
Abstract
Pregnant women can experience high mortality, high rates of abortion, and severe pneumonia when infected with pandemic influenza viruses. In this context, the severity of the 2009 pandemic H1N1 influenza virus compared with seasonal H1N1 influenza virus is not clear. Presently, in a mouse model of pregnancy, the 2009 pandemic H1N1 influenza virus killed up to 60% of pregnant mice and caused abortion in up to 40%, whereas a circulating seasonal H1N1 influenza virus did not cause any deaths or abortions. Higher viral titers and levels of inflammatory cytokines and chemokines such as interleukin (IL)-1α, IL-6, granulocyte colony-stimulating factor, RANTES, monocyte chemotactic protein, and KC (CXCL1), were detected in the lungs of pregnant mice infected with the 2009 pandemic H1N1 influenza virus, compared with the seasonal H1N1 influenza virus. The results of our study with pregnant mice suggest that the observed higher pathogenesis in pregnant women infected with the 2009 pandemic H1N1 influenza virus than the seasonal H1N1 influenza virus may be due to higher viral replication, elevated induction of inflammatory chemokines, and reduced progesterone.
Introduction
In early April 2009, an outbreak by a novel H1N1 influenza virus occurred in the southwestern United States and Mexico, and quickly spread around the world by person-to-person transmission (38,45). In June 2009, the World Health Organization (WHO) raised the pandemic alert to level 6, the highest level of the pandemic warning system. The pandemic virus first appeared in a small village in Vera Cruz, Mexico, with no cases resulting in hospital admission (11). In California, a 10-year-old boy and a 9-year-old girl were infected with the H1N1 influenza virus and were admitted to a hospital for treatment (11). Most countries in the southern and northern hemispheres and tropical areas reported infections of the H1N1 influenza virus (11). The 2009 pandemic H1N1 influenza virus infected more children and young adults than elderly persons (14). In the U.S., the age range of patients infected with the 2009 pandemic H1N1 influenza virus was 3 mo to 81 y, but 60% of patients were younger than age 18 y (8). The high infection rate of young people by the 2009 pandemic H1N1 influenza virus was due to the partial immunity to this virus present in the elderly, since in one survey about 33% of humans over age 60 y possessed cross-reactive antibodies to the 2009 pandemic H1N1 influenza virus (40). The 2009 pandemic H1N1 influenza virus caused a mild illness in the respiratory tract in healthy persons. The clinical signs included fever, cough, sore throat, myalgia, malaise, chills, rhinorrhea, conjunctivitis, headache, shortness of breath, diarrhea, and vomiting (11).
The 2009 pandemic H1N1 influenza virus is genetically distinct from the seasonal influenza viruses circulating in humans, but is closely related to influenza viruses that have been circulating in pigs (7,9,29,46). The hemagglutinin (HA), nucleoprotein (NP), and non-structural protein (NS) genes of the 2009 pandemic H1N1 influenza virus were derived from the classical swine H1N1 influenza virus, the neuraminidase (NA) and matrix (M) genes were from the avian-like Eurasian swine influenza virus, and polymerase acid (PA), polymerase base 1 (PB1), and polymerase base 1 (PB2) were from the North American H3N2 triple-reassortant influenza virus.
Pregnant women are vulnerable to pandemic influenza viruses (13,48). The death rate of pregnant women was 23–71% during the 1918 H1N1 pandemic, and was about 20% during the 1957 H2N2 pandemic (10). During the 2009 H1N1 pandemic, over one-third of infected pregnant women were hospitalized and about 8% died (21,35). The cause for the high mortality rate of pregnant women infected with the pandemic influenza viruses versus the seasonal H1N1 influenza virus may be due to the virulence of the pandemic virus, or the lack of prior immunity to the pandemic virus. It is thought that most pregnant women possess prior immunity to a seasonal influenza virus.
It has been suggested that the excessive production of inflammatory cytokines and chemokines may contribute to the increased disease severity caused by influenza viruses (2,5,6,14,16,17,28,41,45). When macaques were infected with the 1918 pandemic H1N1 influenza virus, the specific groups of genes associated with inflammation and cell death were upregulated in bronchial tissues of infected macaques (6), and the dysregulation of antiviral responses and the higher inductions of inflammatory cytokines were responsible for the severity of disease seen in these animals (16,17). Infection of macaques with the highly pathogenic (HP) H5N1 influenza virus can also induce the sustained expression of type 1 interferons and inflammatory genes (2).
This study was undertaken to investigate whether the 2009 pandemic H1N1 influenza virus was more pathogenic to pregnant mice than a seasonal H1N1 influenza virus, and the reasons for the disparate pathogenesis.
Materials and Methods
Viruses
The seasonal H1N1 influenza virus (A/Brisbane/59/2007) and 2009 pandemic H1N1 influenza virus (A/California/07/2009) were propagated in 10-day-old specific pathogen-free eggs.
