
Abstract
Viral CC chemokine inhibitor (vCCI) of the clone P13 vaccinia virus (VACV) strain PRAHA lacks eight amino acids in the signal peptide sequence. To study the influence of vCCI on virus biology, a virus with the vCCI gene coding for a prolonged signal sequence was prepared. We found that secreted vCCI attenuated the virus in vivo, and that it correlated with decreased levels of RANTES, eotaxin, TARC, and MDC in the blood in comparison with the parental virus. We determined the influence of vCCI on the CTL response against VACV E3(140–148) (VGPSNSPTF) and HPV16 E7(49–57) (RAHYNIVTF) H-2Db-restricted epitopes. The examination of the specific CTL response elicited by immunization with the recombinant VACV-expressing tumor-associated HPV16 E7 antigen by IFN-γ ELISPOT showed that the immunogenicity of the recombinant VACV-producing secretory vCCI was similar to that of the parent virus or deletion mutant in the C23L/B29R locus. Immunization with the secretory vCCI-producing recombinant virus has a lower therapeutic anti-tumor effect against TC-1 tumors. Viral CCI downregulated the E7-specific response induced by gene gun immunization with the DNA vaccines pBSC-SigE7 LAMP and pBSC-vCCI. We also observed that the immune response against vCCI elicited by the DNA vaccine did not affect the multiplication of VACV in vivo.
Introduction
Chemokines play an important role in tumorigenesis. Tumor cells and stromal cells like fibroblasts and leukocytes produce many chemokines which sustain the growth of tumor cells, induce angiogenesis, and facilitate the immune escape of tumor cells. Some CC chemokines contribute to angiogenic progression and development. Monocyte chemotactic protein-1 (MCP-1 or CCL2) is known to potentiate the activity of vascular endothelial growth factor (VEGF) (11) and transforming growth factor-β (TGF-β) (12). MCP-1 expression in the tumor may mediate angiogenesis through prominent recruitment of macrophages, which subsequently produce angiogenic factors such as tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), and IL-8 (13,14). In addition to MCP-1, other CC chemokines have been shown to mediate angiogenesis and modulate extracellular matrix in vivo (15 –20). The chemokines MCP-1 and RANTES (CCL5) produced by tumors induce the formation of the immunosuppressive environment through initiation of migration of tumor-associated macrophages (TAM) into tumor stroma, which leads to the inhibition of tumor-specific T-cell effectors (21). Attracted TAMs are also stimulated by these chemokines to higher levels of production of pro-tumorigenic factors that promote angiogenesis and degradation of the extracellular matrix, such as the matrix metalloproteinases MMP2 and MMP3, and production of inflammatory cytokines like TNF-α, that induce additional production of tumor-supporting factors such as MMP and the chemokines MCP-1 and RANTES by tumor cells. This shows that MCP-1 and RANTES are at the beginning of the malignant process, inducing repeated cycles of attraction of monocytes and secretion of pro-tumorigenic factors (22). Although the described mode of MCP-1 and RANTES action is employed during breast cancer progression, a similar situation may arise during cervical cancer progression. Elevated plasma levels of RANTES were found in higher stages of cervical cancer, and the amount of RANTES was considerably increased in the primary tumor and metastases in all patients. The higher number of macrophages developing the pro-malignant phenotype correlating with carcinogenesis was observed in the tumor stroma of patients with high-grade cervical intraepithelial neoplasia and cancer (23). TAMs also play an important role in the development of TC-1 tumors (24).
Poxviruses have been used many times in experimental cancer therapy. They function either as vehicles for delivering therapeutic genes such as tumor-associated antigens and immunomodulatory molecules, or as oncolytic agents (25). To date, a large number of clinical studies have been conducted with vaccinia virus (VACV) vectors. It has been shown that immunization with the vaccinia virus expressing human papillomavirus 16 and 18 E6 and E7 proteins has an anti-tumor effect, and the capability to elicit antigen-specific T-cell responses in laboratory mice (26,27). The therapeutic effect of these vaccines has also been shown in a number of studies in patients with vulval and vaginal neoplasia (28 –30).
In our work, we wanted to evaluate the contribution of vCCI to the capability of VACV to induce an immune response. To uncover the role of vCCI in the induction of an antitumor effect by immunization with a recombinant vector derived from the vaccinia virus strain Praha, we constructed deletion and insertion mutants encompassing C23L/B29R loci, and expressing tumor-associated antigen HPV16 E7 protein. Beside the evaluation of the specific CTL response by IFN-γ ELISPOT, the anti-tumor effect against TC-1 tumors was tested in therapeutic and preventive arrangements. We also determined the ability of rVACV to modulate chemokine levels in the sera of immunized mice. We showed that vCCI decreased specific cellular immunity elicited by DNA vaccines, and that the vCCI-specific immune response does not affect multiplication of VACV in vivo.
