
Abstract
Our aim was to investigate the effects of interferons (IFNs)-λ (interleukin-29 [IL-29], IL-28A, and IL-28B) on hepatitis C virus (HCV) and hepatitis A virus (HAV) internal ribosomal entry site (IRES)-mediated translation. The effects of these IFNs on HCV/HAV translation from HAV/HCV IRES were investigated using bicistronic reporter constructs. We transfected HCV/HAV IRES constructs into these IFN-expressing cell lines. IL-29 showed stronger inhibition of their IRES-mediated translation. Combining IL-29 with IFN-α or amantadine resulted in stronger inhibition of HAV IRES activity. Our findings demonstrated a novel antiviral effect of IFNs-λ against HAV and HCV through the suppression of IRES-mediated translation.
Introduction
Hepatitis C virus (HCV), an enveloped RNA virus of the Flaviviridae family, causes a spectrum of diseases ranging from an asymptomatic carrier state to end-stage liver disease, including cirrhosis and hepatocellular carcinoma (HCC) (1,14,16). HCV has a 5′ NTR, a long open reading frame, and a 3′ NTR. The HCV genome is approximately 9600 nt in size and encodes a polyprotein precursor of about 3000 amino acids, which is cleaved by both viral and host proteases into structural (core, E1, E2, and p7) and nonstructural (NS2, NS3, NS4A, NS4B, NS5A, and NS5B) proteins. HCV also has IRES containing a 5′ NTR and part of the core coding region which forms a secondary structure, and supports translation initiation of an HCV genome in a cap-independent manner (18,24).
IFNs-λ are the most recently defined members of the class III cytokine family, consisting of IL-28A (IFN-λ2), IL-28B (IFN-λ3), and IL-29 (IFN-λ1), and a component of their receptor, IL28-Rα. IL-28 and IL-29 represent an evolutionary link between type I IFNs and the IL-10 family (39). This receptor-ligand system might contribute to antiviral or other defenses by a mechanism similar to, but independent of, type I IFN (25). Additional study is necessary to determine whether IFN-λ can synergize with IFN-α in viral infections, or whether it plays an independent primary role in the antiviral system (41).
There are several reports that IFN-λ exerts antiviral activity against HBV (36), HCV (36), West Nile virus (WNV) (29), influenza A virus, influenza B virus, respiratory syncytial virus, human metapneumovirus, and severe acute respiratory syndrome (SARS) coronavirus (34,41). Human hepatocytes express the IFN-λ receptor complex and IFNs-λ induce signal transducer and activator of transcription 1 (STAT1) phosphorylation (5). Activation of STAT1 is an important factor for the eradication of HCV after antiviral treatments (33). In addition, cellular proteins known as IRES trans-acting factors (ITAFs) are also required for efficient IRES-mediated translation (23,26). The subset of ITAFs that regulates translation initiation appears to be specific for each IRES element (23,26). IFNs including IFNs-λ affect host factors (21,27).
We reported that HAV IRES is an attractive target of anti-HAV drugs because IRES is located in the 5′ NTR, the most conserved region among different HAV strains (19,20,45), and is well conserved among clinical isolates (15,17). It has also been reported that HCV IRES is an attractive target of anti-HCV drugs because IRES is located in the 5′ NTR, the most conserved region among different HCV strains (18,24). We focused on the IRES as an antiviral target of these viruses because IRES activity seems to be correlated with translation of viral protein, which is important for viral replication, although there are contrary opinions. Recently, several groups reported that IL-28B SNP predicts hepatitis C treatment-induced viral clearance and natural clearance (10,16,36,42). In the present study, we examined the inhibitory effects of IFN-λ against HAV and HCV IRES-mediated translation.
Materials and Methods
Cells and virus
Huh7 cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal calf serum (FCS) at 37°C and 5% CO2. The plasmids pcDNA3.1-IL28A, pcDNA3.1-IL28B, and pcDNA3.1-IL29 [kindly provided by Prof. T. Betakova, Institute of Virology, Slovak Academy of Sciences, Bratislava, Slovak Republic (41)] were plasmids expressing IL-28A, IL-28B, and IL-29, respectively. Huh7 cells were transfected with the expression plasmids pcDNA3.1-IL28A, pcDNA3.1-IL28B, pcDNA3.1-IL29, or pcDNA3.1 in Effectene transfection reagent (Qiagen, Hilden, Germany). After 48 h, G418 (Promega, Madison, WI) was added at 1000 μg/mL for selection of Huh7-IL28A, Huh7-IL28B, Huh7-IL29, or Huh7-pcDNA3.1. After 3 wk, to avoid monoclonal selection, all cells were collected for further analysis.
