
Abstract
Respiratory syncytial virus (RSV) is a primary cause of severe lower respiratory tract disease in infants, young children, and the elderly worldwide, and despite decades of effort, there remains no safe and effective vaccine. RSV modifies the host immune response during infection by CX3C chemokine mimicry adversely affecting pulmonary leukocyte chemotaxis and CX3CR1+ RSV-specific T-cell responses. In this study we investigated whether immunization of mice with RSV G protein polypeptides from strain A2 could induce antibodies that block G protein–CX3CR1 interactions of both RSV A and B strains. The results show that mice immunized with RSV A2 G polypeptides generate antibodies that block binding of RSV A2 and B1 native G proteins to CX3CR1, and that these antibodies effectively cross-neutralize both A and B strains of RSV. These findings suggest that vaccines that induce RSV G protein–CX3CR1 blocking antibodies may provide a disease intervention strategy in the efforts to develop safe and efficacious RSV vaccines.
Introduction
The explanations for the lack of success in RSV vaccine development are often specific to the vaccine type studied; however, shared challenges also impede RSV vaccine development. First, natural RSV infection does not fully protect from re-infection and provides only partial protection from the disease, suggesting that it will be difficult to induce a protective immune response (21,24,30,53). Second, antigenic and genetic differences among circulating RSV strains are likely sufficient to affect the level of cross-protection induced by viruses from different groups (2,33,43,49). Lastly, the RSV proteins have been shown to circumvent several areas of host immunity (3,7,25,36,50,61 –63).
RSV has been classified into two major groups, A and B, with primary differences between the two groups observed in the G protein (2,43). The RSV G protein was first recognized as an attachment protein that allows binding to the host cell surface through glycosaminoglycans and heparin-binding domains on the G protein (37,65,66). Following RSV infection, the G protein is produced in two forms: a full-length, membrane-anchored polypeptide with a short cytoplasmic domain (Gm), and a secreted, soluble polypeptide (Gs) with the truncated transmembrane domain (31,32,56). The sequence of G protein that is common to both the Gm and Gs forms of the protein consists of a conserved central region containing four cysteine residues, and two variable mucin-like regions flanking the central conserved region (40). Studies have shown that strain-specific antibody responses primarily recognize epitopes within the hypervariable C-terminal region of the RSV G protein (39). Interestingly, the glycosylation pattern of the RSV G protein varies with the infected cell type, indicating that the different glycosylation patterns on the RSV G proteins may be one of the mechanisms used to evade the host immune response via the alteration of the G-protein antigenic profile (9,19,20,47,48). This underscores the importance of the antibody response specific to the central conserved region of the RSV G protein in generating cross-protection against both strains of RSV.
RSV G protein has an important role in modifying the innate and adaptive immune response. Several studies have shown that the chemokine, cytokine, and T-cell response is affected by G-protein expression (25,61,63,64). CX3C chemokine mimicry mediated by the G protein (3,25,36,50,62) is attributed to the central conserved region, which contains four cysteine residues (62). The G-protein central conserved region also has structural homology with the fourth subdomain of 55-kDa tumor necrosis factor receptor (30). Since TNF-α and TNF-β are proinflammatory cytokines affecting the inflammatory response, it is possible that the RSV G protein may bind to TNF-α and/or TNF-β, thereby modulating their antiviral activities during RSV infection (7). Interestingly, infection of mice with a mutant RSV lacking the G and SH genes resulted in higher numbers of natural killer (NK) cells recruited to the lung, as well as increased IFN-γ and TNF-α production, suggesting that the G and/or SH surface proteins inhibit NK cell recruitment and proinflammatory cytokine production (63). Furthermore, studies have found that the G and/or SH proteins inhibit the expression of macrophage inflammatory protein (MIP)-1α, MIP-1β, and MIP-2, and monocyte chemoattractant protein (MCP)-1, which attract NK cells to the lungs in mice (61), and G protein has a CX3C chemokine motif at amino acid positions 182–186, which can bind to the fractalkine receptor CX3CR1 (62). Fractalkine, CX3CL1, functions in the recruitment of CX3CR1-expressing leukocytes, such as subsets of NK cells, CD4+ T cells, and CD8+ T cells, to the sites of infection (25,28,61). CX3CR1 mimicry by the G protein has been shown to facilitate RSV infection and alter CX3CL1 chemotaxis of human and mouse leukocytes to the lungs (3,25,28,50,62). Together these findings suggest that RSV G protein functions to modulate immunity and facilitate virus replication. Thus, disease intervention strategies that interfere with the G protein–CX3CR1 interaction are potentially effective disease intervention strategies.
Although RSV G protein modulates immunity, it also can confer protective immunity similar to the F, or fusion, protein (12,58). Studies evaluating the BBG2Na vaccine candidate showed a clear role for the RSV G protein in protection against RSV disease in animals (8,14,55). Additionally, a study that analyzed serum immunogenicity against RSV G protein epitopes using sera from RSV A- and RSV B-infected human subjects reported a significant increase in the homo- and heterosubtypic IgG response against the central conserved region of the RSV G protein (45). The findings from these studies suggest that antibodies specific to the central conserved region of the RSV G protein may be able to cross-neutralize both A and B strains of RSV and provide cross-protection.