Materials and Methods
Plasmids
The pD357 plasmid for deletion of vCCI genes from the vaccinia virus was obtained from A. Patel (Glasgow) (5). To prepare revertant viruses, the 4889-bp fragment containing the C23L gene was amplified by PCR using PfuI polymerase, primers C23L-1: 5′-AACAAAGATTCTTCCTCCAAACA-3′ and C23L-2: 5′-AAAAGAAGAAGTCGCGAGAGG-3′, and genomic DNA of P13 virus as a template. The amplified nucleotide sequence was inserted into pCR Blunt TOPO plasmid (Invitrogen, Carlsbad, CA) and verified by sequencing. The pD357 plasmid was digested partially with HpaI and NruI, and a fragment containing the C23L gene was substituted for the β-galactosidase gene. The resulting plasmid, denoted as pD357-C23L, was cleaved with EcoRI, and the cassette containing the guanosine phosphoribosyl transferase (gpt) gene from E. coli and the vaccinia virus p7.5 promoter was inserted. The gpt cassette had been excised from the plasmid pGPT07 (31) and provided with EcoRI-compatible ends. The resulting plasmid was denoted as pD357-Rev. The plasmid for generation of P13 virus, which could express the secreted vCCI, was prepared from pD357-C23L plasmid by cutting with SphI and ligation with the annealed oligonucleotides 35Sig-1: 5′-TATGTGCCTGGCGGCAGCTGCCATG-3′, and 35Sig-2: 5′-GCAGCTGCCGCCAGGCACATACATG-3′. The sequence and orientation of the insert was determined by sequencing (Fig. 1C). In the next step, the plasmid was cleaved with EcoRI and the fragment containing the gpt cassette was inserted. The resulting plasmid was denoted as pD357-Rev+Sig. The expression plasmid pBSC-Sig vCCI was prepared by the ligation of the pBSC plasmid (32) digested with XhoI and BglII restriction endonucleases with the PCR fragment amplified using vaccinia LIVP1: 5′-ATT

N-terminal sequence of vCCI of the vaccine strain PRAHA. (
Viruses and cells
Vaccinia virus clone P13 was isolated from the Sevac VARIE smallpox vaccine (strain Praha) (33). Vaccinia virus LIVP (Lister strain) was obtained from W. Chernos, Moscow, Russia. Viruses were multiplied in BSC40 cells, purified by sucrose-gradient centrifugation (34), and titrated in CV-1 cells. The recombinant virus P13-E7GGG (35) contains the gene of modified E7 protein inserted in the TK gene. E7GGG expression is controlled from the H5 early-late promoter. The P13-PreS2-S virus was described previously (36). HPV16 E6E7-expressing TC-1 cells, derived from C57BL/6 mice, were kindly provided by T.C. Wu (Baltimore). The cells were cultivated in modified E-MEM (EPL) medium containing bovine serum growth-active proteins but no complete serum (37). RMA-S cells obtained from M. Smahel from our department were grown in RPMI-1640 supplemented with 10% FCS, 2 mM L-glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, and 10 μM β2-mercaptoethanol.
Construction of VV recombinants
Deletion of the vCCI gene (C23L/B29R loci) was achieved by their replacement with the β-galactosidase gene using homologous recombination with the plasmid pD357 (5). Double deletion mutants were selected by repeated isolation of stable “blue plaque” viruses. The absence of the vCCI coding sequence was confirmed by PCR using primers 5′-TCTCTTCTTCGCTATCTG-3′ and 5′-ACACCGACCAATGATAAGAT-3′. The resulting virus was denoted as P13-ΔvCCI. To prepare the virus carrying the gene for vCCI containing the full-length signal sequence (P13-Sig-vCCI), the P13-ΔvCCI virus was recombined with the pD357-Rev+Sig plasmid. The P13-Sig-vCCI virus was isolated using transient dominant selection (31). First, recombinant viruses carrying the gpt gene were selected and double plaque purified under selection pressure (mycophenolic acid 25 μg/mL, xanthine 250 μg/mL, and hypoxanthine 14 μg/mL). Subsequently, another double plaque purification procedure with selection of “white plaques” enabled reversion to the gpt− phenotype and insertion of the vCCI coding region into both loci. The revertants of P13-ΔvCCI and P13-Sig-vCCI to the parental genotype, which are denoted as RevP13-ΔvCCI and RevP13-Sig-vCCI, respectively, were prepared by recombination with pD357-Rev and selected by the same method. Revertant viruses were confirmed by PCR.
DNA immunization
Mice were immunized at a 2-week interval with three doses of plasmid DNA coated onto 0.5-mg, 1-μm gold particles (Bio-Rad, Hercules, CA) by the procedure recommended by the producer. Vaccines were delivered into the shaven skin of the abdomen at a discharge pressure of 400 psi by a gene gun (Bio-Rad).
Antibodies to viral chemokine inhibitor
Mouse antisera against vCCI were prepared by immunization with DNA vaccine encoding pBSC-vCCI. Mice received three doses administered at 2-week intervals. The sera were collected 2 wk after the last immunization.
Animal experiments
Six-week-old C57BL/6 (H-2b) female mice obtained from Charles River, Cologne, Germany, were used for all experiments. The animals were maintained under standard conditions at the National Institute of Public Health (Prague). The experiments were performed in compliance with Acts no. 246/92 and 77/2004 on animal protection against cruelty, and Decree no. 311/97 of the Ministry of Health of the Czech Republic, on the care and use of experimental animals. Mice were injected intraperitoneally (IP) with 0.5 mL PBS containing sonicated suspension of rVACV.
Tumor induction
Mice received a dose of 3×104 TC-1 cells injected subcutaneously (SC) into the back. Tumor growth was measured weekly. Differences were statistically analyzed using Prism 3.0 software (GraphPad Software Inc., San Diego, CA).
Immunoblotting
CV-1 cells were infected at MOI 3 with vaccinia virus. Two hours later, the medium was replaced with RPMI without serum. After 20 h, the supernatants were separated, cell cultures were washed twice with PBS, and resuspended in PBS containing protease inhibitor cocktail (Sigma-Aldrich, Steinheim, Germany). The cells were lysed with Laemmli buffer containing 2-mercaptoethanol and heated at 95°C for 5 min. The supernatant of infected cells was supplemented with protease cocktail inhibitor, clarified, and concentrated using Amicon Ultracell-10k (Millipore, Billerica, MA). The concentrate was mixed with Laemmli buffer containing 2-mercaptoethanol and heated at 95°C for 5 min. Proteins were separated by SDS-PAGE in a 12% gel and transferred to a nitrocellulose membrane (Hybond-C Extra, Amersham, Piscataway Township, NJ). The membrane, preincubated with 10% skim milk in PBS for 1 h, was incubated with mouse antiserum against vCCI diluted 1:50 in PBS containing 5% milk at 4°C overnight. The membrane was cut into strips for the examination of the individual mouse sera. After washing, the membrane was incubated with rabbit anti-mouse IgG horseradish-peroxidase-conjugated secondary antibody diluted 1:4000 (Sigma-Aldrich) for 2 h. Proteins were visualized with the ECL Plus system (Amersham).
