
Abstract
Mosquito salivary proteins inoculated during blood feeding modulate the host immune response, which can contribute to the pathogenesis of viruses transmitted by mosquito bites. Previous studies with mosquito bite-naïve mice indicated that exposure to arthropod salivary proteins resulted in a shift toward a Th2-type immune response in flavivirus-susceptible mice but not flavivirus-resistant animals. In the study presented here, we tested the hypothesis that immunization with high doses of Culex tarsalis salivary gland extracts (SGE) with an adjuvant would prevent Th2 polarization after mosquito bite and enhance resistance to mosquito-transmitted West Nile virus (WNV). Our results indicate that mice immunized with Cx. tarsalis SGE produced increased levels of Th1-type cytokines (IFNγ and TNFα) after challenge with mosquito-transmitted WNV and exhibited both a delay in infection of the central nervous system (CNS) and significantly lower WNV brain titers compared to mock-immunized mice. Moreover, mortality was significantly reduced in the SGE-immunized mice, as none of these mice died, compared to mortality of 37.5% of mock-vaccinated mice by 8 days after infected mosquito bite. These results suggest that development of a mosquito salivary protein vaccine might be a strategy to control arthropod-borne viral pathogens such as WNV.
Introduction
West Nile virus (WNV), a single-stranded positive-sense RNA virus, belongs to the family Flaviviridae, genus Flavivirus, and is a member of the Japanese encephalitis virus serological complex, which includes Japanese encephalitis, Saint Louis encephalitis, and Murray Valley encephalitis viruses (11). WNV is naturally transmitted in an enzootic cycle involving Culex spp. mosquitoes and birds. Human infections occur when ornithophilic mosquitoes, including Culex tarsalis, which is a major vector in western North America (31,33), shift feeding preferences to mammals during periods of avian host dispersal and migration (14). It has been shown that mosquito salivary proteins enhance WNV infections during mosquito feeding in mammals and birds (27,28) and this enhancement in mammals can be attributed to a shift of the host immune response from a protective Th1 response to a less favorable Th2 response, altering cytokine production and affecting antiviral properties of immune system cells (25,42). Although earlier studies suggested that previous exposure of the host to vector saliva could confer protection to parasitic infections (6,13,29), this has not been demonstrated for arbovirus infections. To the contrary, Styer et al. (28) showed that mice with antibodies to Culex spp. mosquito salivary proteins resulting from previous mosquito bites exhibited similar enhancement of mosquito-transmitted WNV infection as nonsensitized mice, and Schneider et al. (23) found that previous sensitization to salivary proteins delivered by multiple A. aegypti bites resulted in intensified subsequent mosquito-transmitted WNV disease due to enhanced early viral replication.
Effective vaccines to protect against leishmaniasis have been developed from specific proteins from sand fly vector saliva (10,34), and a tick salivary protein vaccine disrupted feeding and prevented transmission by ticks infected with tick-borne encephalitis virus (15); but to our knowledge, no mosquito salivary protein vaccine has been tested for protective efficacy against arbovirus infections. In this study, we tested the hypothesis that immunization of mice with high doses of proteins from Cx. tarsalis salivary gland homogenate in the presence of an adjuvant would elicit a robust immune response and potential alteration of Th1–Th2 polarization and alleviate pathogenesis of subsequent WNV infection transmitted by the natural vector.
Materials and Methods
Mosquito rearing and WNV infection
Cx. tarsalis originally captured in Bakersfield, California, were obtained from the Centers for Disease Control and Prevention (CDC), Fort Collins, Colorado. Mosquitoes were reared at 24°–25°C with 70% relative humidity in a 12h/12h cycle of light/darkness at the Arthropod-Borne and Infectious Diseases Laboratory (AIDL). Adult female mosquitoes were offered water and sugar ad libitum for 4 days after emergence. At day 5, sugar and water were removed for 24 h, after which mosquitoes were fed an infectious blood meal of de-fibrinated sheep blood containing 7.3 log10 PFU/ml WNV (strain NY99) using a water-jacketed glass feeder. To attain the number of infected mosquitoes needed for challenge experiments, separate cohorts were infected by intrathoracic (IT) inoculation of 1 μL containing 4.3 log10 PFU WNV. After an infectious blood meal or IT inoculation, 50 mosquitoes were placed in separate plastic cages and supplied with water and sugar ad libitum for 7 days until challenge studies. Mosquitoes were deprived of water and sugar for 24 hours before feeding on mice.