Cell culture-grown HCV JFH1 (genotype 2a) was used in Huh7-derived cell lines (14,43). Tissue culture-adapted HAV strain KRM003 (genotype IIIB) was used in African green monkey kidney GL37 cells (22,37). A HAV DNA-based subgenomic replicon [kindly provided by Prof. V. Gauss-Muller, Institute of Virology, University of Luebeck, Luebeck, Germany (9)] was also used in HuhT7 cells that stably express T7-RNA polymerase in cytoplasm (9,38).
Reagents
The chemicals used were human recombinant IFN-α (Sigma-Aldrich, St. Louis, MO), human recombinant IL29 (IFN-λ1; Acris Antibodies GmbH, Herford, Germany), and amantadine hydrochloride (Sigma-Aldrich).
Bicistronic reporter plasmids
The bicistronic plasmid pSV40-HAV IRES-luc, encoding Renilla reniformis luciferase (Rluc) and firefly luciferase (Fluc), was separated by HAV IRES derived from pHM175 (kindly provided by Prof. S.U. Emerson, U.S. National Institutes of Health) under the control of SV40 promoter, with a polyadenylation signal (polyA; Fig. 1A) (19). The bicistronic plasmid pSV40-HCV IRES-luc (kindly provided by Prof. M. Kruger, Medizinische Hochschule Hannover, Hannover, Germany), carries the Rluc gene, the HCV IRES, including the full-length HCV core, and the Fluc gene under control of the SV40 promoter, with a polyadenylation signal (polyA; Fig. 1B) (18,24).
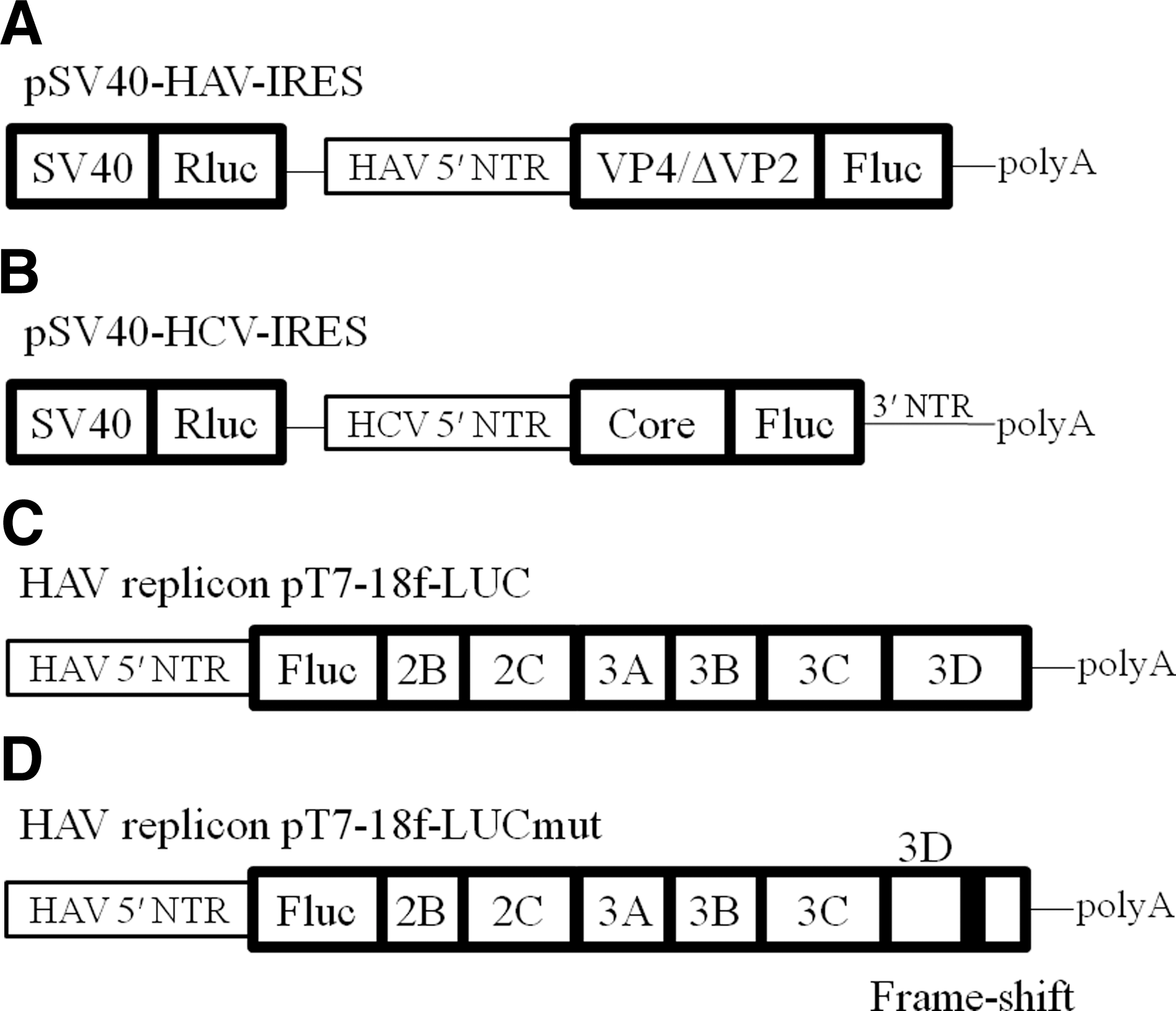
Structure of HAV/HCV reporter constructs used in this study. (
Transfection and in vitro reporter assays
Approximately 1.0×105 cells per well were placed in a six-well plate (Iwaki Glass, Tokyo, Japan) 24 h prior to transfection. The cells were transfected with 0.4 μg of pSV40-HAV (HCV) IRES-luc using Effectene Transfection Reagent (Qiagen) following the manufacturer's protocol. Forty-eight or 72 h after transfection, the cells were harvested using reporter lysis buffer (Toyo Ink, Tokyo, Japan), and luciferase activity was determined by luminometer (Luminescencer-JNR II AB-2300; ATTO, Tokyo, Japan) (44). To control for variations in transcription, IRES activity was assessed by measuring the ratio of Rluc and Fluc activities. The relative ratio of Fluc activity to Rluc activity (Fluc:Rluc) was defined as 100% in the untreated condition. We accept more than 102 of Fluc/Rluc as being positive for IRES activity. All samples were run in triplicate.
RNA extraction, cDNA synthesis, and RT-PCR
The cells were seeded into 6-well plates, and total cellular RNA was extracted 48 h later with the RNeasy Mini Kit (Qiagen) according to the manufacturer's instructions. The RNA samples were then stored at −80°C until use. RNA quality was examined using the A280:A260 ratio (Pharmacia Biotech, Bedford, MA). cDNA synthesis was performed with a random hexamer using Prime Script reverse transcriptase (Takara Bio Inc., Otsu, Shiga, Japan). For detection of ectopic expression of IL-28A, IL-28B, and IL-29, RT-PCR was performed with a Thermal Cycler (TP3000; Takara Bio Inc.) using PrimeSTAR HS DNA polymerase (Takara Bio Inc.) with primers as previously described (41), together with primers for GAPDH (44).