In the present study, we examined an RSV vaccine strategy using RSV G polypeptide vaccination to generate antibodies specific to the central conserved region of the G protein that block the G protein–CX3CR1 interaction of RSV strain A and B viruses, and provide heterosubtypic neutralization of RSV strains A and B. Of note, the RSV A2 G-protein CX3C–CX3CR1 interaction and its relationship to RSV disease are known; however, the RSV B1 G-protein–CX3CR1 interaction and the role it has during RSV infection has not been well documented. Our results show that antibodies specific to the central conserved region of the RSV G protein block binding of RSV A2 and RSV B1 native G protein to CX3CR1, and have some ability to cross-neutralize the two strains of RSV. A better understanding of the host humoral immune response associated with RSV infection is critical for development of safe and efficacious RSV vaccines and therapeutic treatments. This study provides a vaccination strategy to prevent or reduce RSV G-protein-mediated immune evasion and disease pathogenesis.
Materials and Methods
RSV G polypeptides for immunization
RSV G polypeptides spanning different regions of the RSV G protein were designed for vaccination (Fig. 1). G polypeptides were commercially synthesized by Liquid Phase Peptide Synthesis and received as a lyophilized powder (GenScript, Piscataway, NJ). The peptides were resuspended to a stock concentration of 3 mg/mL with DMSO (Sigma-Aldrich, St. Louis, MO) and stored at 4°C. For the studies, the peptides were diluted with PBS, aliquotted at 1 mg/mL, and held at 4°C for general use. For vaccination studies, PBS was used to dilute the polypeptides to the desired concentration. The G polypeptides that were used for immunization correspond to the N-terminal variable glycosylated region (G1 polypeptide, aa 67–147), the central conserved region (G2 polypeptide, aa 148–198), and the C-terminal hypervariable region (G3 polypeptide, aa 199–298) of the G protein of the RSV A2 strain.
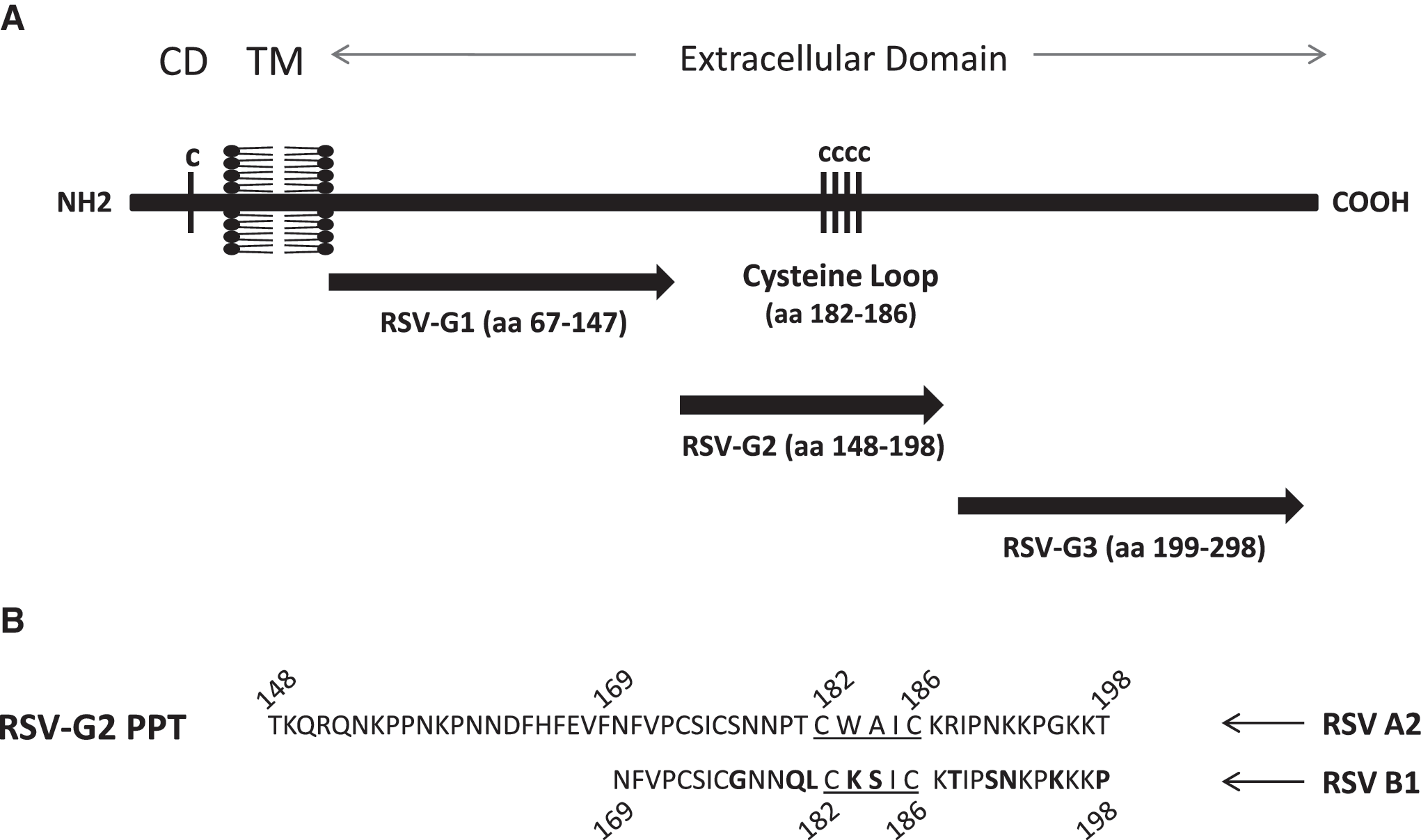
Schematic of RSV G protein and location of G polypeptides. (
RSV native G-protein purification
Native G protein was purified from RSV A2- or RSV B1-infected Vero E6 cells (MOI 1) using affinity chromatography as previously described (Supplementary Fig. S1; see online supplementary material at
G polypeptide vaccination
Four- to six-week-old specific-pathogen-free female BALB/c mice were purchased from Charles River Laboratories, housed in microisolator cages, and fed sterilized water and food ad libitum. The studies were reviewed and approved by the University of Georgia Institutional Animal Care and Use Committee. All RSV G polypeptides and UV-inactivated RSV A2 were emulsified in a 1:1 ratio with TiterMax (Sigma-Aldrich), and mice were immunized intramuscularly (IM) with 50 μg vaccine/mouse in the hindquarters. Each mouse was immunized with 100 μL of the RSV G polypeptide+TiterMax adjuvant mixture (50 μL/hindleg). At day 14 post-vaccination, the mice were boosted with equal an amount of RSV G polypeptide+TiterMax or UV-inactivated RSV A2+TiterMax emulsion. After receiving the boost, vaccinated mice generated an RSV G protein-reactive antibody titer of>3 standard deviations (SD) above background as determined by enzyme-linked immunosorbent assay (ELISA). The sera from the G polypeptide-vaccinated and UV-inactivated RSV A2-vaccinated mice were collected and stored at −80°C for further experiments.