Animals
Female BALB/c mice were mated with male BALB/c mice at a 5:1 ratio and 14-day pregnant mice were used to study the pathogenic difference between the 2009 pandemic and seasonal H1N1 influenza viruses. The mice were housed in HEPA-filtered cages in a BSL-3 facility approved by the Korean government. When sera from mice were tested for 2009 pandemic or seasonal H1N1 influenza virus, no mice were positive.
Ethics statement
This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory animals of the Korean veterinary quarantine and service. The protocol was approved by the Committee on the Ethics of Animal Experiments of Chungnam National University. All surgery was performed under Zoletil® anesthesia, and all efforts were made to minimize suffering.
Infection of pregnant and female mice with influenza viruses and observation of clinical signs
Fourteen-day pregnant (n=10) or 5- to 6-week-old non-pregnant female mice (n=10) were intranasally (IN) infected with 50 μL of 105 egg infectious dose 50 (105 EID50/mL) of A/Brisbane/59/2007 or A/California/07/2009, and the infected mice were observed for the survival and abortion rate for 7 days post infection (d.p.i.).
Determination of viral titers in the tissues of infected mice
Fourteen-day pregnant mice (n=10) or 5- to 6-week-old non-pregnant female mice (n=10) were IN infected with 105 EID50/mL of A/Brisbane/59/2007 or A/California/07/2009, and the five surviving mice were euthanized with a lethal dose of Zoletil at 3 and 5 d.p.i. before the lungs were collected. The placental tissues were also collected in the pregnant mice. Lung and placenta tissues (0.5 g) were homogenized in 0.5 mL of phosphate-buffered saline (PBS, pH 7.4), supplemented with 2×antibiotic-antimycotic solution (Sigma-Aldrich, St. Louis, MO). The prepared samples were serially diluted 10-fold in PBS, and each diluted sample was inoculated into 10-day-old fertilized eggs. The presence of virus in the inoculated eggs was determined by a hemagglutination assay using 0.5% turkey red blood cells. Viral titers were determined by log10 EID50/mL as previously described (34).
Histopathology of lungs of infected pregnant and non-pregnant female mice
Infected pregnant or non-pregnant female mice (n=3) were euthanized with a high dose of Zoletil at 5 d.p.i. and the left cranial lobes of the lungs were collected. The lung tissues were fixed in 10% neutral buffered formalin and embedded in paraffin. Five-micron-thick sections were stained with hematoxylin and eosin (H&E) as previously described (1). The stained tissue sections were evaluated using a model DP70 microscope (Olympus, Tokyo, Japan).
Measurement of inflammatory cytokines and chemokines in the lungs of infected pregnant and non-pregnant female mice
Infected pregnant or non-pregnant female mice (n=5) were euthanized with a high dose of Zoletil at 3 or 5 d.p.i. and the left cranial lobes of the lungs were collected. The collected lungs were homogenized in PBS and the supernatants were collected. The collected supernatants were used to determine the induced amount of cytokines and chemokines. Mouse inflammatory cytokines and chemokines were analyzed using a multi-analyte ELISArray kit (SAbioscience, Valencia, CA), and single-analyte ELISA kits (SAbioscience). The assays were performed as indicated by the manufacturer. Briefly, 50 μL of sample dilution buffer was added to each well of an enzyme-linked immunosorbent assay (ELISA) plate prior to the addition of 50 μL of the particular supernatant. The plate was gently shaken and incubated for 30 min at room temperature. The wells were washed with wash buffer and 100 μL of a dilution of the particular detection antibody was added to each well. After incubation for 1 h at room temperature, each well was washed and 100 μL of horseradish peroxidase-conjugated avidin was added to each well. Following incubation for 20 min at room temperature, each well was washed and 100 μL of development solution was dispensed. After incubating for 15 min, 100 μL of stop solution was added to each well. The absorbance of the fluid in each well was read at 450 nm using an ELISA plate reader (Tecan, Männedorf, Switzerland). The amount of the individual cytokine was determined based on the standard curve of each cytokine.
Detection of pregnant hormones in the infected mice
The mouse estrogen and progesterone ELISA kits (MyBioSource, San Diego, CA) were used to evaluate hormone modulation in pregnant and non-pregnant female mice (n=5) infected with 2009 pandemic or seasonal H1N1 influenza virus. Serum and uterus tissue samples were collected from the infected mice 5 d after infection. Uterus tissue samples were diluted in PBS (pH 7.4, 1 g/mL), and were homogenized before they were centrifuged to remove debris at 1000 g for 10 min. The collected supernatants were used to measure the amount of estrogen and progesterone. The assay was performed according to the manufacturer's instructions. Briefly, the supernatants of uterus tissue samples were treated with 10 μL of lysis buffer solution for 1 h. Fifty-microliter samples (serum and homogenized uterus tissue) were added to the wells of a microtiter plate pre-coated with antibody against estrogen or progesterone, and 50 μL of horseradish peroxidase (HRP)-conjugated polyclonal antibody specific for estrogen or progesterone were added prior to incubation at 37°C for 1 h. The plate was washed with the diluted wash solution before 50 μL of substrate A and 50 μL of substrate B were added to each well, and the plates were incubated at room temperature for 15 min. Fifty microliters of stop solution were added to each well of plate, and the optical density (OD) was read at 450 nm using a microtiter plate reader.