Enzyme-linked immunosorbent assay (ELISA) for IL-28A and IL-29
Cell culture fluid was analyzed for human IL-28A and for IL-29 by ELISAs (R&D Systems, Minneapolis, MN and eBioscience, San Diego, CA, respectively), following the manufacturers' protocols. Briefly, cell culture fluid samples were incubated in plates at 37°C overnight, followed by incubation with biotinylated monoclonal antibodies. Avidin-conjugated peroxidase was added to the plates, and enzyme activity was detected with a Bio-Rad iMark microplate reader (Bio-Rad, Hercules, CA) (44). The sensitivities of human IL-28A and IL-29 by these ELISA kits were 8 pg/mL and 12.5 pg/mL, respectively.
MTS assay
To evaluate cell growth and cell viability, dimethylthiazol carboxymethoxyphenyl sulfophenyl tetrazolium (MTS) assays were performed with the CellTiter 96 Aqueous One-Solution cell proliferation assay (Promega).
Statistical analysis
Data were expressed as mean±SD. Statistical analysis was done by Student's t-test. A value of p<0.05 was considered significant.
Results
Overexpression of IL-28A, IL-28B, or IL-29, in Huh7 cells
We used three protein plasmid vectors under control of the CMV promoter: pcDNA3.1-IL28A, pcDNA3.1-IL28B, and pcDNA3.1-IL29 (41). We established three IFN-λ-overexpressing Huh7 cells, designated as Huh7-IL28A, Huh7-IL28B, and Huh7-IL29. We also used pcDNA3.1 for the establishment of a control cell, Huh7-pcDNA3.1. For the generation of stable cell lines, Huh7 cells were transfected with these vectors and treated with G418. Antibiotic-resistant colonies were expanded for further analysis. To test the ability of these cells to express IL-28A and IL-29, we detected these mRNAs by RT-PCR and measured these cytokines by ELISA (Fig. 2A, upper and lower panels). IL-28A or IL-29 mRNAs were detected only in Huh7-IL28A or Huh7-IL29 cells, respectively. IL-28A or IL-29, respectively, could be measured in each cell culture fluid of Huh7-IL28A or Huh7-IL29. We confirmed the expression of IL-28B mRNA in the cellular RNA of Huh7-IL28B since we could not use ELISA for IL-28B at this time (Fig. 2A, upper panel). We next examined whether overexpression of IFN-λ had any effect on cell proliferation. Equal numbers of control Huh7-pcDNA3.1 and IFN-λ-overexpressing Huh7 cells (Huh7-IL28A, Huh7-IL28B, and Huh7-IL29) were plated, and cell viability was counted at 24 h by MTS assay (Fig. 2B). There were no differences in cell viabilities among these cell lines.