ELISA
The antibody titers in sera collected from vaccinated mice and controls were determined using a modified indirect ELISA (68). Briefly, flat-bottom microtiter plates (Corning, Corning, NY) were coated with 1 μg/well of immunizing antigen, RSV A2 native G protein, or RSV B1 native G protein, and left overnight at 4°C. Serial dilutions of sera in PBS were added to the wells and incubated for 1 h at 37°C. The plates were washed three times with washing buffer (PBS containing 0.05% Tween), and incubated for 1 h at 37°C with alkaline phosphatase-conjugated goat anti-mouse IgG (H+L; Millipore, Temecula, CA). After being washed, the plates were developed with pNpp substrate (Pierce Protein Research Products, Rockford, IL), as indicated by the manufacturer.
Transfection and selection of 293-CX3CR1 cells
Human 293 cells (CRL-1573; ATCC) were transfected with pcDNA3.1 expression plasmids (Invitrogen Corp., Carlsbad, CA) encoding CX3CR1 as previously described (62). Briefly, plasmid inserts were derived from genomic DNA by high-fidelity PCR amplification (Invitrogen), and were sequenced bidirectionally. After G418 selection for at least 3 wk, stable receptor expression was verified by flow cytometry. Stably-transfected cells (293-CX3CR1) were stained with a fluorescein isothiocyanate (FITC)-conjugated anti-CX3CR1 monoclonal antibody (MAb 2A9), obtained from MBL International (Nagoya, Japan). Cell sorting was performed using a Dako Cytomation MoFLo high-speed cell sorter after gating of dead cells by the use of propidium iodide and correction of results for nonspecific staining by the use of isotype antibody controls. The expression level of CX3CR1 was determined by flow cytometry, and showed that >85% of 293-CX3CR1 cells expressed CX3CR1 compared to the untransfected 293 cells.
G protein-CX3CR1 binding inhibition assay
Immunoglobulin G (IgG) was purified from sera of vaccinated mice using immobilized protein G (Thermo Scientific), following the manufacturer's protocol. To evaluate the ability of RSV G polypeptide-specific antibodies to prevent RSV G protein binding to CX3CR1, 1 μg of purified serum IgG antibody was incubated with 1 μM of native G protein purified from either RSV A2 or B1 virus, or with a control peptide (i.e. LH93 polypeptide, INGKWIILLSKF), for 1 h at 4°C. IgG purified from naive mouse serum was used as negative antibody control, and MAb 131-2G was used as positive antibody control in all the assays. 293-CX3CR1 cells and untransfected 293 cells were plated in a round-bottom 96-well plate at 2×105 cells per well, washed with PBS, and incubated with PBS containing anti-human CD32 (Fc block; Millipore) at 1 μg/mL and 4°C for 15 min. After incubation, the cells were resuspended in a pre-incubated mixture of purified IgG and native RSV G protein, and 5 μg/mL of heparin (Sigma-Aldrich) was added to prevent any nonspecific binding, and incubated for 1 h at 4°C. After the incubation, the cells were washed in PBS containing 1% bovine serum albumin (fluorescence-activated cell sorting [FACS] buffer), and incubated with MAb 130-2G conjugated to Alexa-Fluor 488 (AF488; Molecular Probes, Eugene, OR) for 30 min at 4°C. The percentage of G-protein binding to 293-CX3CR1 or 293 cells only was determined by flow cytometry using a BD LSRII and FloJo analysis software. The percent inhibition was calculated using the formula:
RSV plaque reduction assay
The ability of serum antibodies from RSV G polypeptide-vaccinated mice to neutralize RSV A2 or B1 virus was determined by plaque reduction assay. Briefly, 50 μL of 1 μg/mL purified IgG from RSV G polypeptide-specific sera was mixed with RSV A2 or B1 virus that was previously titrated to yield 100 PFU/50 μL/well, and was incubated for 2 h at 37°C. Confluent monolayers of Vero E6 cells were prepared in 24-well plates (Corning), and 100 μL/well of the IgG-RSV mixture was added in triplicate. After virus adsorption for 2 h at 37°C, the cell monolayers were overlaid with 2.0% methylcellulose in MEM with 2% FBS at 100 μL/well. The plates were incubated at 37°C and 5% CO2 for 5 d. At day 5 post-infection, the methylcellulose overlay was aspirated from the plate, the cells were fixed with a 60%/40% methanol-acetone mixture, and the fixed cells were air-dried overnight. The fixed cells were incubated with a mouse MAb specific for RSV F protein (clone 131-2A), followed by a secondary goat anti-mouse IgG antibody conjugated with alkaline phosphatase (KPL, Gaithersburg, MD). The plaques were developed using 200 μL/well of 1-STEP NBT/BCIP (Thermo Scientific) at room temperature for 10 min. The plaques were counted using a dissecting microscope. Titers were calculated from the averages of triplicate experiments.