Ligand binding
The cross-linking experiment was performed as described by Alcami et al. (6). The supernatant of infected CV1 cells was prepared by the same procedure as for immunoblotting. The concentrate, equivalent of 20,000 cells, was used for each cross-linking reaction. We utilized radioiodinated human recombinant 0.4 nM macrophage inflammatory protein-1α (MIP-1α) 125I (2000 Ci/mmol) (GE Healthcare, Pittsburgh, PA), 40 mM 1-ethyl-3(3-dimethyl-aminopropyl)-carbodiimide (EDC) (Sigma-Aldrich), and 5 μL of concentrated supernatant in 25 μL of binding medium (RPMI 1640, 20 mM HEPES [pH 7.4], 0.1% BSA). The samples were analyzed by SDS-PAGE on a 12% gel. The gel was exposed and scanned on a Phosphorimager (Fuji, Cypress, CA).
Detection of chemokine levels by ELISA
Levels of 12 common mouse chemokines (RANTES, MCP-1, MIP-1α, MIP-1β, SDF-1, IP10, MIG, EOTAXIN, TARC, MDC, KC, and 6Ckine) in mouse sera were determined using a Multi-Analyte ELISArray Kit (SABiosciences, Frederick, MD), according to the manufacturer's instructions. To be able to perform the complete assay the sera aliquots of four animals were pooled in each group.
Detection of VACV multiplication in vivo using quantitative PCR (Q-PCR)
Mice were anesthetized with halothane (Narcotan; Léčiva, Prague, Czech Republic) and sacrificed. The ovaries were dissected, washed in PBS, and homogenized. DNA was extracted using a DNeasy Tissue Kit (Qiagen, Hilden, Germany). Real-time quantitative PCR was performed as described previously (38).
Assay of immune responses
E7- and VACV-specific T-cell response
The spleens were removed from mice 12 d after the last immunization and the response of interferon-γ (IFN-γ)-producing cells was determined by ELISPOT IFN-γ assay upon in vitro restimulation with 0.025 μg/mL HPV16 E7(49
–57) (RAHYNIVTF), or 1 μg VACV E3(140–148) (VGPSNSPTF) H-2Db-restricted peptides for 20 h as described previously (38). The working concentration was optimized for each peptide. All peptides were purchased from Clonestar (Brno, Czech Republic). The CTL epitope VACV E3(140–148) was predicted using the RANKPEP, SYFPEITHI algorithm. We have shown repeatedly using the ELISPOT assay and intracellular cytokine staining test (ICS), that the splenocytes of C57BL/6 mice, which had been immunized with various strains of VACV including MVA, responded to stimulation with E3 peptide. To confirm here the applicability of the VACV E3(140–148) (VGPSNSPTF) peptide for the detection of VACV-specific CTLs, we demonstrate that VACV E3(140–148)-specific CTLs are present in animals immunized with recombinant vaccinia virus P13 expressing glucuronidase of E. coli (39), and are absent in non-immunized mice. We show also that such splenocytes are not activated by the HPV16 E7(49
–57) peptide, which is irrelevant for vaccinia virus. The response of splenocytes was determined by ELISPOT-IFN-γ (Supplementary Fig. S1A; see online supplementary material at
Flow cytometry detection of surface molecules on splenocytes
The spleens were mechanically disrupted and a single-cell suspension was acquired after red blood cell lysis using ACK buffer (0.15 M NH4Cl, 10 mM KHCO3, and 0.1 mM EDTA). Cells were stained with the fluorescently-conjugated mAbs CD3-FITC, CD4-FITC, CD8-FITC, CD11b-PE, CD11b-FITC, CD11c-PE, F4/80-PE, and Gr-1(Ly6G-FITC). F4/80-PE mAb was purchased from ABD-Serotec (Dusseldorf, Germany), and the other mAbs were supplied by BD Biosciences Pharmingen. The expression of surface molecules was determined by flow cytometry using a Coulter-Epics XL flow cytometer (Beckman Instruments, Williamsville, NY) and FlowJo software (Tree Star, Inc., Ashland, OR).
Results
The putative viral inhibitor of CC chemokines encoded by the vaccine strain Praha contains a shortened signal peptide sequence
Our goal was to determine whether the viral inhibitor of CC chemokines influences the immunogenicity and anti-tumor activity of recombinant vectors derived from an attenuated vaccinia virus strain. First, we sequenced the region encoding the N-terminal part of vCCI of the P13 clone of the strain PRAHA. The comparison of the amino acid sequence translated from a portion of a 4889-bp fragment amplified using C23L-1 and C23L-2 primers with the amino acid sequence of vCCI from other poxviruses revealed that the strain PRAHA encodes vCCI lacking 8 amino acids inside the signal peptide sequence (Fig. 1A and B).
Prolongation of the signal sequence of vCCI results in restoration of functional vCCI production
The production of vCCI by CV1 cells infected with recombinant viruses derived from P13 virus was examined by immunoblot (Fig. 2A). For the detection, a specific antiserum prepared by immunization of mice with the vCCI gene of LIVP in the form of DNA vaccine was used (see materials and methods section). We did not detect any vCCI in the concentrated culture medium of cells infected with the parent P13 virus carrying the E7GGG gene or in mock-infected cells. In the lysate prepared from cells infected with P13-E7GGG, we found only a faint nonspecific band, which was also present in the lysate of mock-infected cells and cells infected with P13-ΔvCCI-E7GG. To prepare a virus P13 that produces secreted vCCI similarly to the Lister virus, the signal sequence of vCCI-P13 was restored by the insertion of an oligonucleotide coding for eight additional amino acids (Fig. 1C). Immunoblot analysis showed that CV1 cells infected with the resulting virus P13-Sig-vCCI E7GGG secreted the vCCI of the same size as did the LIVP virus (Fig. 2A).

Viral CCI production in vitro by (

The band of intracellular vCCI detected in samples of cells infected with LIVP and P13-Sig-vCCI-E7GGG was more intense than the non-specific band. In the lysate of LIVP-infected cells, two more bands of a smaller size were detected, probably representing vCCI fragments. Moreover, the supernatant of the cells infected with the P13-Sig-vCCI virus showed similar MIP-1α chemokine binding activity as the supernatant from the cells infected with the LIVP virus (Fig. 2B). The supernatant of mock-infected cells or cells infected with the parental P13 virus showed no chemokine binding activity. A faint band of lower size appeared if cross-linking agent was used in the absence of any supernatant.