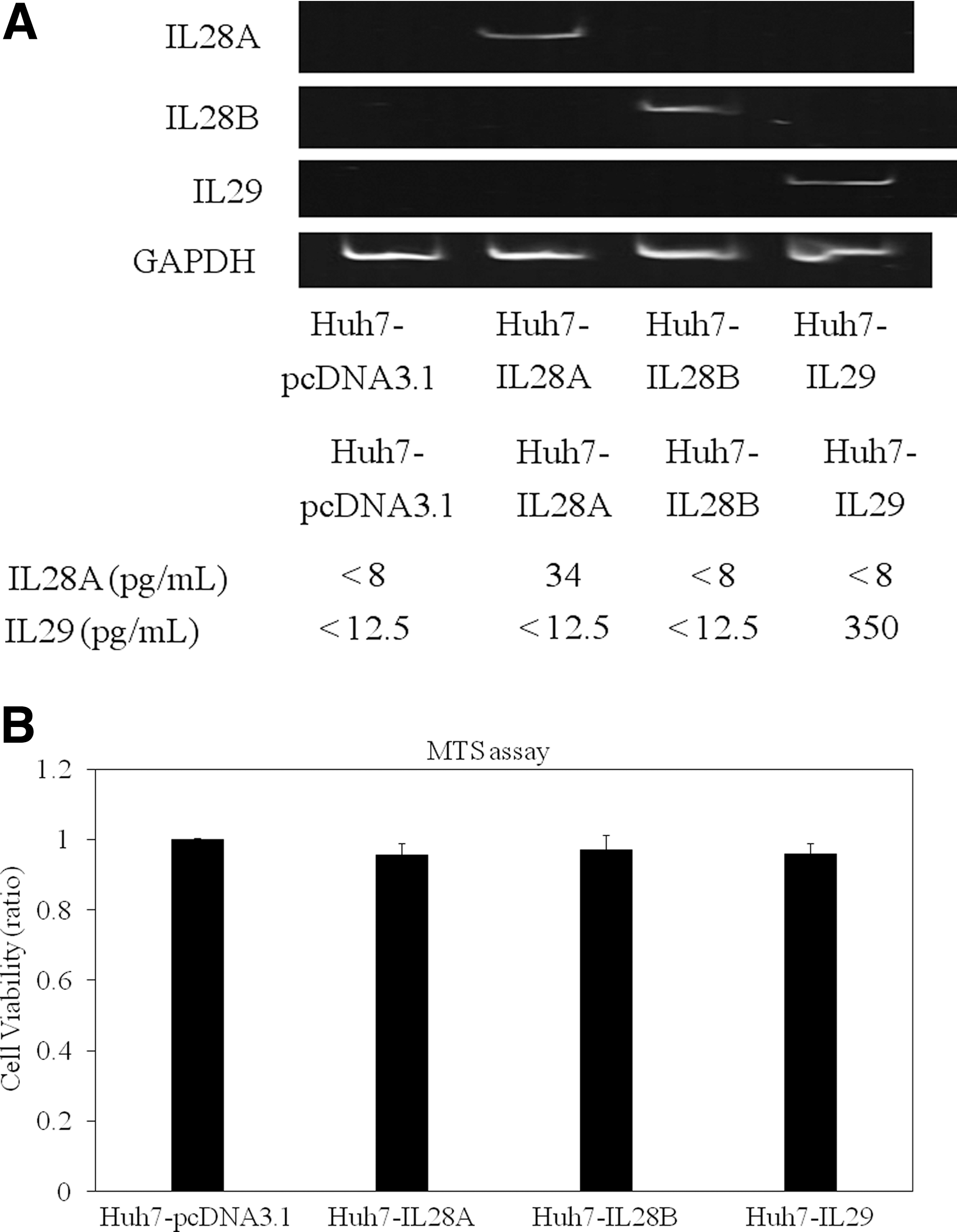
Overexpression of IL-28A, IL-28B, or IL-29 in human hepatoma cell line Huh7. (
HAV IRES activity in Huh7-IL28A, Huh7-IL28B, and Huh7-IL29
Next, we examined the effects of these IFNs-λ on HAV IRES-mediated translations using a luciferase reporter assay. Huh7-IL28A, Huh7-IL28B, Huh7-IL29, and Huh7-pcDNA3.1 were transfected with pSV40-HAV IRES reporter vector encoding SV40 promoter driven-Rluc and Fluc, separated by HAV IRES (19), and 48 h later, luciferase activity was measured and IRES activity was determined (Fig. 3A). HAV IRES activity was inhibited in Huh7-IL28A (56.7%; n=3, p<0.0001), Huh7-IL28B (95.3%, n=3, p=0.0021), and Huh7-IL29 (14.9%, n=3, p<0.0001), compared to that in control Huh7-pcDNA3.1 (n=3, 100%). IL-28A and IL-28B demonstrated inhibitory effects on HAV IRES activity, but they seemed less efficient than IL-29 (Fig. 3A).