Statistical analysis
The data are presented as the mean±standard error of the mean (SEM). Student's t-test was used to compare G polypeptide-immunized groups and control groups. p Values<0.01 were considered statistically significant.
Results
Inhibition of fractalkine binding to CX3CR1 in the presence of RSV native G protein
The effects of the RSV G protein-CX3CR1 interaction on immunity and disease pathogenesis has been linked to G-protein binding to the fractalkine receptor, CX3CR1 (62). Specifically, the presence of the RSV G protein CX3C motif has been shown to reduce RSV-specific T-cell responses by inhibiting CX3CL1-mediated trafficking of CX3CR1-expressing immune cells to the site of infection (25), and clinically, a variation in the CX3CR1 gene has been associated with an increased risk for severe RSV bronchiolitis (57). Previous studies have only examined the effects of RSV G protein purified from RSV/A-infected Vero E6 cells (62). To determine if RSV/A and RSV/B G proteins mediate similar activities, and mimic the activities of fractalkine (FKN) for binding to CX3CR1, inhibition of FKN binding to CX3CR1 in the presence of RSV/A2 or RSV/B1 native G proteins was examined (Table 1). The results show that RSV/A2 G protein inhibits FKN binding to CX3CR1 in a dose-dependent fashion, at a level similar to that previously shown (62), and that native RSV/B1 G protein showed similar dose-dependent inhibition of FKN binding to CX3CR1. These results suggest that RSV G protein from A and B strains have similar capacities to mimic the activities of FKN.
293-CX3CR1 cells were incubated with 1 μg, 500 ng, or 250 ng of RSV A2 or RSV B1 native G protein in the presence of 5 μg/mL heparin. The cells were then treated with 1 μg of 5FAM-conjugated fractalkine peptide, and the FKN peptide binding to 293-CX3CR1 cells was determined by FACS analysis. The percent inhibition of FKN peptide binding to 293-CX3CR1 cells by RSV A2 or RSV B1 native G proteins was calculated, and is shown relative to the percent FKN peptide binding on 293-CX3CR1 cells in the absence of RSV native G protein.
Generation of RSV G polypeptide-specific polyclonal antibodies
Three RSV G polypeptides were used to immunize BALB/c mice to generate polyclonal antibodies corresponding to N-terminal variable glycosylated region (G1 polypeptide, aa 67–147), the central conserved region (G2 polypeptide, aa 148–198), and the C-terminal hypervariable region (G3 polypeptide, aa 199–298) of the G protein of RSV strain A2 (Fig. 1). Mice were IM immunized with 50 μg of G1, G2, or G3 polypeptide adjuvanted with TiterMax and equivalently boosted 2 wk later. Three weeks following the second vaccination, sera from vaccinated mice were collected and pooled. Then IgG was purified and quantified from the pooled sera of vaccinated mice in order to normalize IgG levels, and to remove any endogenous CX3CL1 or other serum factors that might affect downstream assays assessing G-protein binding to CX3CR1. IgG antibodies purified from the sera of mice vaccinated with the G2 polypeptide (containing the central conserved region), or inactivated