Salivary gland homogenate protein preparation
Adult female Cx tarsalis were cold-anesthetized and surface sterilized by submerging in 70% ethanol. Salivary glands were dissected and placed into ice cold lysis buffer (10 mM Tris-HCl [pH 7.5], 150 mM NaCl, 5 mM EDTA, 1% sodium deoxycholate, 1% Triton-X-100, 0.1% SDS, 1 mM phenylmethylsulfonyl fluoride). One hundred salivary glands were pooled and mechanically disrupted with a pestle in a 1.5 mL microcentrifuge tube and the homogenate was centrifuged at 3500 rpm for 5 min at 4°C. Cell-free supernatant was collected, and proteins were concentrated by 10% trichloroacetic acid precipitation. Proteins were resuspended in 100 μL of sterile phosphate-buffered saline (PBS, pH 7.4) containing a protease inhibitor cocktail (Roche Laboratories). Protein concentrations were determined with a BCA Protein Assay Kit (Pierce) according to the manufacturer's instructions, and salivary gland extracts (SGEs) were diluted to a protein concentration of 10 ng/μl and mixed with an equal volume of 10 ng/μL synthetic peptide adjuvant, N-acetylmuramyl-L-alanyl-D-isoglutamine (MDP; Sigma), for vaccine preparation.
Mouse immunization protocol
All vertebrate animal experiments were performed in accordance with an approved Institutional Animal Care and Use Committee protocol. Two week-old NIH Swiss mice were purchased from Charles River Laboratories and housed in a BSL-3 facility at Colorado State University. Ten mice per group (in two independent experiments) were immunized intramuscularly with 500 ng Cx. tarsalis SGPs mixed with 500 ng MDP (100 μL total volume). Two booster immunizations were given intramuscularly at days 7 and 21 after the first immunization. The control group was mock-immunized with PBS mixed with 500 ng of MDP and given booster mock-immunizations at days 7 and 21. Vaccinated and mock-vaccinated mice were bled from the tail vein 1 week after the last booster immunization for antibody analysis.
Immunoblot analysis of anti-salivary gland protein antibodies
Confirmation of antibody generation against mosquito salivary gland proteins in vaccinated mice was accomplished by immunoblot assay. Homogenate proteins were prepared from ten female Cx. tarsalis salivary glands as described above and re-suspended in 30 μL PBS containing a protease inhibitor cocktail mixed with 7.5 μL of 4X lithium dodecyl sulfate buffer. Twelve microliters of this solution were loaded in each lane of a SDS 10% polyacrylamide gel, fractionated at 200 volts for 40 min, and proteins were transferred to a nitrocellulose membrane. Because of limited blood volumes obtained by tail-vein bleeding, serum samples from each experimental group were pooled in 100 μL minimum essential medium (MEM), centrifuged at 3500 rpm for 10 min, and the supernatant collected. One hundred microliters of pooled serum mixture were diluted in 1000 μL blocking buffer and incubated overnight on nitrocellulose strips containing fractionated proteins. The serum mixture was removed and strips were washed 3X with 0.01% Tween 20 in PBS and incubated with 1:1000 peroxidase-labeled secondary rabbit anti-mouse IgG (KPL, Kirkegaard & Perry Laboratories, Inc.) in blocking buffer at room temperature for 1 h, washed 3X again, and developed with 3,3’- diaminobenzidine (Sigma). The reaction was stopped with deionized water and immunoblots were scored by direct visualization of bands.
Mouse infection by mosquito bite
Ten days after the last booster immunization, mice were anesthetized and placed into cages containing WNV-infected mosquitoes. Three to five mosquitoes were allowed to take a blood meal on each anesthetized mouse, and engorged mosquitoes were quickly aspirated from the cages and immediately frozen at −80°C until assayed for WNV infection. Mice were observed daily and euthanized when they exhibited severe clinical signs (i.e., hunched posture, ruffled fur, and neurologic signs such as hind leg paralysis or severe hind limb weakness).
