cells, even in the presence of pro-survival co-stimulatory factors such as IL-4, LPS, or anti-CD40 Ab (11). Following the initiation of a B-cell immune response, this apoptotic signal of IL-21 might help to eliminate “by-stander” B cells responsible for the nonspecific hypergammaglobulinemia that is initially observed (11,12). Here, our results indicate that IL-21 decreases Ig production in plasma cells. We hypothesize that the effect of IL-21 on plasma cell differentiation uses a novel signal pathway, resulting in its ability to increase Bcl-6 and Pax-5.
In summary, our results show for the first time that an HIV DNA vaccine combined with an IL-21-expressing plasmid can enhance Tc1-mediated cytotoxic responses, and decrease humoral responses due to the inhibition of gene expressions that are important for plasma differentiation. Thus, the use of IL-21 as an adjuvant for DNA vaccination may be limited to situations when the Tc1 response is needed.
Footnotes
Acknowledgments
This work was supported in part by National Key Technology R&D Program of China (2004BA519A39 and 2006BAD06A06), National Nature Science Foundation (30771602), and National Mega Grand Program on Key Infectious Diseases of China (2008ZX10001-012) to B.W. The authors would also like to thank Dr. Jane Q.L. Yu and Zhonghuai He for their technical assistance.
Author Disclosure Statement
No competing financial interests exist.