RSV/A2, reacted against both RSV A2 (Fig. 2A) and RSV B1 (Fig. 2B) native G proteins. In contrast, IgG antibodies purified from the sera of mice vaccinated with G1 or G3 polypeptide (comprising the N- and C-terminal variable regions, respectively) failed to recognize RSV A2 (Fig. 2A), and RSV B1 (Fig. 2B) native G proteins relative to negative controls, suggesting that the G2 polypeptide region that encompasses the CX3C motif is antigenic, and can induce IgG antibodies that have some cross-reactivity with both RSV/A2 and RSV/B1 native G proteins.
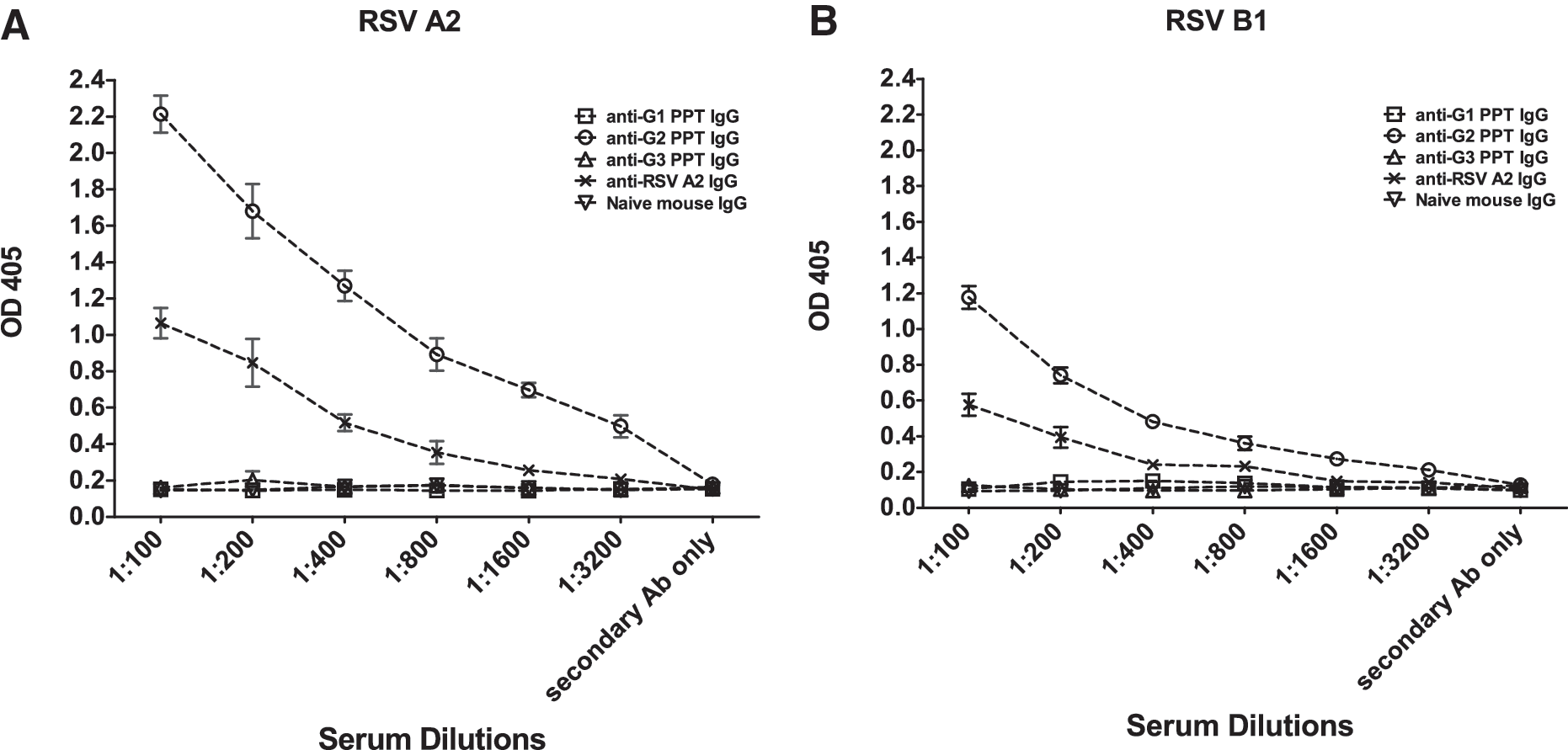
RSV G polypeptide-specific IgG antibodies are reactive against native RSV/A2 and RSV/B1 G proteins. Purified IgG antibodies from the serum of G polypeptide (PPT) G1, G2, G3, or UV-inactivated RSV/A2 mice were evaluated for reactivity to native RSV/A2 (
RSV G polypeptide-specific antibodies inhibit G-protein binding to CX3CR1
Our laboratory previously reported that IgG antibodies purified from the sera of RSV/A2 G polypeptide-immunized mice show various levels of blocking activity of RSV/A2 G protein binding to CX3CR1 (68). It has not been previously determined if antibodies to the G polypeptide vaccines in this study (Fig. 1) block native RSV/A as well as RSV/B1 G-protein CX3C-CX3CR1 binding. To address this, G protein-CX3CR1 binding inhibition assays were performed with either native RSV/A2 (Fig. 3A) or native RSV B1 (Fig. 3B) G protein. Anti-RSV G protein monoclonal antibody (clone 131-2G) reactive to the central conserved region in the G protein (27,29,41), and IgG antibodies purified from naïve BALB/c mice, were used as positive and negative controls, respectively. As expected, treatment of native RSV/A2 G protein with purified IgG from mice immunized with G2 polypeptides significantly (p<0.01) inhibited G-protein binding to CX3CR1 by approximately 40% (Fig. 3A). A similar level of inhibition was observed following treatment of native RSV/A2 G protein with IgG antibodies purified from mice vaccinated with inactivated RSV/A2 virus, and predictably, treatment with monoclonal antibody 131-2G significantly (p<0.01) inhibited G-protein binding to CX3CR1 by approximately 85% (Fig. 3A). Interestingly, IgG from mice immunized with G1 or G3 polypeptides also significantly (p<0.01) inhibited G-protein binding to CX3CR1, by approximately 30% (Fig. 3A). These results suggest that antibodies generated to epitopes at or proximal to the G protein CX3C motif can reduce G-protein binding to CX3CR1. Notably, treatment of RSV/B1 native G protein with purified IgG antibodies from G2 polypeptide-vaccinated mice showed significant (p<0.01) inhibition of G protein binding to CX3CR1; however, treatment with IgG antibodies purified from the mice vaccinated with G1 or G3 polypeptides had no substantial inhibitory effect (Fig. 3B). In addition, IgG from inactivated RSV/A2-infected mice did not significantly inhibit RSV/B1 G-protein binding to CX3CR1 (Fig. 3B). These results indicate that the IgG antibodies generated to the central conserved region of the G protein can cross-react and inhibit G protein-CX3CR1 interaction.