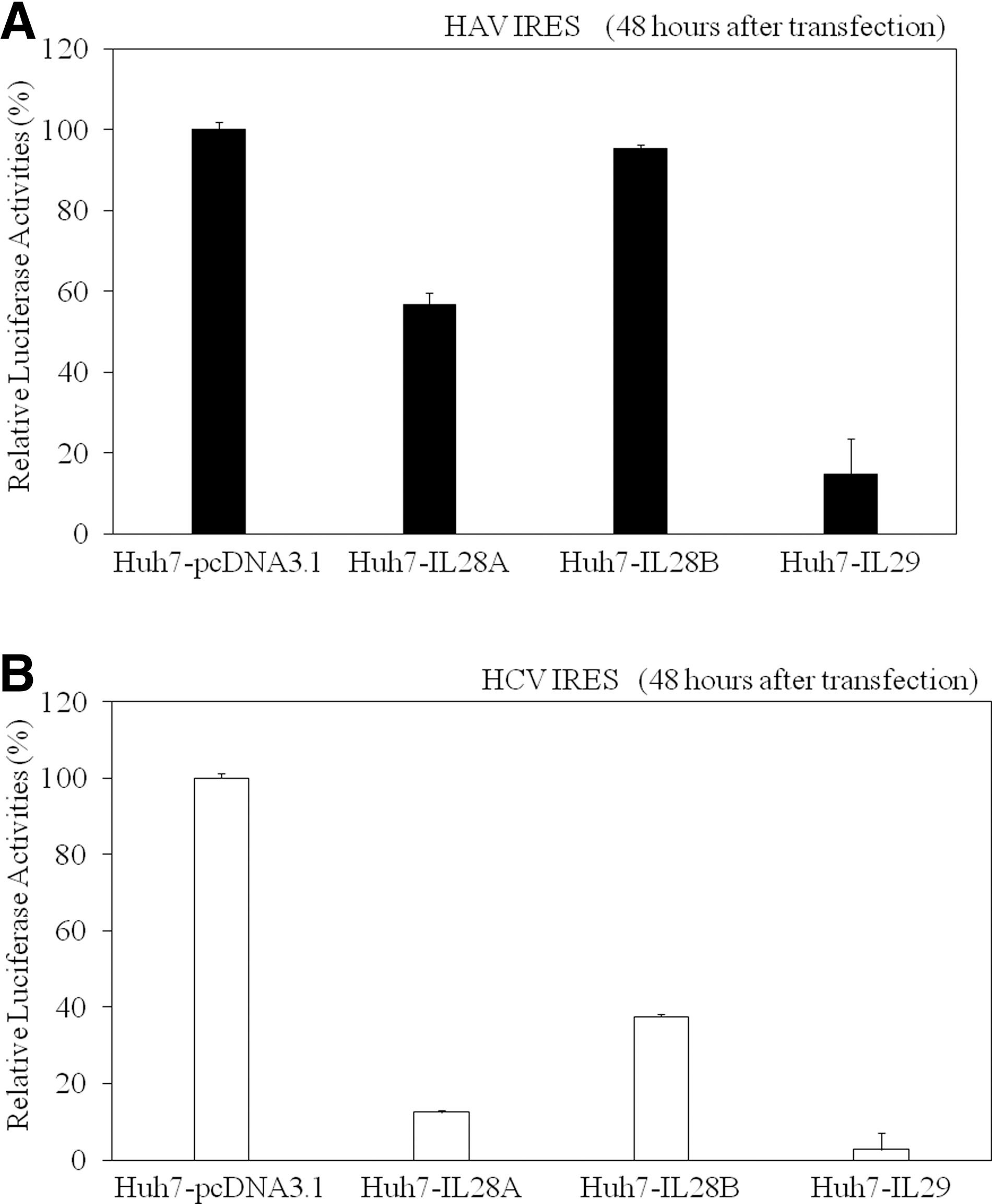
Interferon-λ inhibits hepatitis A virus (HAV) (
HCV IRES activities in Huh7-IL28A, Huh7-IL28B, and Huh7-IL29
It is known that HCV also has IRES structures and plays an important role in the translation of HCV proteins (18,24). In order to compare the effects of IFNs-λ on HCV IRES-mediated translation with those on HCV, we next tested their effects on HCV IRES-mediated translation using a luciferase reporter assay. Huh7-IL28A, Huh7-IL28B, Huh7-IL29, and Huh7-pcDNA3.1 were transfected with pSV40-HCV IRES reporter vector encoding SV40 promoter driven-Rluc and Fluc, separated by HCV IRES (24), and 48 h later, luciferase activity was measured and IRES activity was determined (Fig. 3B). HCV IRES activity was inhibited in Huh7-IL28A (12.5%, n=3, p<0.0001), Huh7-IL28B (37.5%, n=3, p<0.0001), and Huh7-IL29 (2.7%, n=3, p<0.0001), compared to that in control Huh7-pcDNA3.1 (n=3100%). Similarly to HAV IRES, IL-28A and IL-28B demonstrated inhibitory effects on HCV IRES activity, but they seemed less efficient than IL-29 (Fig. 3B).
