
Abstract
The use of virosomes as a vaccine platform has proven successful against several viruses. Here we examined the protective efficacy of a virosome-based vaccine consisting of avian influenza virus (AIV) A/Duck/Czech/56/H4N6 in chickens against a homologous AIV challenge. Virosomes adjuvanted with CpG-ODN or recombinant chicken interferon (IFN)-γ significantly reduced virus shedding after virus challenge. Furthermore, immunization with virosomes adjuvanted with CpG-ODN increased hemagglutination inhibition (HI) and virus-specific neutralizing serum antibodies, as well as virus-specific serum IgG and mucosal IgA responses. We also found a significant increase in the expression of type I and II interferon genes in the protected birds following virus challenge. In summary, this study demonstrated the ability of virosomes adjuvanted with CpG-ODN to reduce AIV shedding, and elicit virus-specific protective antibody responses in vaccinated birds.
Introduction
Recent studies have revealed the usefulness of virosomes as a means of vaccination and protection against a wide range of viral infections, including influenza viruses (7,18). Reconstituted influenza virosomes consisting mainly of viral surface glycoproteins, hemagglutinin (HA), and neuraminidase (NA), can induce both antigen-specific antibody- and cell-mediated immune responses (4,11). Although virosome-based vaccines against influenza viruses have been developed experimentally and also commercially for mammalian species (7,26), the usefulness of these vaccines for conferring protection against AIV in chickens has not yet been addressed. In a recent investigation, we have shown that H4N6 avian influenza virosomes adjuvanted with either CpG-ODN or recombinant chicken interferon-γ (rIFN-γ) had immunogenic ability in chickens (21). In the present study, we tested the potential of AIV-H4N6 virosomes alone, as well as adjuvanted with CpG-ODN or rIFN-γ, in an experimental challenge infection with AIV in chickens. We chose AIV A/Duck/Czech/56/H4N6 as a prototype virus, to take advantage of its low pathogenicity, with the eventual aim of employing the same strategy for highly pathogenic AIV in chickens. We hypothesized that immunizing chickens with AIV-H4N6 virosomes alone or in combination with CpG-ODN or rIFN-γ will confer protection against homologous virus challenge, leading to a reduction in virus shedding from mucosal tissues of infected chickens. Furthermore, we sought to examine the elicitation of virus-specific systemic as well as cell-mediated immune responses to subsequent vaccination and challenge.
Methods and Materials
AIV A/Duck/Czech/56/H4N6 was propagated in embryonated chicken eggs (ECEs), and virus purification was performed as described by Reynolds and Maraqa (15), with minor modifications (21). Virosomes from inactivated influenza A/Duck/Czech/56/H4N6 viruses were prepared following the method originally described by Bagai and Sarkar (5), and modified by Markgraf et al. (23). In order to enhance the immunogenic potential of virosomes, rIFN-γ protein, expressed using the baculovirus expression system (22), or CpG-ODN (T
Results
At day 6 post-challenge, a significant (p≤0.05) reduction in virus shedding was observed in the swab samples collected from chickens that had been immunized with virosomes adjuvanted with CpG-ODN or rIFN-γ compared to control chickens. Although virus shedding was higher at day 3 compared to day 6 post-challenge, a significant reduction was also detected in chickens immunized with CpG-ODN or rIFN-γ compared to control chickens (Fig. 1A; p≤0.05). This was associated with significantly higher HI antibodies in chickens vaccinated with virosomes containing CpG compared to those receiving virosomes alone or in combination with rIFN-γ (Fig. 1B). Chickens that received virosomes containing CpG-ODN also developed significantly higher serum IgG or mucosal IgA antibodies at all post-challenge time points (days 7, 14, and 21 post-challenge), compared to the unimmunized challenged chickens or the chickens that received either virosomes alone or virosomes containing rIFN-γ (p≤0.05; Fig. 1C and D). This observation is in agreement with recent studies that suggested similar CpG-mediated effects in enhancing antibody responses against influenza viruses in mice (33), as well as in chickens (31). However, immunization with virosomes alone did not reduce viral shedding at any of the time points following virus challenge (Fig. 1A). In fact, compared to the unimmunized challenged group, there was significantly more virus shedding in the virosome-treated group. This finding raises the possibility that due to the presence of non-neutralizing antibodies, virus entry into host cells was facilitated. This phenomenon, which is called antibody-dependent enhancement, has been suggested to contribute to the pathogenesis of secondary dengue virus infections (14), West Nile virus infections (24), and some formalin-inactivated vaccines (6), including those for measles and respiratory syncytial viruses (25).