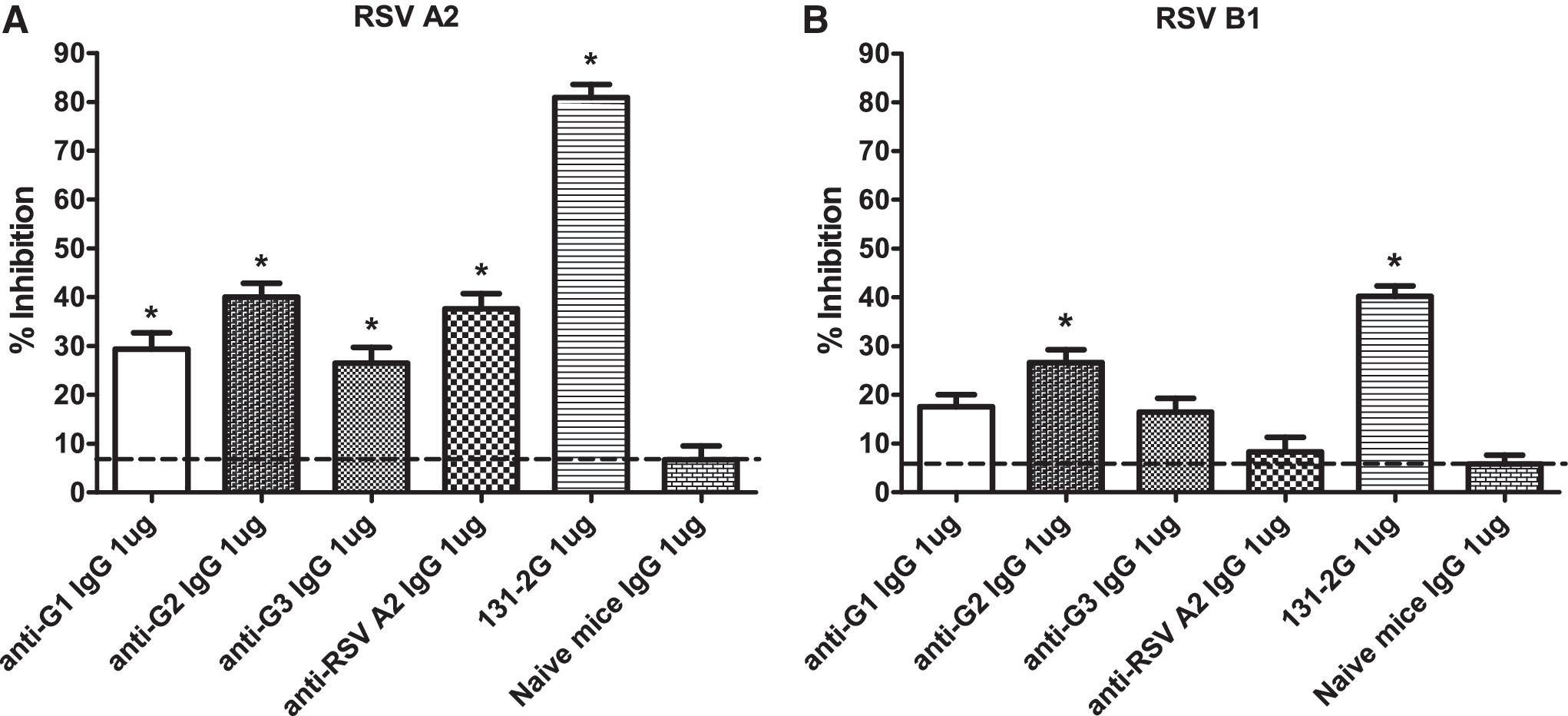
Purified IgG antibodies from G2 polypeptide-vaccinated mice inhibit RSV/A2 and RSV/B1 native G-protein binding to CX3CR1. IgG antibodies purified from the sera of G polypeptide- or UV-inactivated RSV A2-vaccinated mice were examined for their ability to inhibit binding of purified RSV/A2 or RSV/B1 native G protein to 293-CX3CR1 cells. Data represent the percent inhibition of RSV/A2 native G protein (
RSV is neutralized by RSV G polypeptide-specific antibodies
RSV F and G proteins have been shown to induce neutralizing antibodies and confer protective immunity (12,58), and studies that evaluated a BBG2Na vaccine candidate demonstrated a role for RSV G protein in protection against RSV disease (8,14,55). Moreover, a recent study that analyzed the serum from RSV A- and B-infected human subjects reported a substantial homo- and heterosubtypic IgG response to the central conserved region of the RSV G protein (45). To determine the level of antibody neutralization associated with RSV G polypeptide-specific IgG antibodies, purified IgG antibodies specific to G1, G2, or G3 polypeptides, or to inactivated RSV/A2, were tested for inhibition of RSV/A2 (Fig. 4A) or RSV/B1 (Fig. 4B) infection. While IgGs from G1- and G3-vaccinated mice did not exhibit any significant neutralizing antibody response against either strain of RSV, IgG from G2-vaccinated mice showed significant (p<0.01) neutralizing ability against both RSV/A2 (60%) and RSV/B1 (35%) compared to the native IgG treatment. Meanwhile, IgGs from mice vaccinated with the inactivated RSV/A2 showed significant (p<0.01) neutralizing ability against RSV/A2 (80%), but did not show significant neutralization of RSV B1 compared to the native IgG treatment. Taken together, these results indicate that polyclonal IgG antibodies generated following vaccination of BALB/c mice with G2 polypeptide can neutralize RSV/A2, and provide a level of cross-neutralization against RSV/B1.