Secretory vCCI modulates chemokine levels in the blood of VACV-infected mice
To determine the activity of rescued vCCI production in vivo, we examined the amount of common chemokines in the blood of infected mice by ELISA. The results of two experiments are shown in Table 1. The administration of the P13-E7GGG virus increased the level of RANTES, MCP-1, MIG, eotaxin, TARC, MDC, and KC, on day 3 in comparison with mock-infected mice. The blood of all groups of mice contained non-detectable amounts of MIP-1α and GCkine. In comparison with undeleted P13-E7GGG virus, the infection with P13-Sig-vCCI-E7GGG reduced the levels of RANTES, eotaxin, TARC, and MDC in the blood. Production of vCCI by vaccinia virus had no effect on the amount of MCP-1, MIG, or KC on day 3. The levels of eotaxin and MDC decreased gradually, and the differences found on day 4 were smaller than those on day 3. The values could not be statistically analyzed, because the chemokine levels were measured in the pooled sera.
Animals were injected with 3.106 PFU in 0.5 mL PBS IP.
In Experiment 1 blood was collected on day 3. The pool of sera (n=2) was examined for the presence of mouse chemokines.
In Experiment 2 blood was collected on day 4. The pool of sera (n=4) was examined for the presence of CCL11 and CCL22.
The absorbance values were normalized to the baseline value (0%), and to the absorbance that gave the specific chemokine-positive control supplied in the kit (100%). The data were calculated from absorbance values OD 450–OD 630.
Production of secretory vCCI attenuated P13 VACV in vivo
It has been shown previously that the insertion of the C23L/B29R encoding region from the Lister strain into the genome of a highly virulent virus of the WR strain resulted in a decrease of its virulence in mice. We investigated whether secretory vCCI could further reduce the virulence of the highly attenuated vaccine strain Praha. Groups of mice (n=16) were injected IP with 1×106 PFU of each rVACV on day 0. The ovaries from four animals of each group were collected on days 3, 4, 5, and 7. The viral DNA was determined in the DNA isolated from the ovaries by PCR. DNA of the viruses P13-E7GGG and P13-ΔvCCI-E7GGG was detected in five animals, and RevP13-ΔvCCI-E7GGG in six animals, during the observed period. Virus P13-SigvCCI-E7GGG was only found three times on days 3 and 4. Though the differences were not significant due to considerable variability, the results of the DNA assay together with the lower levels of anti-VACV antibodies (Fig. 4B) found in the infected mice, suggested that rescuing secretory vCCI production in P13 virus led to attenuation of viral replication in vivo (Fig. 3A).
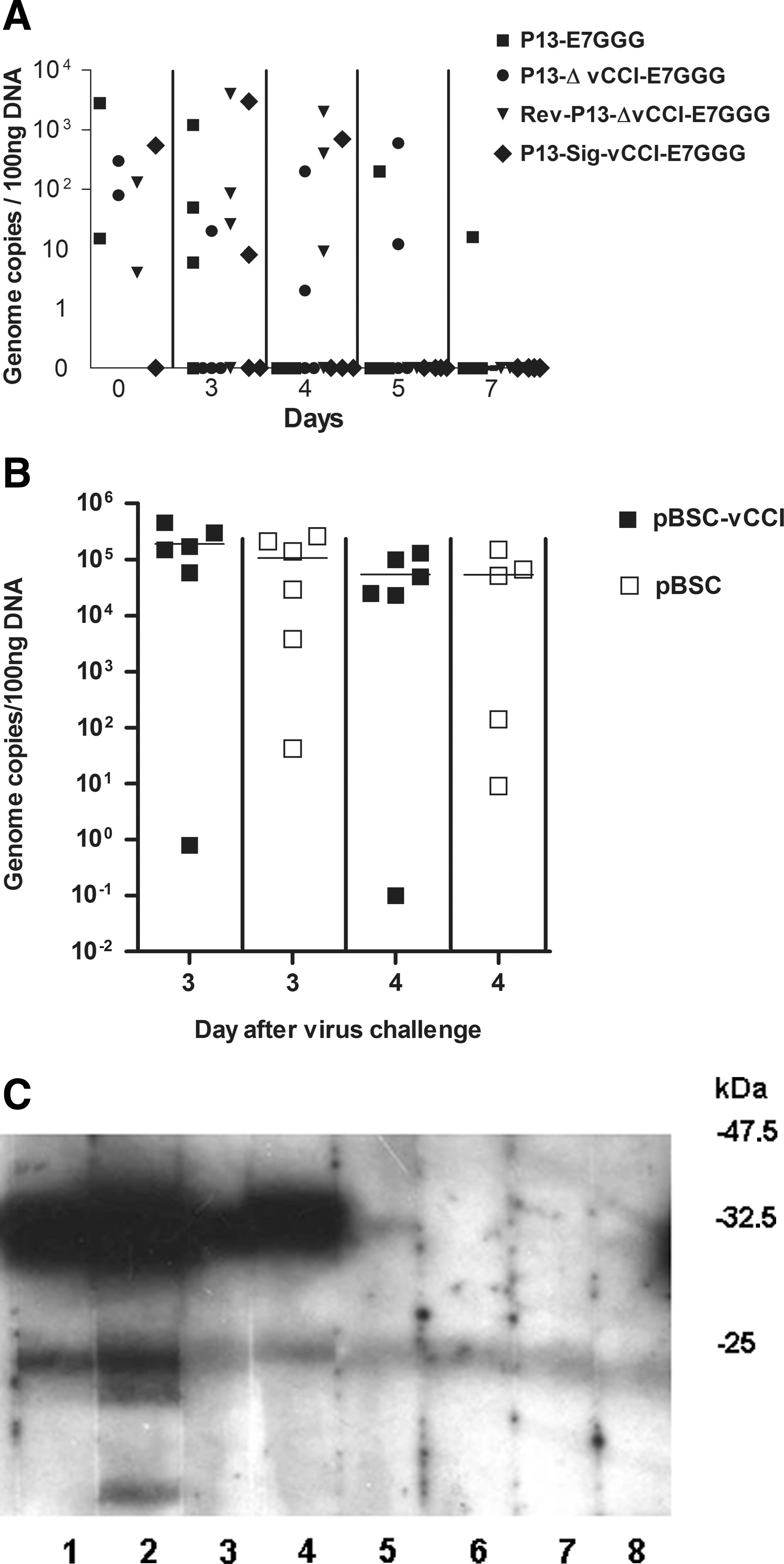
The effect of vCCI on in vivo multiplication of VACV in naïve or immunized mice. (
The response induced in mice by immunization with rVACVs or DNA vaccine co-expressing vCCI. (
Induction of an antigen-specific immune response toward vCCI protein elicited by the DNA vaccine had no effect on VACV replication in vivo
Next we investigated whether the immunity to vCCI could modulate viral replication in vivo. We speculated that neutralization of the secretory vCCI by antibodies might result in the increase of virulence of VACV. Alternatively, the VACV-infected cells could be lysed by the vCCI-specific cytotoxic T cells, which would lead to decreased virus replication in vivo. Mice immunized with three doses of pBSC-vCCI or pBSC vaccine were challenged with 106 PFU P13-Sig vCCI-E7GGG on day 39. The viral DNA was measured by PCR in the DNA isolated from the ovaries on days 3 and 4 after the challenge. No statistically significant difference was found between the groups (Fig. 3B). The efficiency of DNA vaccine was determined by the detection of vCCI-specific antibodies in individual mouse sera by immunoblot, as shown in Fig. 3C. We concluded that the presence of antibodies against vCCI did not affect multiplication of a VACV that produced secretory vCCI.