Statistical analysis
Statistical significance was determined by the Mann-Whitney U test, and a p value <0.05 was considered significant.
Results
Comparison of clinical signs in pregnant and non-pregnant female mice
Pregnant or non-pregnant female mice were infected with the 2009 pandemic or a seasonal H1N1 influenza virus, and were observed for the occurrence of abortion and mortality. Pregnant mice and non-pregnant female mice infected with the 2009 pandemic H1N1 influenza virus experienced higher rates of mortality and abortion than those infected with the seasonal H1N1 influenza virus. The survival rates of pregnant mice and non-pregnant mice infected with the 2009 pandemic H1N1 influenza virus were 40% and 80%, respectively, while all the pregnant and female mice infected with the seasonal H1N1 influenza virus survived and did not suffer from abortion (Fig. 1A). The loss of body weight was also higher in pregnant mice and non-pregnant female mice infected with 2009 pandemic H1N1 influenza virus than with the seasonal H1N1 influenza virus (Fig. 1B). When we measured the viral titers in the lungs of infected pregnant or non-pregnant female mice, higher viral titers were detected in the pregnant (8.0 EID50/mL) and non-pregnant female (6.5 EID50/mL) mice infected with the 2009 pandemic H1N1 influenza virus, than in the pregnant (6.8 EID50/mL) and non-pregnant female (5.2 EID50/mL) mice infected with the seasonal H1N1 influenza virus at 5 d.p.i. (Fig. 1C). No virus was detected in the placentas of pregnant mice infected with 2009 pandemic or seasonal H1N1 influenza virus (data not shown). Abortions occurred only in pregnant mice infected with the 2009 pandemic H1N1 influenza virus, up to 40% (Fig. 1D).
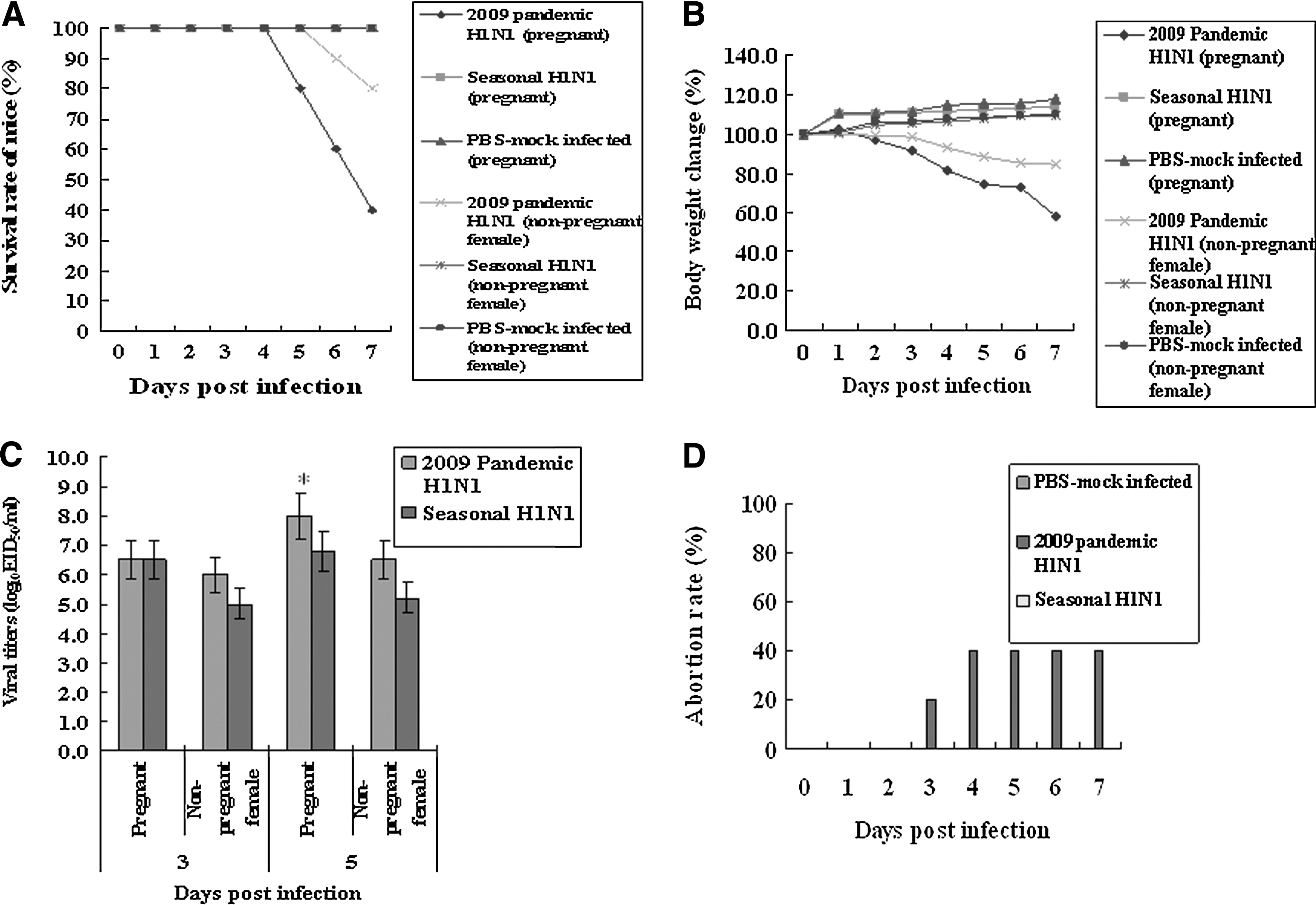
Comparison of the clinical signs and viral titers in the pregnant and non-pregnant female mice infected with 2009 pandemic or seasonal H1N1 influenza virus. Pregnant and non-pregnant female mice (n=10 per group) were IN infected with the 2009 pandemic or seasonal H1N1 influenza virus, and were observed for mortality, body weight change, and abortion, for 7 d after infection. The viral titers (n=5) in the lungs of pregnant mice were determined by log10 EID50/mL. The viral titers were the mean of five mice±standard error. The viral titers with 2009 pandemic H1N1 influenza virus were statistically compared with those of seasonal H1N1 influenza virus (
Comparison of pathological damage to the lungs of pregnant and non-pregnant female mice
To clarify the pathological damage incurred in the lungs of infected pregnant mice, lung tissues collected following infection with the 2009 pandemic or seasonal H1N1 influenza virus at 5 d.p.i. were stained with H&E. The 2009 pandemic H1N1 influenza virus produced severe interstitial pneumonia with mass infiltration of lymphocytes and neutrophils (Fig. 2c), compared to the much milder interstitial pneumonia with some infiltration of lymphocytes and neutrophils seen in the lungs of mice infected with the seasonal virus (Fig. 2d). Lungs of PBS-mock or uninfected pregnant mice showed no interstitial pneumonia (Fig. 2a and b). The bronchial lesions of pregnant mice infected with the 2009 pandemic H1N1 influenza virus showed mucus accumulation and detached cilia, in contrast to those of pregnant mice infected with seasonal H1N1 influenza virus (Fig. 2g and h). The bronchial anatomy of PBS-mock or uninfected pregnant mice was normal (Fig. 2e and f). The alveolar and bronchial lesions of non-pregnant female mice infected with 2009 pandemic or seasonal H1N1 influenza virus (Fig. 2k, l, o, and p) showed milder pathological damage than those of pregnant mice infected with the 2009 pandemic or seasonal H1N1 influenza virus.