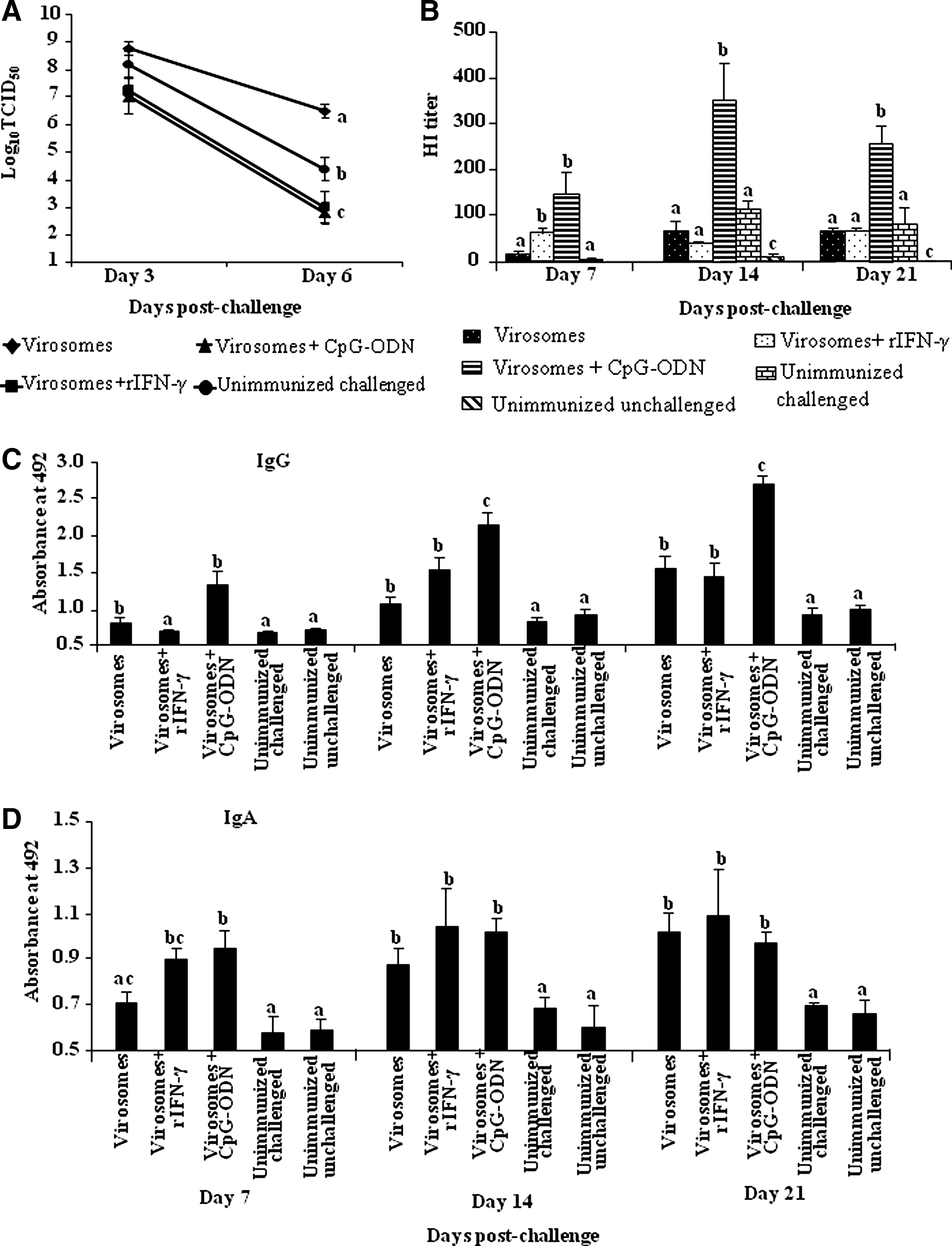
Virus shedding and elicitation of antibody responses following immunization with adjuvanted virosome formulations. (
In addition to HI and antigen-specific antibodies, we examined the presence of neutralizing antibodies in sera of chickens vaccinated with the virosome formulation containing CpG-ODN. A significantly higher virus-neutralizing activity was observed in serum of chickens that received virosomes + CpG-ODN compared to unimmunized control chickens (data not shown).