Secretory vCCI does not enhance the immune response to P13 VACV recombinants and DNA vaccine
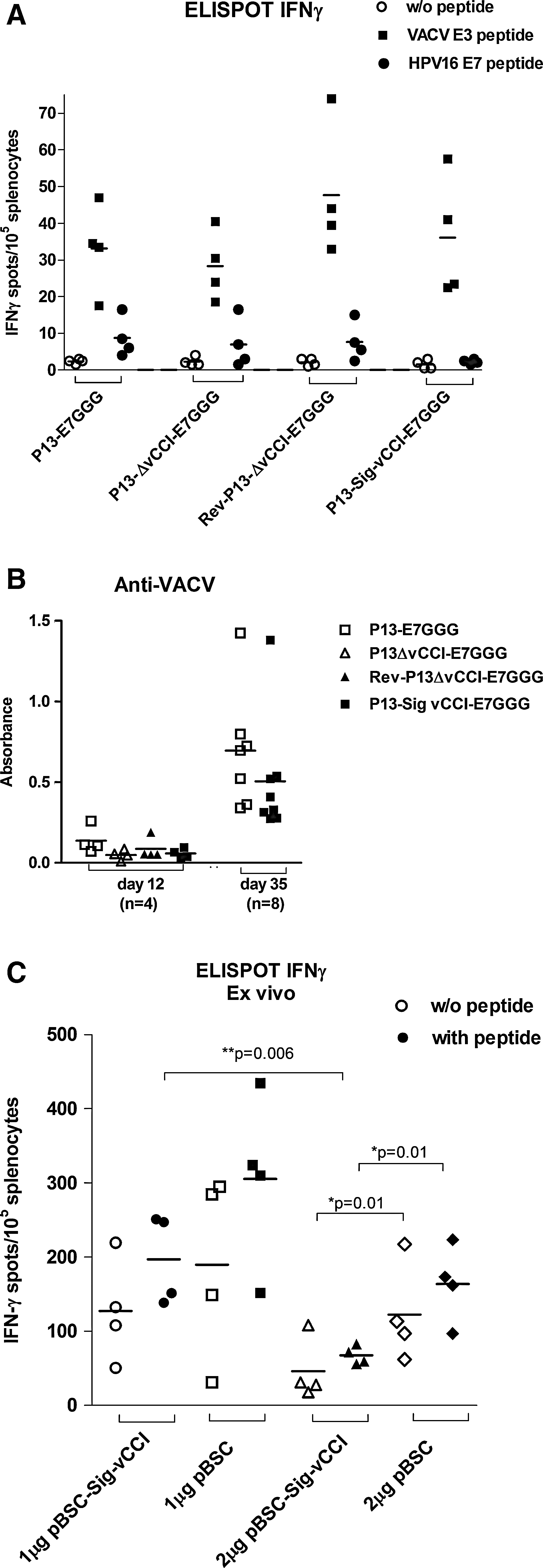
In the next step, we examined the effect of vCCI production on the immunogenicity of recombinant viruses expressing the modified HPV16 E7 protein. We determined the CTL response against the dominant CTL epitope HPV16 E7(49 –57) (RAHYNIVTF), and against the VACV E3(140–148) (VGPSNSPTF) epitope. We found cellular responses elicited by one dose of 106 PFU P13-E7GGG, P13-ΔvCCI-E7GGG, Rev-P13-ΔvCCI-E7GGG, or P13-Sig-vCCI-E7GGG, as detected by IFN-γ ELISPOT (Fig. 4A), to be similar in magnitude, with the differences not being statistically significant. It seems that the response of CD8+ T cells elicited by the recombinant virus is not affected by any form of the vCCI gene. To determine the effect of vCCI expressed by P13-E7GGG on the immune cells, we quantified the major subpopulations of splenocytes by flow cytometry, using surface markers CD4, CD8, CD11b, CD11c, CD25, F4/80, and Gr1. The animals were injected IP with a single dose of 3.106 PFU, and the splenocytes were analyzed on days 3 and 15. We observed that none of the markers had significant changes in comparison with the splenocytes of naïve mice (data not shown). The antibody response against the vaccinia virus was examined at two time points. In the first experiment, blood was collected from animals examined for a CTL response on day 12. In the second experiment, mice were injected with 3.106 PFU and blood was collected on day 35 after virus administration (Fig. 4B). In both experiments P13-E7GGG elicited a stronger response than P13-Sig-vCCI-E7GGG in both groups, though the difference was not statistically significant.