Histopathology of lungs of pregnant and non-pregnant female mice infected with the 2009 pandemic or seasonal H1N1 influenza virus. The lungs of pregnant or non-pregnant female mice infected with the 2009 pandemic or seasonal H1N1 influenza virus were collected at 5 d.p.i., and the left cranial lobes were stained with H&E (200×). (
Comparison of inflammatory cytokines and chemokines in the lungs of pregnant and non-pregnant female mice
Pregnant and non-pregnant female mice infected with the 2009 pandemic H1N1 influenza virus showed more severe clinical signs than those infected with the seasonal H1N1 influenza virus. Therefore we measured the mean amounts of inflammatory cytokines and chemokines thought to be responsible for pathogenesis in humans and animals infected with influenza viruses. We first assessed inflammatory cytokines and chemokines in the lungs of infected pregnant mice by simultaneously measuring many cytokine levels (interleukin-1A [IL-1A], IL-1B, IL-2, IL-4, IL-6, IL-10, IL-12, IL-17A, interferon-γ [IFN-γ], tumor necrosis factor-α [TNF-α], granulocyte colony-stimulating factor [G-CSF], and granulocyte macrophage colony-stimulating factor [GM-CSF]), or chemokines (RANTES, monocyte chemotactic protein-1 [MCP-1], macrophage inflammatory protein-1α [MIP-1α], MIP-1β, stromal cell-derived factor-1 [SDF-1], interferon-γ-induced protein-10 [IP-10], monokine induced by gamma interferon [MIG], eotaxin, thymus and activation-regulated chemokine [TARC], macrophage-derived chemokine [MDC], KC[CXCL1], and 6Ckine), to find which cytokines or chemokines would be differentially expressed. IL-1α, IL-6, and G-CSF were more avidly induced in the lungs of pregnant mice infected with 2009 pandemic H1N1 influenza virus than mice infected with the seasonal H1N1 influenza virus (Fig. 3A). The mean quantities of IL-1α induced in the lungs of pregnant mice infected with the 2009 pandemic H1N1 influenza virus and seasonal virus were 2746.091±247.148 pg/mL and 1241.842±93.138 pg/mL, respectively, at 3 d.p.i. (p<0.05), and the levels of IL-1α induced in the lungs of non-pregnant females infected with the 2009 pandemic H1N1 influenza virus and the seasonal virus were 2446.135±234.371 pg/mL and 1141.532±73.241 pg/mL, respectively, at 3 d.p.i. (p<0.05; Fig. 3B). The mean amounts of induced IL-6 and G-CSF in the lungs of pregnant mice infected with the 2009 pandemic H1N1 influenza virus were 4482.365 pg/mL and 3996.40 pg/mL at 3 d.p.i., respectively (p<0.05), while those in the lungs of pregnant mice infected with the seasonal H1N1 influenza virus were 1040.6 pg/mL and 1040.72 pg/mL at 3 d.p.i., respectively. The induced levels of IL-6 and G-CSF in the lungs of non-pregnant females infected with the 2009 pandemic H1N1 influenza virus were 4281.267 pg/mL and 3471.32 pg/mL at 3 d.p.i., respectively (p<0.05), while those in the lungs of non-pregnant females infected with the seasonal H1N1 influenza virus were 1010.2 pg/mL and 1008.53 pg/mL at 3 d.p.i., respectively (Fig. 3C and 3D).