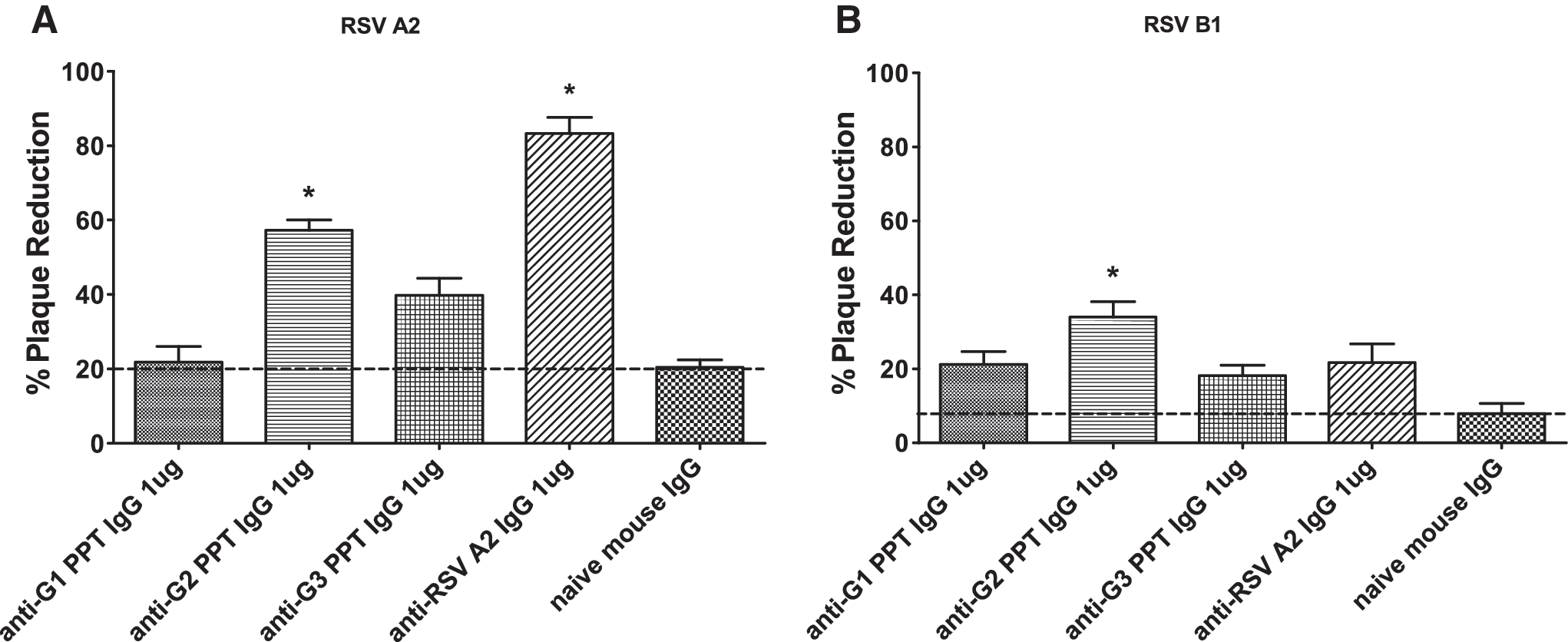
Neutralization of RSV A2 and RSV B1 by IgG antibodies purified from sera of RSV G polypeptide-vaccinated mice. Purified RSV G polypeptide (G1, G2, or G3)-specific IgG antibodies were examined for their ability to neutralize RSV A2 (
Discussion
Despite the significance of RSV as a leading agent of severe lower respiratory tract disease in infants and young children worldwide, there is still no safe and effective vaccination available. This study evaluated RSV G-protein subunit vaccines composed of RSV G polypeptides to induce antibodies that block the G protein–CX3CR1 interaction. RSV is classified into two antigenic groups: A and B, with their differences located in the major attachment G protein (31,32,43). However, both RSV strains share the conserved central region containing the CX3C chemokine motif, which binds to the fractalkine receptor CX3CR1 (62). Interestingly, a previous study has demonstrated that a single nucleotide polymorphism in the CX3CR1 gene is associated with an increased risk for severe RSV bronchiolitis in children hospitalized for bronchiolitis (1). The G protein central conserved region containing the CX3C chemokine motif has been shown to have an important role in the modification of host immune responses during RSV infection, a feature that likely contributes to disease persistence and pathogenesis (25 –28,38,60,62). For example, a previous study from our group has shown that expression of the RSV G protein or the G protein CX3C motif during infection is associated with reduced CX3CR1+ T-cell trafficking to the lung, reduced frequencies of RSV-specific MHC class I-restricted IFN-γ-expressing cells, and lower numbers of IL-4- and CX3CL1-expressing cells (25). Related to the study presented here, we have previously shown that BALB/c mice vaccinated with RSV G-protein peptides containing the CX3C motif generated antibodies that inhibit the RSV/A2 G protein–CX3CR1 interaction, reduce lung virus titers, and prevent body weight loss and pulmonary inflammation (68). These studies underscore the importance of the G protein–CX3CR1 interaction in the modulation of host antiviral responses during RSV infection, and suggest a disease intervention targeted to inhibiting the G protein–CX3CR1 interaction.
In the present study, to determine the ability of the RSV G protein-specific antibodies to inhibit the RSV G protein–CX3CR1 interaction, BALB/c mice were vaccinated with three G polypeptides: G1 polypeptide (aa 67–147), G2 polypeptide (aa 148–198), and G3 polypeptide (aa 199–298), spanning the majority of the RSV G protein. We observed that, although all of the vaccinated mice generated substantial humoral responses against the immunizing peptides, only the serum IgG purified from G2 polypeptide-vaccinated mice strongly reacted with the RSV A2 and RSV B1 native G proteins. The lack of an antibody response against the RSV A2 and RSV B1 native G protein by G1- or G3-polypeptide-vaccinated mice may be explained by the highly glycosylated nature of the N-terminal and C-terminal regions (corresponding to the amino acid sequences represented by the G1 and G3 polypeptides) within the RSV G protein that are flanking the central conserved region. Previous studies have found that the glycosylation pattern of the RSV G-protein hypervariable regions changes with the infected cell type, indicating that the different glycosylation patterns on the RSV G proteins may be one of the mechanisms used to evade the host immune response by altering the G-protein antigenic profile (9,19,20,47,48). It is also possible that the G1 and G3 polypeptides fold or aggregate differently compared to native G protein, and therefore differences may be associated with conformational epitopes that may not be present or are hidden in native G protein.