To determine the effects of secretory vCCI on the E7-specific CTL response in the absence of virus infection, we immunized mice against E7 protein with the DNA vaccine in combination with the plasmid carrying the vCCI gene from LIVP virus. The pBSC-SigE7LAMP DNA vaccine and either 1 or 2 μg of pBSC-vCCI or pBSC were shot with a gene gun subsequently to the same site. The animals received two doses 2 wk apart. The CTL response was examined 12 d after the last immunization by IFN-γ ELISPOT (Fig. 4C). We observed that the HPV16-E7-specific response was lower in animals that received the pBSC-vCCI plasmid compared to the empty plasmid, which implies that the viral CCI has no enhancing activity on the antigen-specific CTL response. The difference was statistically significant (p=0.01) when 2 μg of the plasmids were used. Moreover, the immunization with the mixture with 2 μg of the pBSC-vCCI plasmid in comparison with the empty vector resulted in a decrease in the number of IFN-γ spots (p=0.01), when the splenocytes were cultured in the absence of E7 peptide. The effect was dose-dependent, as the addition of 2 μg of the pBSC-vCCI plasmid reduced the response more than 1 μg (p=0.0057).
Anti-tumor effect of therapeutic immunization against TC-1 tumors is decreased by the production of secretory vCCI
To learn whether the binding of chemokines to vCCI produced in mice with growing tumors can improve the anti-tumor effect of therapeutic immunization with VACV expressing HPV16 E7, we immunized mice with P13-E7GGG or P13-Sig-vCCI-E7GGG and its deletion and reversion mutants the day following the administration of the TC-1 cells. As the control virus we used P13-PresS2S, which expresses the coding sequence of the middle protein of the surface antigen of the hepatitis B virus. We observed (Fig. 5) that the immunization with P13-E7GGG or its deletion mutant had a significant anti-tumor effect in comparison with the P13-PresS2S, as determined by the t-test on day 27. The reversion mutants also had an anti-tumor effect, though it was not significant because of the smaller size of the animal groups. P13-Sig-vCCI-E7GGG had the lowest anti-tumor effect, and the difference was statistically significant in comparison with P13-E7GGG. The outcome was different when mice were first immunized with one dose of rVACV and TC-1 cells were administered 2 wk later. The immunization with both P13-E7GGG and P13-Sig-vCCI-E7GGG resulted in similar inhibition of the growth of tumors (data not shown).
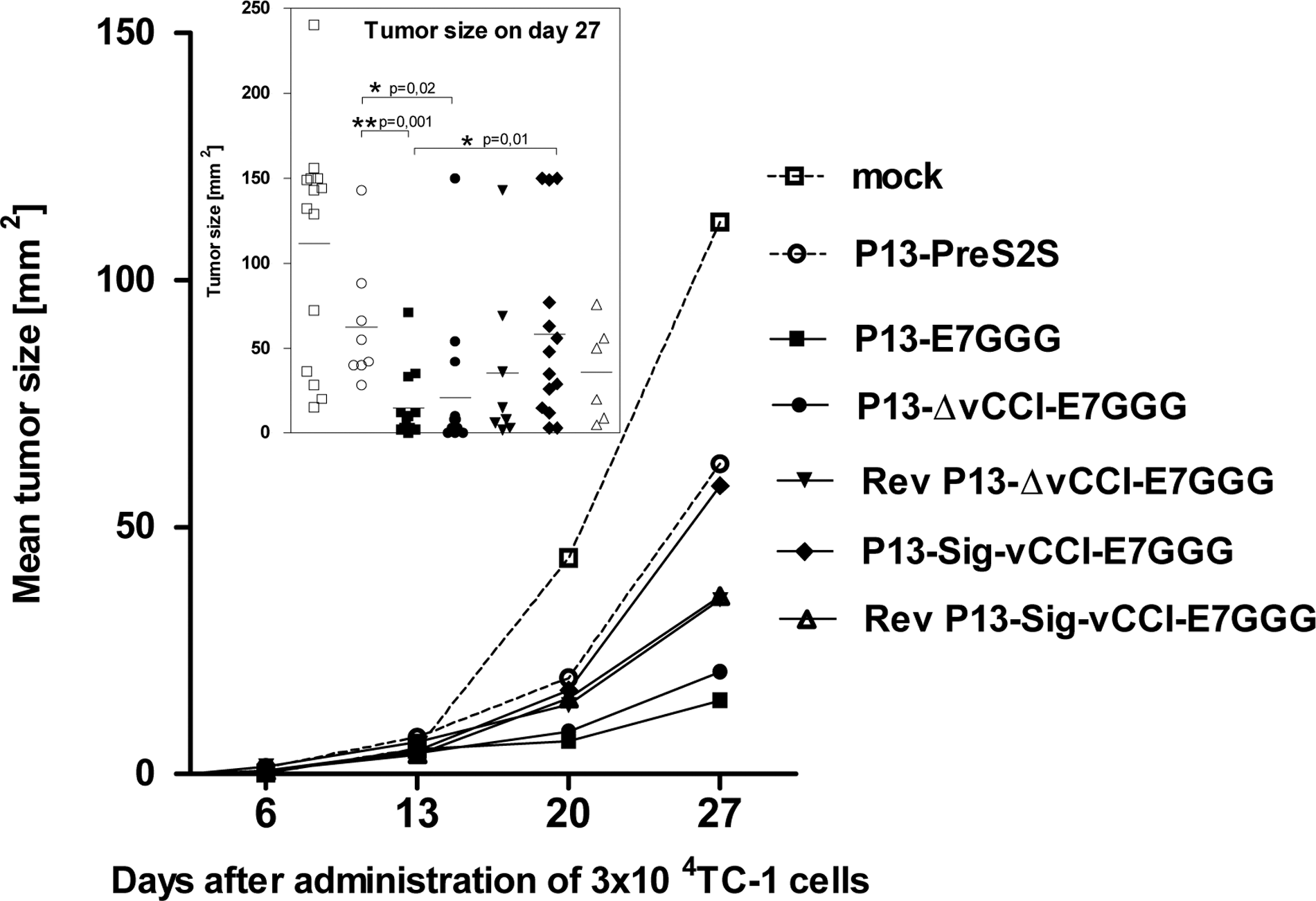
Therapy of TC-1 tumors by immunization against HPV16 E7 with rVACV. Mice were injected SC with 3×104 TC-1 cells. The next day after the administration of tumor cells, the animals were immunized once IP with 3×106 PFU P13-E7GGG (n=14), P13-ΔvCCI-E7GGG (n=14), RevP13-ΔvCCI-E7GGG (n=8), P13-Sig-vCCI-E7GGG (n=14), or RevP13-Sig-vCCI-E7GGG (n=6). The graph shows summarized results of two independent experiments. Tumor growth is shown as the mean tumor area for each group. The differences in the tumor size on day 27 were statistically analyzed by the t-test.