Detection of inflammatory cytokines in the lungs of pregnant and non-pregnant female mice infected with 2009 pandemic or seasonal H1N1 influenza virus. The lungs of pregnant and non-pregnant female mice (n=5 per group) infected with 2009 pandemic or seasonal H1N1 influenza virus were collected at 3 or 5 d.p.i. Inflammatory cytokines in the homogenized lungs were detected using a multi-inflammatory cytokine kit (
When we measured mean chemokine levels in the lungs of pregnant and non-pregnant female mice infected with the 2009 pandemic or seasonal H1N1 influenza virus, RANTES, MCP-1, and KC(CXCL1) (keratinocyte chemoattractant), which are involved in chemotaxis and cell activation of neutrophils, were differentially expressed in the lungs for the two viruses (Fig. 4). RANTES was more avidly induced in the lungs of pregnant mice infected with the 2009 pandemic H1N1 influenza virus (1231.0 pg/mL and 1488.0 pg/mL at 3 and 5 d.p.i., respectively; p<0.05), compared with seasonal H1N1 influenza virus (908.0 pg/mL and 745 pg/mL at 3 and 5 d.p.i., respectively), and RANTES was also more avidly induced in the lungs of non-pregnant females infected with the 2009 pandemic H1N1 influenza virus (1028.0 pg/mL and 1173.0 pg/mL at 3 and 5 d.p.i., respectively; p<0.05), compared to the seasonal H1N1 influenza virus (612.0 pg/mL and 542 pg/mL at 3 and 5 d.p.i., respectively; Fig. 4B). MCP-1 and KC were also more highly induced in the lungs of pregnant and non-pregnant female mice infected with the 2009 pandemic H1N1 influenza virus than in the lungs of pregnant mice infected with the seasonal H1N1 influenza virus (Fig. 4C and D). The induced mean amounts of MCP-1 in the lungs of pregnant mice infected with the 2009 pandemic H1N1 influenza virus were 3349.0 pg/mL and 2994.0 pg/mL at 3 and 5 d.p.i., respectively (p<0.05), and those in the lungs of pregnant mice infected with the seasonal H1N1 influenza virus were 1306.0 pg/mL and 1348.0 pg/mL at 3 and 5 d.p.i., respectively. The levels of MCP-1 in the lungs of non-pregnant females infected with the 2009 pandemic H1N1 influenza virus were 3237.0 pg/mL and 2672.0 pg/mL at 3 and 5 d.p.i., respectively (p<0.05), and those in the lungs of non-pregnant females infected with the seasonal H1N1 influenza virus were 1201.0 pg/mL and 1121.0 pg/mL at 3 and 5 d.p.i., respectively (Fig. 4C). The induced amounts of KC in the lungs of pregnant mice infected with the 2009 pandemic H1N1 influenza virus were 1423.0 pg/mL and 1054.0 pg/mL at 3 and 5 d.p.i., respectively (p<0.05), while those in the lungs of pregnant mice infected with the seasonal H1N1 influenza virus were 764.0 pg/mL and 736.0 pg/mL at 3 and 5 d.p.i., respectively, and those of KC in the lungs of non-pregnant females infected with the 2009 pandemic H1N1 influenza virus were 1231.0 pg/mL and 951.0 pg/mL at 3 and 5 d.p.i., respectively (p<0.05), while those in the lungs of non-pregnant females infected with the seasonal H1N1 influenza virus were 652.0 pg/mL and 532.0 pg/mL at 3 and 5 d.p.i., respectively (Fig. 4D). These results suggest that the higher induction of inflammatory cytokines and chemokines in the lungs may be among the factors causing more severe symptoms in pregnant mice infected with the 2009 pandemic H1N1 influenza virus.