IL-29 inhibits both HAV and HCV replication
Next we investigated the effect of IL-29 on HAV subgenomic replication in HuhT7 cells (9). IL-29 at 100 ng/mL led to 22.8% (n=3, p=0.038) inhibition of HAV replication, but we observed no reduction of HAV mut replicon replication (Fig. 4A). We also examined whether IL-29 inhibits HAV strain KRM003 propagation in GL37 cells, but at 50 ng/mL of IL-29 we could not observe any effect on the inhibition of HAV propagation. Two-hundred and fifty and 500 ng/mL of IL-29 showed a tendency to inhibit HAV propagation without cell damage. However, it was difficult to obtain a stable reaction. Further study will be needed.

IL29 suppresses hepatitis A virus (HAV) (
We also examined whether IL-29 inhibits HCV replication in Huh7-derived cell lines. Huh7-IL29 and Huh7-pcDNA3.1 were infected with HCV JFH1 (genotype 2a) (14,43), and 72 h later, HCV RNA was detected less in Huh7-IL29 (0.6%; n=3, p<0.0001) than in Huh7-pcDNA3.1 (100%; n=3, p<0.0001; Fig. 4B). HCV core protein expression was also less observed in Huh-IL29 than in Huh7-pcDNA3.1 (Fig. 4C).
Exogenous IL-29 with or without IFN-α or amantadine inhibits HAV IRES activity in Huh7
As Huh7-IL29 cells had the strongest inhibitory effect on HAV IRES-mediated translation (Fig. 3A), we investigated whether exogenous IL-29 had similar effects on HAV IRES-mediated translation using a luciferase reporter assay (Fig. 5A). Huh7 cells were transfected with pSV40-HAV IRES reporter vector (19), and 48 h later, cells were treated with IL-29 with or without amantadine or IFN-α, and 24 h after this, luciferase activity was measured and IRES activity was determined (Fig. 5A).
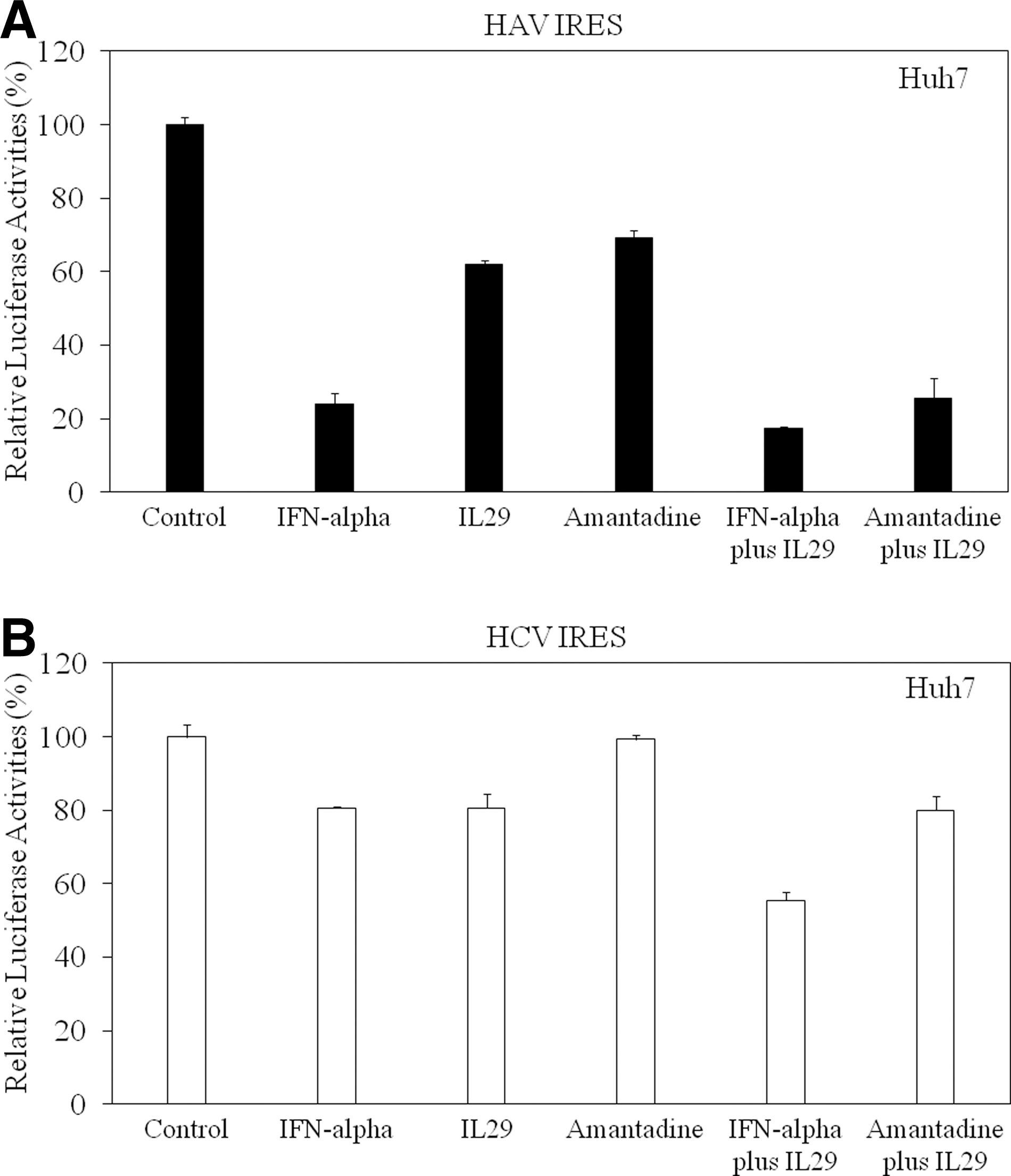
Effects of IL-29 with or without interferon (IFN)-α or amantadine on hepatitis A virus (HAV) (