Discussion
In our study, we examined the influence of the immunomodulatory protein vCCI (35K) on the immunogenicity of recombinants derived from clone P13 of the vaccine strain Praha, and their anti-tumor effect against TC-1 tumors. The vCCI protein belongs to the group of secreted viral immunomodulatory proteins. Thanks to its capability to efficiently inhibit CC chemokines that direct migration of macrophages, monocytes, DCs, and lymphocytes, it can influence different stages of the antiviral immune response. The importance of CC chemokines in this process was demonstrated for infection with the MVA virus, which induces an increased level of MCP-1 in contrast to the strains Elstree, Wyeth, and WR. The failure of MVA to prevent chemokine expression by blocking NF-κB activation and sequestering MCP-1 by vCCI presumably contributes substantially to its extraordinary vaccination potential (42). We speculated that the vCCI gene deletion from the vaccine virus Praha could lead to the creation of a virus with enhanced immunogenicity that would induce stronger cellular immunity.
VACV P13 was prepared in our laboratory as a plaque-purified clone of the vaccine strain Praha (33). When compared with the VACV strains Wyeth, and LIVP (Lister), the immunogenicity and virulence of the strain Praha is between that of LIVP and Wyeth virus. The P13 clone is the most virulent in comparison to the other Praha clones. The virus LIVP was the most immunogenic and virulent of the three strains tested. To determine the influence of vCCI on the biological properties of the vaccine virus P13, we constructed deletion mutants with insertion inactivation of the C23L/B29R gene, and revertant viruses with genes inserted at both loci in the ITRs. In our experiments, we used the P13 virus expressing the weakly immunogenic HPV16 antigen E7GGG, because it is more suitable for the evaluation of minor differences in the immunogenicity of the vector than the other VACV P13-SigE7LAMP, producing a more immunogenic form of E7 antigen. This recombinant induces the complete rejection of TC-1 tumors (35), and would not allow the measurement of the influence of the small difference in virulence associated with the immunogenicity of Sig-vCCI. Immunization with P13-E7GGG does not inhibit TC-1 tumor formation, but it is able to slow tumor growth.
The comparison of the P13 vCCI ORF nucleotide sequence with the orthopoxvirus homologues revealed the highest 5′ end homology of P13 with the vaccinia Copenhagen, WR, Ankara, MVA, and Tian Tian strains, as well as with most variola and monkeypox strains. The gene of these viruses encodes CCI protein lacking the N-terminal signal peptide, unlike vCCI of VACV Lister, the cowpox viruses Brighton and GRI-90, and mousepox virus, which have an N-terminal signal peptide with high cleavage probability between proline 18 and alanine 19. A serine-rich sequence at the N-terminus that is a part of a random coil of the vCCI molecule contains four residues in the Praha strain. The chemokine binding activity of vCCI is not changed, whether it contains four or seven serines in orthopoxviruses, or the TKGI sequence, which was found in leporipoxviruses (8). The signal peptide is essential for CCI secretion, and therefore rVACV P13 with the complete signal sequence of vCCI was created. It was confirmed by Western blot that the virus prepared secretes CCI protein into the medium, and the cross-linking experiment showed that vSigCCI is able to bind the 125I-radiolabeled chemokine MIP-1α. The biological activity of vSigCCI produced by P13 virus was also observed in vivo.
The role of immunomodulatory proteins such as CC chemokine inhibitors in the pathogenesis of poxvirus infection can be tested by comparison of the reactions of the host after the application of the wild-type virus, and the deletion mutants lacking the studied immunomodulatory protein. The application route used for the evaluation of VACV CC chemokine inhibitors in the mouse model was usually intradermal or intranasal administration (43). We have shown that the secreted form of vCCI attenuated the multiplication of IP-injected virus similarly to that previously described for IN-applied virus (10). However, the IN and ID routes appropriately mimic the typical natural routes of poxvirus infection, and they result in high antibody responses but very low CTL activity. On the other hand, the systemic IP and IV application routes lead to induction of a strong CTL response (44). The attenuated P13-E7GGG was able to elicit a significant CTL response to HPV E7 only after IP inoculation of a high dose 106 PFU, and for this reason we chose this route of application. Moreover, IP injection was more convenient than the IN route, as we wanted to determine the influence of the secreted form of vCCI on the anti-tumor effect of immunization. It is known that the CC chemokines play an important role during the development of various tumors (45). Even though the influence of chemokines is primarily exerted locally, elevated serum levels are often observed, which correlate with the severity of disease. Blockade of the CC chemokines after the systemic administration of their antagonists may contribute to the inhibition of the growth of tumors, as was tested in the past in several preclinical studies (46 –49). The positive effect of CC chemokine antagonists on the immunotherapy of tumors can be also detected via the analysis of the systemic and intratumoral immune responses (50). We also wanted to evoke the systemic production of vCCI, which could eventually act in the tumor microenvironment in the course of tumorigenesis and induce an anti-tumor response.
When measuring chemokine levels in the serum of mice on day 3 after IP infection by the parental virus P13-E7GGG, we found increased levels of the chemokines MCP-1, MIG, RANTES, and MDC. The P13-E7GGG virus is similar in this feature to the MVA virus, which induces the expression of the chemokines MCP-1, MIP-1α, RANTES, and MIG, both after in vitro infection of macrophages (51), and after IN application to mice (42). On the contrary, the strains NYVAC and WR after in vitro infection of macrophages, and the strains Elstree, Wyeth, and WR after IN application to mice, do not induce the production of CC chemokines (42). This presumably relates to the influence of other immunomodulatory proteins present in the strains mentioned above.