Tissue harvesting and WNV RNA extraction
At 4 and 8 days post-infection, mice were humanely euthanized and blood, brains, and spleens were collected for analysis. Serum was separated, diluted 1:2 in MEM and frozen at −80°C. Portions of brain and spleen (50 μg) were placed in 300 μL MEM containing 2% fetal bovine serum (FBS) plus antibiotics (Gibco) and tissues were homogenized, centrifuged, and supernatants frozen at −80°C for titration by plaque assay. RNA was extracted from tissue pellets using Trizol (Invitrogen) according to the manufacturer's recommendations.
RNA detection and measurement
Superscript II RT (Invitrogen) was used to generate cDNA from WNV RNA in mosquito and mouse tissues and cytokine mRNAs in mouse tissues. Manufacturer's recommendations were followed using 100 ng of total RNA as template with 1 pmol of either WNV or cytokine-specific primers and 10 mM dNTPs in a total volume of 12 μL. Five microliters of cDNA were used as template for PCR or qPCR with SYBR green (Stratagene) master mix and Platinum Taq polymerase (Invitrogen) with 1 pmol of forward and reverse primers, following the manufacturer's instructions. The cycling program for WNV cDNA was 95°C for 5 min, 35 cycles of 95°C for 1 min, 50°C for 1 min, and 72°C for 2 min, with a final extension at 72°C for 8 min. Resultant DNA was evaluated by electrophoresis in a 1% agarose gel. Primers used for WNV RT-PCR (16) are listed in Table 1.
Forward primer (+), reverse primer (-), β-actin, beta-actin; IFNγ, interferon-gamma; IL-4, interleukin-4; IL-10, interleukin-10; TNFα, tumor necrosis factor-alpha; WNV, West Nile virus.
Murine Th1-associated (IFNγ and TNFα) and Th2-associated (IL-4 and IL-10) cytokine mRNA were quantitated by RT-qPCR as previously described (41). Reactions were standardized according to Overbergh et al. (21), and primers amplified regions that spanned intron-exon junctions to avoid amplification of contaminating DNA. Sequences for primers are given in Table 1 (9,21). Controls included amplification without adding RNA and qPCR without previous reverse transcription. The cycling program for qPCR of cytokine cDNA was as follows: 95°C for 10 min, 40 cycles of 95°C for 30 sec, and 60°C for 1 min. For normalization, murine β-actin cDNA was amplified from each sample in parallel. The ratio of test-gene cDNA to β-actin cDNA represented the relative level of cytokine mRNA in each sample. Cytokine quantitation data were analyzed using the iQ5 system software (BioRad).
Infectious virus titration by plaque assay
WNV titers in mosquitoes and mouse tissues were determined by plaque assays as described previously (41). Briefly, Vero cells were grown at 37°C in 12-well cell culture plates in MEM with 10% FBS for 24 h before infection. Medium was removed and serial 10-fold dilutions of supernatant from tissue homogenates in MEM were added to triplicate plates and incubated for 2 h. Subsequently, MEM containing 2% FBS and 1% low-melting-point agarose was added and the plates were incubated for 3 days. A second overlay of 1% agarose medium containing 0.01% neutral red vital stain was added for 24 h to visualize plaques, which were counted and averaged for each dilution. Virus concentrations were calculated as PFU per ml of supernatant.
Plaque reduction neutralization test (PRNT80)
WNV antibody titers in mouse serum were determined by 80% plaque reduction neutralization test (PRNT80) using a previously published protocol (26) with the following modifications: mouse sera were diluted twofold from 1:2 to 1:128 in MEM containing heat inactivated 2% FBS, and incubated with 100 PFU WNV(NY 99) in a total volume of 0.1 ml overnight at 4°C. WNV-serum complexes were added to Vero cell monolayers and incubated as described above for plaque assays. Plaques were counted and the reciprocal of the highest serum dilution that reduced plaque numbers by 80% was reported as the end-point titer.