With respect to expression of interferon genes, we found a significant (p≤0.05) upregulation of the IFN-γ gene on day 14 post-challenge in spleens of chickens immunized with virosomes containing rIFN-γ compared to those that were immunized with virosomes alone or virosomes + CpG-ODN, as well as unimmunized challenged chickens (Fig. 2A). This finding indicates that, at least for the rIFN-γ-containing formulation, the presence of this cytokine might be a correlate of protection. However, IFN-γ did not appear to be correlated with immunity conferred by the CpG-containing formulation. The reason behind this finding is not clear, but it may be related to the time points we selected for sampling, or alternatively, immunity afforded by the CpG-ODN-adjuvanted formulation may be mediated through mechanisms that are IFN-γ-independent.
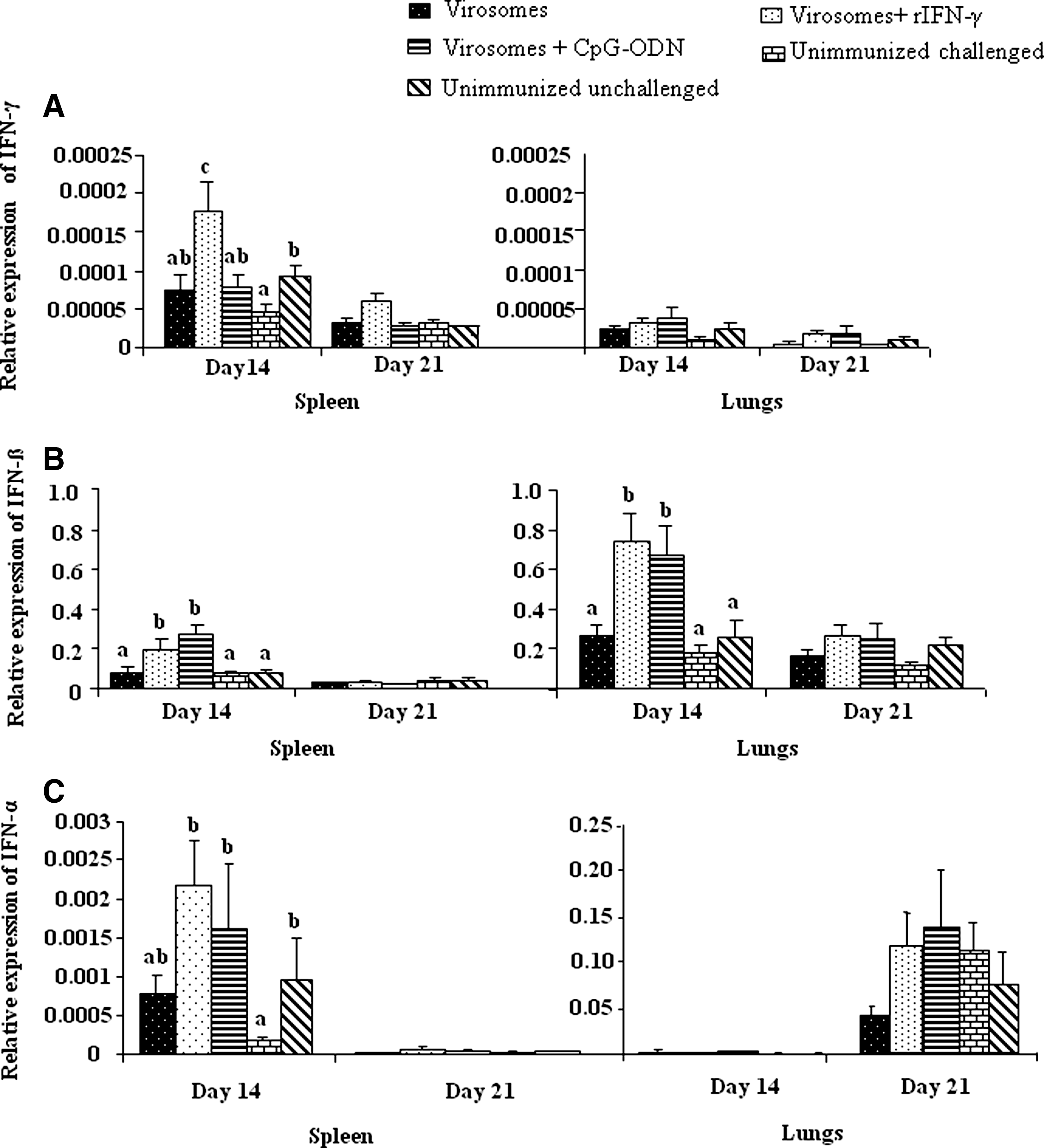
Relative expression of interferon genes in tissues at different post-challenge time points. Interferon gene expression for IFN-γ (
Evaluation of transcriptional changes in type I IFN gene expression in immunized chickens showed significantly (p≤0.05) higher expression of IFN-β in both spleen and lungs of chickens immunized with virosomes containing CpG-ODN or rIFN-γ at day 14 post-challenge, compared to chickens immunized with virosomes alone (Fig. 2B). In addition, significantly higher expression of IFN-α was also observed in the spleens of chickens immunized with virosomes alone, or virosomes containing CpG-ODN or rIFN-γ, at day 14 post-challenge, compared to unimmunized controls (Fig. 2C, p≤0.05). At this point, the role of type-I IFNs in adjuvanted virosome-mediated immunity is not clear. However, it is possible that the increased transcription of these interferon genes might have resulted from either the increased proliferation of IFN-α/β-producing cells such as lymphoid and myeloid cells, or other cell types. Alternatively, this may be due to increased migration of interferon-producing cells to