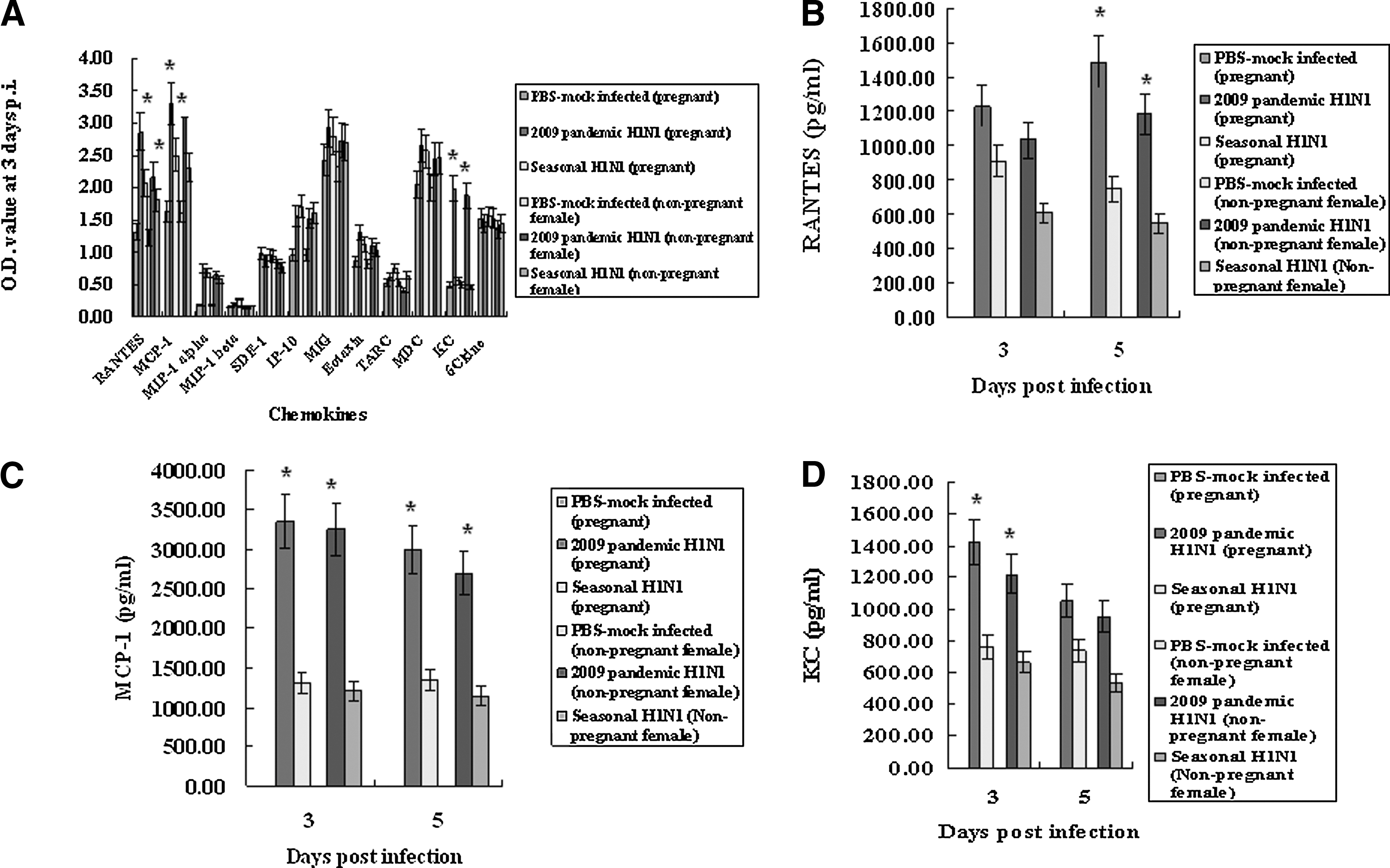
Detection of chemokines in the lungs of pregnant and non-pregnant female mice infected with 2009 pandemic or seasonal H1N1 influenza virus. The lungs of pregnant and non-pregnant female mice (n=5) per group infected with 2009 pandemic or seasonal H1N1 influenza virus were collected at 3 or 5 d.p.i. The chemokines in the homogenized lungs were detected with a multi-chemokine kit (
Detection of hormones in the infected pregnant and non-pregnant female mice
We measured the amount of progesterone and estrogen in the pregnant and non-pregnant female mice in serum and uterus samples collected 5 d.p.i. The amount of progesterone found in serum and uterine samples of the pregnant mice infected with the 2009 pandemic H1N1 influenza virus was lower than that found in the pregnant mice infected with the seasonal H1N1 influenza virus, and was similar to that found in non-pregnant female mice (Fig. 5A). The amounts of estrogen found in the serum of pregnant mice infected with the 2009 pandemic or seasonal H1N1 influenza viruses were lower than those of PBS-mock infected mice, but those found in uterine samples were similar among infected and PBS-mock infected mice (Fig. 5B). The estrogen levels in non-pregnant female mice were similar in uterine tissues and serum samples for all groups (Fig. 5B). These results suggest that the reduced progesterone levels seen in the pregnant mice infected with the 2009 pandemic influenza virus may account for the higher abortion rates than those of the pregnant mice infected the seasonal H1N1 influenza virus.