Statistical analysis
RT-qPCR results were analyzed by nonpaired Student's t-tests for a single mean comparison. PRNT results were analyzed assuming a Poisson distribution and using logarithm transformation. Results are presented by experimental group and time after challenge with 95% confidence intervals. Survival curves for both groups of animals were generated and differences in surviving numbers were analyzed by Fisher's exact test as described in a previous study (23).
Results
Quantitation of viral RNA and infectious virus in tissues of vaccinated mice
After immunized or mock-immunized mice were fed upon by WNV-infected mosquitoes, their brains and spleens were collected at days 4 and 8 post-challenge to analyze WNV RNA and infectious virus titers. At day 4 post-mosquito challenge, viral RNA was detected by RT-PCR in the brains of 3 of 8 mock-vaccinated mice (37.5%) compared to 0 of 8 vaccinated mice (0%). WNV RNA was detected in 3 of 8 spleens of immunized mice and 1 of 8 mock-vaccinated mice (Table 2). Although there was a delay in WNV CNS infection in vaccinated mice, by day 8 the brains of all mosquito-challenged mice were positive for WNV RNA by RT-PCR. Likewise, infectious virus was detected in brains of mock-immunized mice at 4 days post-challenge but was below the level of detection by plaque assay in immunized mice (Table 2). Infectious virus was found in the brains of all challenged mice by 8 days post-challenge. An exact test of proportions showed a significant difference (p<0.009) in brain virus titers between vaccinated (2.8 log10 PFU/mL) and mock-vaccinated (5 log10 PFU/mL) mice 8 days after mosquito challenge. Interestingly, vaccinated mice had significantly higher infectious virus titers in spleens at day 4 post mosquito challenge (3 log10 PFU/mL) compared to mock-immunized mice (0.5 log10 PFU/mL). However, no infectious virus was detectable in spleens of either group by day 8 after mosquito challenge (Table 2).
WNV RNA was detected by RT-PCR and infectious WNV was quantitated by plaque assay in brains and spleens of mice immunized with mosquito salivary gland extract (SGE) or mock immunized at 4 and 8 days post WNV challenge (dpi) by infected Cx. tarsalis bite.
Between days 5 and 7 post infection, 7 of 8 mock-vaccinated mice demonstrated clinical signs of WNV infection (extreme weakness, lack of mobility in the hind limbs), and 3 of those 7 were euthanized according to animal use protocols. By comparison, between days 7 and 8 post-challenge, all 8 immunized mice showed only mild clinical signs of infection (reduced activity and increased huddling during daylight hours) but did not require euthanasia (Fig. 1). When all data from two independent experiments for both time points for vaccinated and mock-vaccinated groups were compared, post-challenge virus titers were significantly higher (p<0.0025) in all tissues of mock-vaccinated mice. Likewise mortality rates (due to euthanasia) were significantly higher in the mock-immunized group (Fisher's exact test; p=0.03), as no fatalities occurred and euthanasia was not required for the SGE-immunized group by day 8 post-mosquito challenge.