these tissues. In the present study, we also noted a significant reduction in the transcription of IFN-α and IFN-γ in the unimmunized challenged birds compared to those that neither received the vaccine nor the virus challenge. This finding may suggest the influence of AIV non-structural 1 (NS1) gene on IFN expression as an immune-evasive mechanism (12).
Discussion
Altogether, the findings presented here demonstrated that vaccination of chickens with virosomes adjuvanted with CpG-ODN can induce high HI, as well as systemic and mucosal antibodies, and more importantly, can significantly lower virus shedding following virus challenge. It is known that CpG-ODN can lead to the induction of innate and adaptive antiviral responses (29). Importantly, CpG-ODN as a vaccine adjuvant can effectively enhance B-cell activation, and subsequent plasma cell proliferation and differentiation, leading to enhanced antigen-specific antibody production in mammals (11). In chickens, TLR21, a functional homologue of mammalian TLR9, has been demonstrated to play a key role in the recognition and response to CpG-ODN (8). In the present study, it is possible that the physical association of CpG-ODN with influenza virosomal surface proteins may have offered an advantage for delivery of these molecules to the host cell cytosol due to the membrane fusogenic activity of virosomes (32). This may lead to combined cell activation through CpG-ODN, as well as presentation of virosomal antigens by the same cell type.
Footnotes
Acknowledgments
This study was funded by the Saskatchewan Chicken Industry Development Fund, the Ontario Ministry of Agriculture, Food, and Rural Affairs, the Poultry Industry Council (PIC), the Canadian Poultry Research Council (CPRC), and the Natural Sciences and Engineering Research Council of Canada (NSERC). We would also like to thank the staff of the Animal Isolation Facility of the University of Guelph.
Author Disclosure Statement
No competing financial interests exist.