As we have previously shown, purified IgG antibodies from the sera of RSV G polypeptide-immunized mice have various levels of RSV G protein-CX3CR1 blocking activity for RSV A2 native G protein (68). However, in this study we compared antibody efficacy for inhibiting both RSV A2 and B1 native G-protein binding to CX3CR1, and inhibiting virus replication. Our results show that IgG antibodies purified from the sera of mice vaccinated with G2 polypeptide significantly inhibited RSV A2 and B1 native G protein binding to CX3CR1, and that IgG antibodies purified from the sera of G1- and G3-vaccinated mice also reduced RSV A2 G protein binding to CX3CR1. These results compare with a previous study from our laboratory that demonstrated that IgG antibodies from mice vaccinated with G1, G2, and G3 polypeptides were able to significantly inhibit RSV A2 G-protein-mediated leukocyte chemotaxis, with G2-specific IgG showing the greatest inhibition (68). However, IgG antibodies purified from the sera of G1- and G3-vaccinated mice did not significantly inhibit RSV B1 G-protein binding to CX3CR1. Previous antigenic relatedness studies of RSV G protein as determined by ELISA showed that the F proteins of RSV A and B strains have ∼50% antigenic relatedness, while the G proteins of the two groups are more distantly related, with ∼5% antigenic relatedness (33). In the same study, cotton rats immunized with the recombinant vaccinia virus expressing RSV/A2 G protein showed reduced heterosubtypic protection against RSV B strain compared to the cotton rats immunized with vaccinia virus expressing RSV/A2 F protein (33). Thus, the limited inhibition of RSV B1 G-protein binding to CX3CR1 by G1- and G3-polypeptide-specific IgG antibodies may relate to known differences in the hypervariable regions of the G proteins. However, the lack of glycosylation in the central conserved region of the G protein shared by both RSV/A2 and RSV/B1 provides a target for G2-specific polypeptide IgG antibodies to bind and inhibit its interaction with CX3CR1.
In this study, it was evident that G2 polypeptide vaccination induced the highest level of antibodies reactive to both RSV A2 and B1 G proteins, while inactivated RSV vaccination also raised cross-reacting antibodies to RSV A2 and B1 G proteins (Fig. 2). Despite the fact that anti-G2 polypeptide IgG antibody bound well to native A2 and B1 G proteins, these antibodies did not cross-react effectively with RSV B1 G protein in the ELISA assay, and did not effectively inhibit RSV B1 G-protein binding to CX3CR1 compared to antibodies to RSV/A2 G polypeptides. However, this is not unexpected because the ratio of anti-G, anti-F, and anti-SH protein-specific IgG antibodies raised against inactivated RSV is unknown. It is possible that the presence of anti-F and/or anti-SH antibodies may interfere with the binding of anti-G antibodies to their epitopes, resulting in reduced inhibition of the G protein–CX3CR1 interaction, as was observed in the CX3CR1 binding inhibition assay performed with RSV B1 G protein. Further, anti-G-protein antibodies present in the sera raised against inactivated RSV A2 may be specific to other immunogenic epitopes (possibly conformational epitopes) within the G protein, but not specific to the CX3C motif, thus these antibodies may show reactivity against the RSV B1 G protein in ELISA, while not effectively blocking the RSV B1 G protein–CX3CR1 interaction.
There is evidence that the RSV G protein can provide protection from challenge. A study evaluating a BBG2Na vaccine candidate in combination with different adjuvants and by different routes of administration has shown a role for the RSV G protein in protection against RSV in small and large animals (8,14,55). Furthermore, a study that analyzed the serum immunogenicity against various RSV G epitopes using sera from RSV A- and B-infected human subjects reported a significant increase in homo- and heterosubtypic IgG responses against the central conserved region of the RSV G protein (45). Since the neutralizing ability of antibodies is a good indication of protection against viral pathogens, the neutralizing ability of the RSV G polypeptide-specific IgG antibodies to block the RSV G protein–CX3CR1 interaction was determined using an in vitro plaque reduction assay. The findings in this study showed that the polyclonal IgG antibodies generated from the vaccination of BALB/c mice with G2 polypeptide can effectively cross-neutralize both RSV/A2 and RSV/B1, indicating that the polyclonal IgG antibodies that are specific for the central conserved region of RSV A2 G protein may be able to offer heterosubtypic protection. These findings are consistent with an earlier study showing induction of cross-neutralizing antibodies induced by G protein expressed by recombinant vaccinia virus, which generated cross-neutralizing antibodies against both RSV/A and RSV/B strains (33). In addition, it has been shown that infants and children who are primarily infected with RSV A strain produced significant levels of cross-neutralizing antibodies that could also neutralize RSV B strain (42). Also, when the response to the RSV G protein was examined in the same study, significant levels of RSV B strain G protein-specific cross-neutralizing antibodies were detected in children infected with RSV A strain (42). The central conserved region of the RSV G protein is an immunogenic region that can generate RSV G protein-specific antibodies in humans that can inhibit RSV G protein-mediated leukocyte chemotaxis and interaction of the G protein CX3C motif with the chemokine receptor CX3CR1 (26). The results from this study show that mice vaccinated with RSV G polypeptide containing the CX3C motif in the central conserved region generate IgG antibodies that block RSV A2 and RSV B1 G protein–CX3CR1 interactions and neutralize both the A and B subtypes of RSV. The importance of these findings is that RSV G-polypeptide vaccination can induce antibodies that recognize central conserved G-protein peptide sequences shared by the RSV A2 and B1 strains, and the reactivity of these antibodies can inhibit G-protein binding to CX3CR1 and virus replication, a discovery that has not been described previously, and one that is important in RSV vaccine development
Footnotes
Acknowledgments
This research was supported by the National Institutes of Health (5R01AI69275-4) and the Georgia Research Alliance.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.