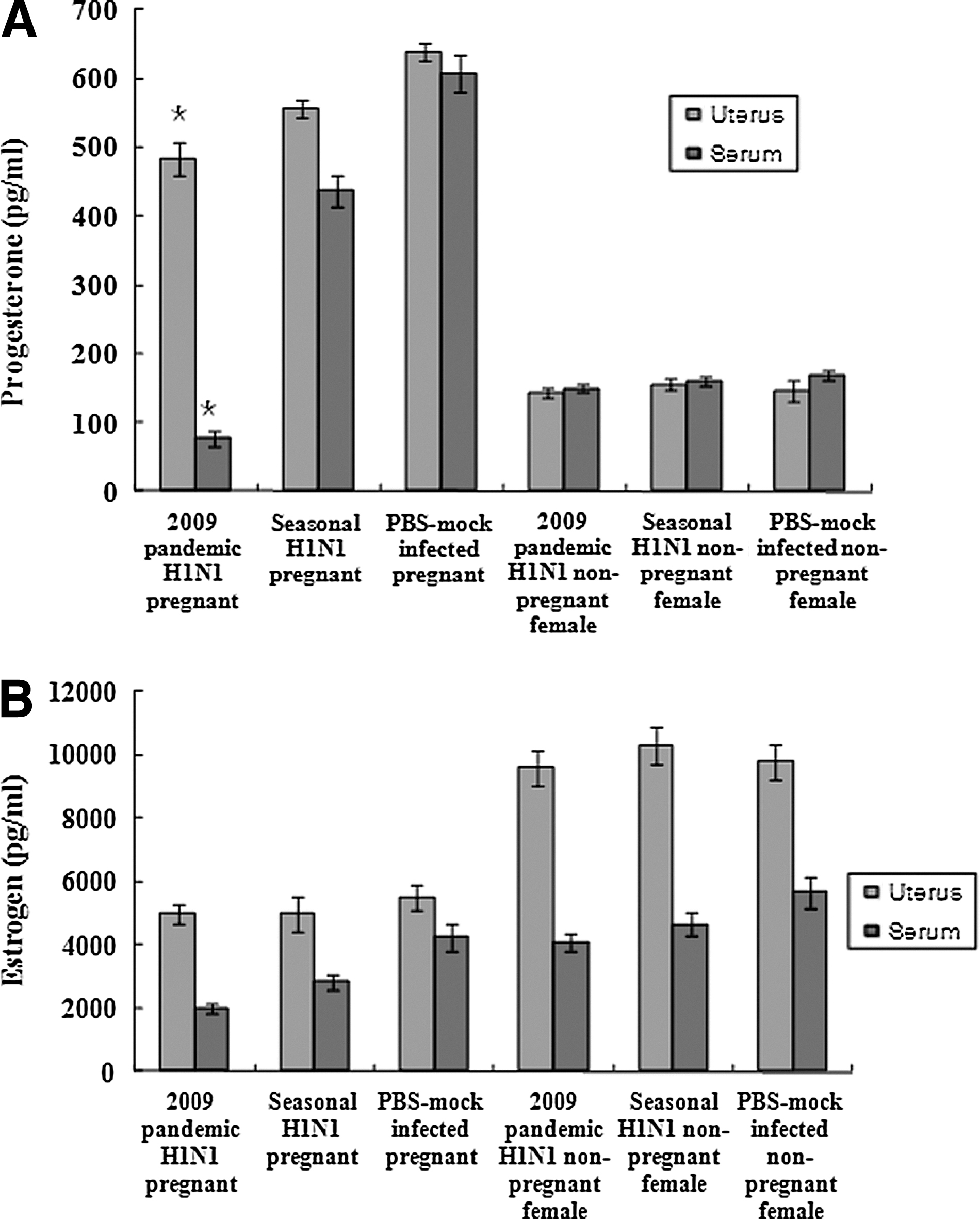
Detection of hormones in infected pregnant and non-pregnant female mice. Serum and uterine tissue samples were collected from pregnant and non-pregnant female mice infected with 2009 pandemic or seasonal H1N1 influenza virus 5 d after infection. Uterine tissue samples were diluted in PBS (pH 7.4) and were homogenized before they were used for the assay. ELISA kits for mouse progesterone (A) and estrogen (B) were used for measuring the induced amount of these hormones in the infected pregnant mice. Data were the mean of five mice±standard errors. The data for the 2009 pandemic H1N1 influenza virus were statistically compared with those of the seasonal H1N1 influenza virus (*p<0.05).
Discussion
Pandemic influenza viruses cause more severe disease in pregnant women than in non-pregnant women, since they are immunocompromised due to childbearing (19). The 2009 pandemic H1N1 influenza virus was also responsible for high mortality in pregnant women. However, it is not clear whether the 2009 pandemic H1N1 influenza virus is more pathogenic to pregnant women than a seasonal H1N1 influenza virus. Our study with a mouse model of pregnancy demonstrates that the 2009 pandemic H1N1 influenza virus does indeed produce higher rates of mortality and abortion in pregnant mice than a seasonal H1N1 influenza virus. Higher viral titers and elevated inflammatory cytokines and chemokines were detected in the pregnant mice infected with the 2009 pandemic H1N1 influenza virus than a seasonal H1N1 influenza virus.