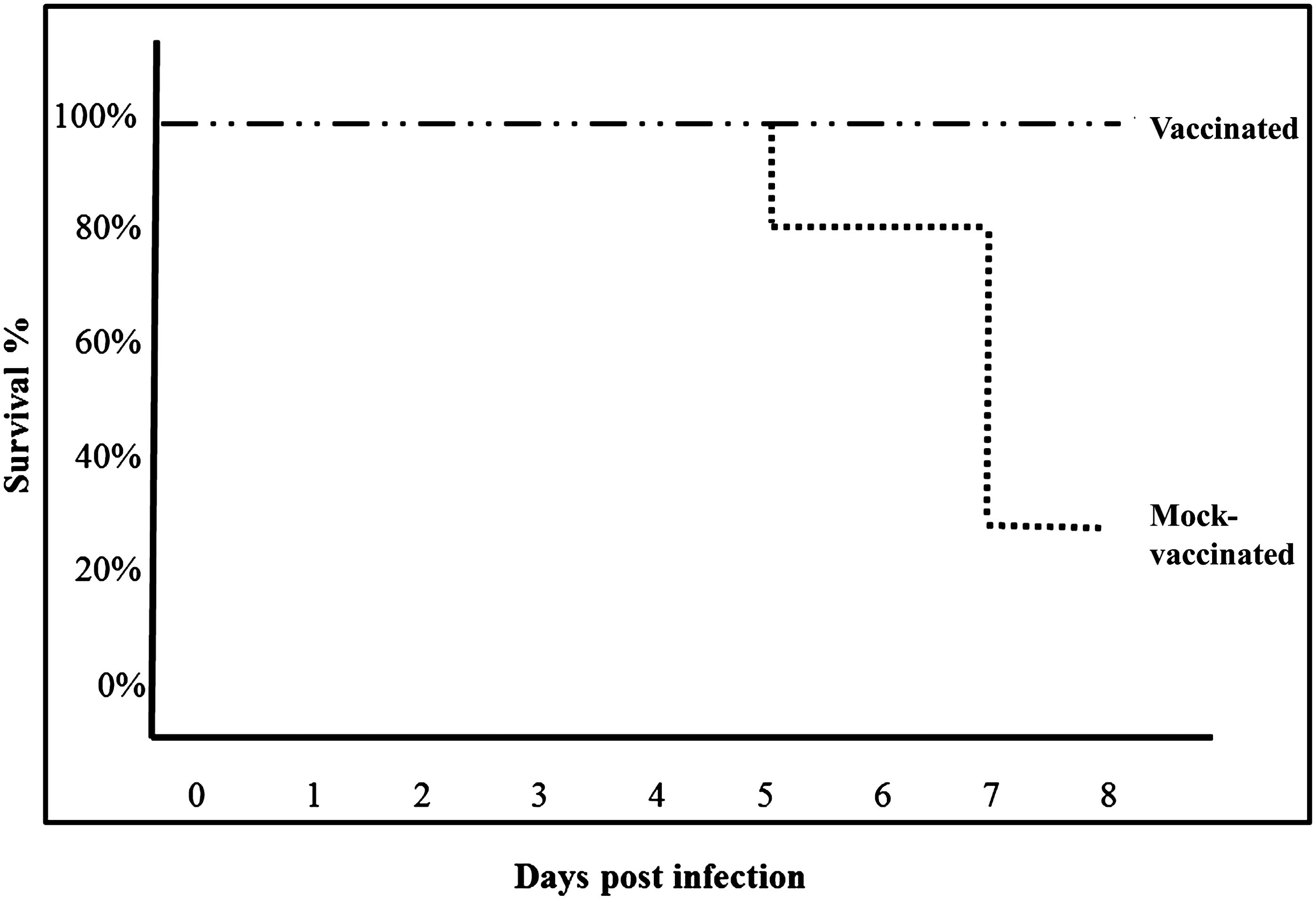
Survival of mice immunized with Cx. tarsalis salivary gland homogenates or mock-immunized following WNV challenge by bites of infected Cx. tarsalis. Groups of 8 mice were either vaccinated with mosquito salivary gland extract or mock-immunized and challenged by bites of 3–5 WNV-infected Cx. tarsalis mosquitoes. Mice were euthanized immediately when they exhibited severe neurologic signs of disease. Data from two independent replicates were analyzed; survival was significantly higher in immunized mice (Fisher's exact test; p=0.03).
Antibody and cytokine responses of mice vaccinated with Cx. tarsalis salivary gland extracts
Mice were immunized or mock-immunized as described in Materials and Methods, and 7 days after the last booster immunization sera were collected for immunoblot analysis. Proteins concentrated from Cx. tarsalis salivary gland homogenate were fractionated by PAGE, transferred to a nitrocellulose membrane, and incubated with pooled sera of vaccinated or mock-vaccinated mice to confirm humoral immunity. Reactivity to SGE antigens of immunized mouse sera compared to controls (Fig. 2) demonstrated that immunization elicited antibodies against a number of salivary gland proteins. Prominent antibody reactivity was directed to SGE proteins with approximate masses of 17 kDa, 30 kDa, 36 kDa, 47 kDa, 49 kDa, 68 kDa, and 85 kDa.