While testing the influence of vCCI produced by the IP-administered virus P13-SigCCI-E7GGG on the levels of chemokines, we observed, when compared to controls, a decrease of the chemokines RANTES and eotaxin (i.e., the second and the third chemokines for which vCCI has the highest binding specificity). We also detected a drop in the chemokines TARC and MDC. The chemokine MIP-1α that binds best to the vCCI, and by the help of which we proved the binding capability of vCCI, was not detected in the mouse serum by the test we used. The level of MCP-1, the fourth best vCCI-bound chemokine, was not influenced by the production of vCCI. This suggests that vCCI is not able to neutralize MCP-1 produced by TC-1 as detected by expression analysis (52), and could partly explain why P13-Sig-vCCI-E7GGG does not exert improved anti-tumor effects. Despite the differences in the levels of chemokines induced by rVACV, the analysis of the immune cells isolated from the mouse spleens after IP application of rVACV with various forms of vCCI found that vCCI produced by P13 did not affect the amount of DCs, myeloid suppressor cells, T-regulatory lymphocytes, CD8+ lymphocytes, and macrophages.
The examination of revertants carrying vSigCCI showed that the production of the secretory vCCI protein does not influence virus replication in vitro, though the measurement of viral DNA in the mouse ovaries injected IP with VACV revealed that the revertant expressing vSigCCI is more attenuated, because after systemic administration, it replicates to lower titers than the parental virus. The decreased replication was also confirmed by the detection of lower levels of specific anti-VACV antibodies on day 32 after infection in response to P13-SigCCI-E7GGG compared to the antibodies elicited by the parental virus. We and others have observed previously that attenuated viruses induced lower titers of antibodies (53). Similar results were obtained by Reading et al. (10) after IN infection of mice with the WR virus that produced the vCCI of the Lister virus. Decreased titers of WR-vCCI were accompanied by less severe infiltration of the lungs by macrophages and lymphocytes, which correlated with reduced levels of the chemokines MIP-1α, MCP-1, and eotaxin detected in the BAL fluid of IN-infected mice. We also observed a decrease in the level of RANTES.
RANTES is produced in the early stages of infection, mainly by the antigen-presenting cells (DCs, macrophages, NKT cells, and γðT cells), which have receptors for inflammatory cytokines, so they significantly respond to inflammatory conditions (54). RANTES attracts to locations of inflammation and infection cells that express the receptors CCR1, CCR3, and CCR5. They are primarily the cells of innate immunity, such as monocytes, basophils, eosinophils, immature DCs, NK cells, and T cells. It has been demonstrated (55) that activation of CCR5 induced by VACV infection leads to the induction of the permissive phenotype for VACV replication in primary T cells. Since permissive infection of T cells and other CCR5-positive cells might represent a mechanism for VACV dissemination throughout the lymphatic system, sequestration of CCL5 by vCCI, which could result in a decrease in CCR5 activation, could be manifest as a decrease in VACV virulence. It has been recently shown that CCR5−/− mice are less susceptible to systemic VACV infection than WT mice (56).
The analysis of immunization experiments and specific immunity against E7 protein and VACV E3 protein by ELISPOT did not prove any significant differences in the immunogenicity between the parental virus, deletion mutant, and revertants, despite the lower virulence of P13-SigCCI-E7GGG. Lower activity of CC chemokines is probably compensated for by the production of large amounts of extrinsic antigens and direct priming of T cells in secondary lymphoid organs during infection with replicating recombinant VACV (57). Moreover, DCs store antigens on a long-term basis, whereas chemokine sequestering during VACV infection coincides with vCCI production up to 7 days when the virus is eliminated, with the greatest effect seen on day 4, when the virus replicates most. The impact of reduced levels of CC chemokines on the T-cell response is not so significant, probably due to the low production of vCCI from day 5 post-infection.
A comparable number of E7-specific IFN-γ+ CD8+ cells was detected after immunization with P13 E7GGG-SigCCI and with the parental virus P13-E7GGG. However, the therapy of TC-1 tumors was worse. We can speculate that influencing the early phase of innate immunity with smaller quantities of CC chemokines might lead to weaker direct functional activation of NK cells (58).
To explore the influence of the vCCI on the induction of specific immunity in the absence of viral infection, we constructed a DNA vaccine coding the secretory form of vCCI protein. In this case, the production of vSigCCI caused, in comparison with the control plasmid, the induction of weaker specific immunity. The extent of this effect was dependent on the amount of DNA vaccine. The importance of chemokines for the immunogenicity of DNA vaccine was demonstrated for MIP-1α. Co-administration of the plasmid encoding MIP-1α had an adjuvant effect on HIV-1 envelope DNA vaccine (59). We assume that the presence of vCCI at the site of intradermally-administered DNA vaccine against E7 can deeply affect the levels of chemokines, with the greatest impact on chemokines like MIP-1α, which are bound to the highest-affinity sites and are available at low concentrations. This can lead to the suppression of the T-cell response.
In summary, the results of our investigation show that the P13 clone of the attenuated rVACV strain PRAHA does not produce, due to a shortened signal peptide sequence, a functional secretory form of vCCI. This finding correlates with the observed increased levels of the CC chemokines RANTES, MCP-1, MIG, eotaxin, TARC, and MDC in the blood of VACV P13-infected mice. The restoration of a functional form of the vCCI gene reduced the multiplication of the virus in vivo; nevertheless, it did not affect its ability to elicit a T-cell immune response. In comparison to the parental virus, the anti-tumor activity was reduced in the therapeutic arrangement. The co-expression of 35K protein even impaired the immunization capabilities of DNA vaccines. Our results could be helpful in the selection of the parental viruses used for the construction of immunization vectors for therapeutic purposes.
Footnotes
Acknowledgments
We thank Dr. A. Patel (Glasgow) for plasmid pD357 and Dr. T.C. Wu from Baltimore for kindly providing TC-1 cells. We also thank Michal Smahel for help with the production of DNA vaccines. This work was supported by the grants NS-10660-3/2009 IGA and MZ0UHKT2005 from the Ministry of Health of the Czech Republic, and ERDF OPPK CZ.2.16/3.1.00/24001.
Author Disclosure Statement
No conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.