Our results suggest that the pregnant mice could be more vulnerable to infection caused by the 2009 pandemic H1N1 influenza virus than to the infections caused by a circulating seasonal H1N1 influenza virus. Two previous studies also investigated pathogenesis in pregnant mice infected with the 2009 pandemic H1N1 influenza virus (4,24). These studies compared the pathogenesis between pregnant and healthy mice. In contrast, our study was designed to address the pathogenesis of the 2009 pandemic H1N1 influenza virus in pregnant mice compared to a seasonal H1N1 influenza virus in pregnant mice. Our study demonstrates that the 2009 pandemic H1N1 influenza virus killed up to 60% of pregnant mice, while no pregnant mouse infected with a seasonal H1N1 influenza virus died. This result implies that the higher mortality of pregnant women infected with the 2009 pandemic H1N1 influenza virus may be due to a difference in the virus rather than to immunity, since most pregnant women would be expected to have seasonal exposure to H1N1 influenza virus, and since we used specific pathogen-free mice that do not have any immunity to either the 2009 pandemic or seasonal H1N1 influenza virus.
It is noteworthy that seasonal influenza viruses also cause severe disease in pregnant woman (30). A previous study showed that 25 out of 10,000 women in their third trimester were hospitalized due to influenza-related symptoms. Our study showed that the seasonal influenza virus did not cause severe clinical signs such abortion and mortality in pregnant mice. This discrepancy may be due to the different subtypes of influenza viruses that were involved in causing the disease. We used only H1N1 influenza virus, but pregnant women may have been infected with H1N1, H3N2, and human B influenza viruses.
Viral titers in the lungs of pregnant mice infected with the 2009 pandemic H1N1 influenza virus were higher than those in the lungs of pregnant mice infected with a seasonal H1N1 influenza virus. This is consistent with the view that the 2009 pandemic H1N1 influenza virus may be better able to replicate in the lungs of pregnant mice than a seasonal H1N1 influenza virus, and that higher viral titers might contribute to higher mortality of pregnant mice infected with 2009 pandemic H1N1 influenza virus. Previous studies using non-pregnant mice and ferrets also showed that the 2009 pandemic H1N1 influenza virus grew more vigorously in the respiratory tracts of these animals than a seasonal H1N1 influenza virus (12,23,27). In ferrets, the 2009 pandemic H1N1 influenza virus replicated well in the trachea, bronchi, and bronchioles, compared with seasonal H1N1 influenza virus (27,23). Itoh et al. (12) also showed that the 2009 pandemic H1N1 influenza virus could replicate in mice, ferrets, and primates, better than a currently circulating human H1N1 influenza virus.
Our data indicate that the elevated induction of inflammatory cytokines and chemokines such as IL-1, IL-6, G-CSF, RANTES, MCP, and KC contribute to higher mortality and abortion rates in pregnant mice infected with the 2009 pandemic H1N1 influenza virus than a seasonal H1N1 influenza virus. IL-1 is a potent proinflammatory cytokine involved in the activation of lymphocytes, the infiltration of leukocytes at sites of infection, and fever and anorexia (22). IL-6 and G-CSF can stimulate the production of neutrophils and modulate the function, activity, and development of mature neutrophils (37,42). The RANTES, MCP-1, and KC chemokines are chemotactic for monocytes, basophils, eosinophils, and neutrophils at the site of infection (26,31). Induced inflammatory cytokines and chemokines contribute to the pathogenesis of influenza virus (2,32). In mice, infection with either the 1918 pandemic H1N1 influenza virus or the HP H5N1 influenza virus is manifest as severe lung pathology with high induction of proinflammatory cytokines in the lungs (32). A study with HP H5N1 influenza virus in human primary alveolar and bronchial epithelial cells reported higher induction of chemokines and cytokines such as IP-10, interferon-β, RANTES, and IL-6, compared with a seasonal H1N1 influenza virus (5). Infections of mice with a reassorted influenza virus expressing HA from the 1918 pandemic H1N1 influenza virus can induce high levels of macrophage-derived chemokines and cytokines (16,17). Furthermore, non-human primates infected with HP H5N1 influenza virus can develop severe bronchiolar and alveolar lesions with the dramatic and sustained expression of type I interferons and inflammatory and innate immune genes (2).
In conclusion, the 2009 pandemic H1N1 influenza virus is more pathogenic to pregnant mice than a seasonal H1N1 influenza virus, indicating that the higher mortality rate of pregnant women caused by the 2009 pandemic H1N1 influenza virus may be due to the pathogenic nature of this virus, and was not due to a lack of pre-existing immunity against this pandemic virus.
Footnotes
Acknowledgments
This study was financially supported by the research fund of Chungnam National University in 2010. The staff of HARRISCO, an English editing company, edited this manuscript.
Author Disclosure Statement
No competing financial interests exist.