We previously reported that amantadine with or without IFN-α inhibits HAV IRES-mediated translation in human hepatoma cells (19,45). HAV IRES activity was significantly inhibited, to 24.1% (n=3, p<0.0001), 62.1% (n=3, p<0.0001), and 69.1% (n=3, p<0.0001), by 100 IU/mL IFN-α, 50 ng/mL IL-29, and 50 μg/mL amantadine, respectively (Fig. 5A). The combination of IL-29 with IFN-α or amantadine led to 82.7% (n=3, p<0.0001), or 74.6% (n=3, p<0.0001) inhibition of HAV IRES activity, respectively, with these combinations demonstrating stronger effects than IL-29 alone (Fig. 5A).
In order to compare the effects of exogenous IL-29 on HAV IRES-mediated translation with those on HCV, we next tested the effects of IL-29 with or without amantadine or IFN-α on HCV IRES-mediated translation using a luciferase reporter assay. Huh7 cells were transfected with pSV40-HCV IRES reporter vector (24), and 48 h later, the cells were treated with IL-29 with or without amantadine or IFN-α, and 24 h after this, luciferase activity was measured and IRES activity was determined (Fig. 5B). HCV IRES activity demonstrated significant inhibition, to 80.6% (n=3, p=0.00061) and 80.6% (n=3, p=0.00027) by IFN-α and IL-29, respectively, but showed no inhibition by amantadine only (99.3%, n=3; Fig. 5B). The combination of IL-29 with IFN-α led to a 44.5% (n=3, p<0.0001) inhibition of HCV IRES activity, with this combination demonstrating stronger effects than IL-29 alone. However, the combination of IL-29 with amantadine resulted in only 20% inhibition (n=3, p=0.00027) of HCV IRES activity, similarly to the effect of IL-29 alone (Fig. 5B).
Discussion
We demonstrated that IL-29 inhibited HAV as well as HCV IRES activity in human hepatoma cell line Huh7, and that Huh7-IL29 had stronger effects than Huh7-IL28A, Huh7-IL28B, and Huh7-pcDNA3.1. The combination of IL-29 with IFN-α or amantadine seemed to have stronger inhibitory effects on HAV IRES activity than IL-29 alone.
IFNs-λ modulate innate and adaptive immune responses to environmental pathogens and protect the host against diseases such as cancer. Expression of IFNs-λ is tightly regulated by viral infection, including hepatitis viral infection (6,7). IFNs-λ utilize a receptor complex different from IFN-α, but both types of IFN induce STAT1 and STAT2, as well as STAT3 activation (33,48). Binding of IFN to the IFN receptor leads to the activation of receptor-associated Janus tyrosine protein kinase (Jak1). IFN stimulation results in tyrosine phosphorylation, dimerization, and nuclear import of STATs (3). STATs and the IFN-stimulated gene factor 3 (ISGF3) transcription factor complex move into the nucleus and bind to IFN-stimulated response elements (ISRE) in the promoters of the IFN-stimulated genes (ISGs). ISGs inhibit viral replication and activate numerous downstream cellular responses.
In contrast to IFN-α, IFNs-λ bind to a heterodimeric receptor consisting of IL-28Rα subunit and IL10Rβ receptor subunit, that is also shared by IL-10, IL-22, and IL-26. Because the IFN-λ receptor is different from that of IFN-α, their uses as alternative therapies for viral hepatitis need to be examined (6). Although the biological activities of IFNs-λ could overlap with IFN-α, the expression of IL-28Rα receptor is limited in contrast to the ubiquitously expressed IL10Rβ, and IFNs-λ might have fewer adverse events than type I IFN (28). We also found that the combination of IL-29 with IFN-α or amantadine demonstrated stronger inhibitory effects on HAV IRES activity. The combination of IL-29 with amantadine may also be useful in some HAV patients.
We did not observe any differences in HAV 5′ NTR or HCV 5′ NTR RNA detection by RT-PCR among Huh7-pcDNA3.1, Huh7-IL28A, Huh7-IL28B, and Huh7-IL29 at 72 h after transfection of pSV40-HAV IRES-luc or pSV40-HCV IRES-luc (data not shown), although we could not completely exclude the destruction of IRES mRNAs, because IFNs-λ as well as IFN-α activate double-stranded protein kinase PKR and 2′,5′-oligo A (2–5A) synthetases (21). Several noncanonical translation initiation factors such as La protein and polypyrimidine tract binding protein (PTB) have been implicated in translation from HAV and HCV IRESes (4,13,40,46). The effects of IL-29 on these proteins should be examined in future studies.
We previously demonstrated that siRNAs targeted against HAV IRES, amantadine, and IFN-α, inhibited HAV IRES-mediated translation and HAV replication (15,19,33,45). In the present study, we planned to examine the effects of IFN-λ on HAV IRES-mediated translation. IFNs are proteins induced by lymphocytes and other cells including hepatocytes in response to viruses such as HAV. Our study also supports the notion that IFNs-λ might inhibit HAV IRES-mediated translation as one of the host defense mechanisms against HAV infection.
It has been reported that genetic variations in IL-28B SNPs predict hepatitis C treatment-induced viral clearance and natural clearance (10,16,33,42). Tanaka et al. (42) reported that IL-28B minor SNP was associated with a null virological response in the treatment of Japanese patients infected with HCV genotype 1. Yu et al. (47) also reported that the IL-28B rs8099917 TT genotype is significantly independently predictive of RVR, which is the single best predictor of SVR, in Asian HCV genotype 2 patients. Pegylated IL-29 induces antiviral gene expression and represses hepatitis B and C replication in vitro (6), and HCV replication in vivo (35). Among IFNs-λ, it was reported that IL-28A inhibits HCV IRES-mediated translation and suppresses HCV replication (49). Kato et al. (21) reported that IFN-α, as well as IFN-β, specifically suppress the translation from HCV IRES. We also demonstrated that siRNAs targeted against HCV IRES were potent inhibitors of HCV IRES-mediated translation and HCV replication (18). In the present study, we demonstrated that IL-29, as well as IFN-α, inhibited HCV IRES-mediated translation, although amantadine did not inhibit HCV IRES-mediated translation in our experimental condition.
In conclusion, we demonstrated that IL-29 suppressed HAV as well as HCV IRES-mediated translation. Viral IRES activity may influence the level of replication (8,15,18), although it was reported that the preponderance of host factors might determine the clinical presentation (30). To inhibit HAV or HCV IRES-mediated translation, the combination IL-29 with IFN-α or amantadine has a stronger inhibitory effect. IFNs-λ might also play an important role in host defense mechanisms and in HAV pathogenesis.
Footnotes
Acknowledgments
The authors thank Prof. T. Betakova, Prof. M. Kruger, Prof. S.U. Emerson, Prof. V. Gauss-Muller, Prof. R.B. Ray, and Prof. S.M. Lemon for providing the plasmids and HuhT7 cells. This work was supported by a grant from the Japan Society of Hepatology (T.K.), and a grant from the Chiba University Young Research-Oriented Faculty Member Development Program in Bioscience Areas (T.K.), and a grant from the Ministry of Health, Labour and Welfare of Japan (O.Y.).
Author Disclosure Statement
No competing financial interests exist.