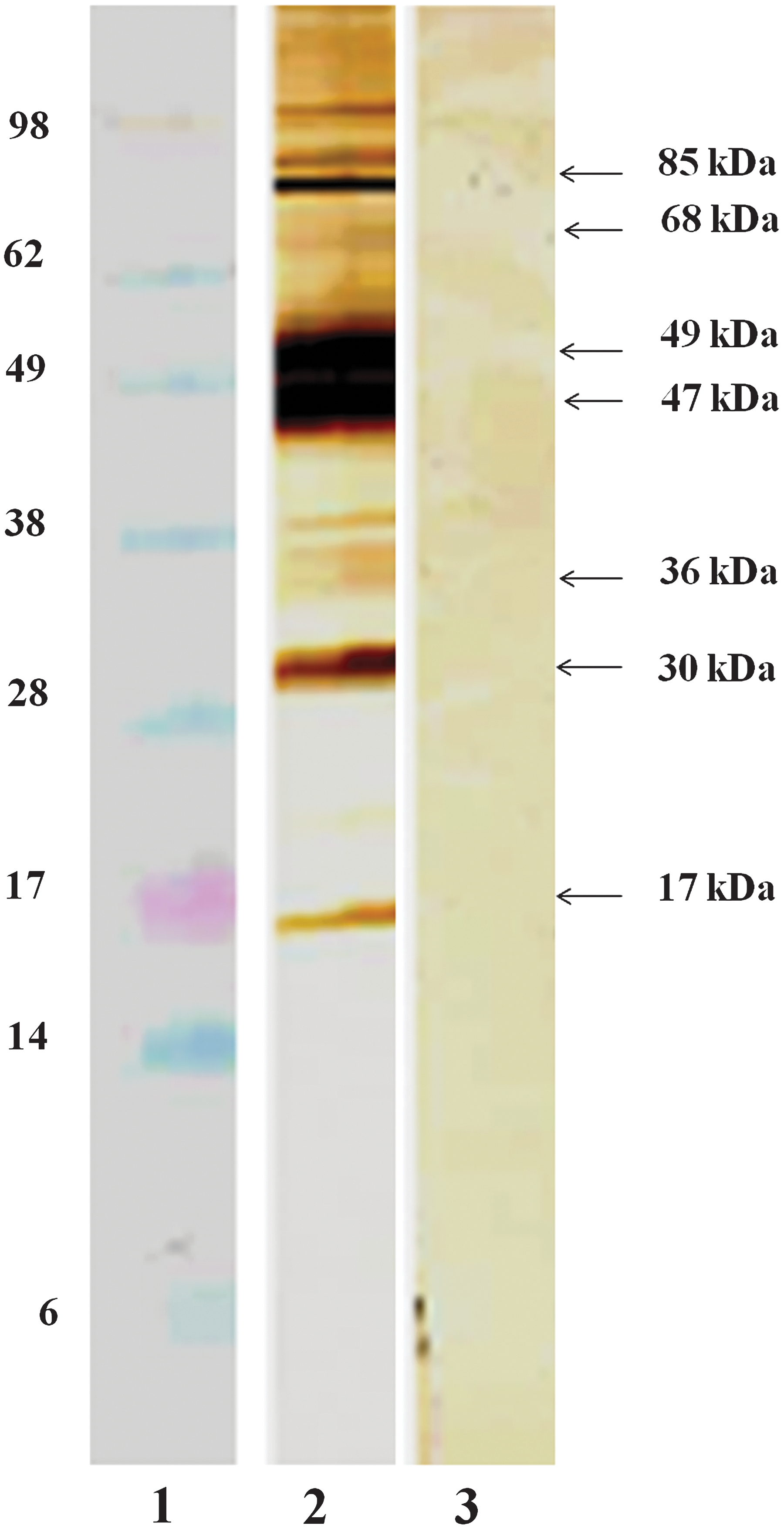
Immunoblot analysis of pooled serum from mice immunized with Cx. tarsalis salivary gland homogenates. Lane 1, molecular mass markers, Lane 2, pooled serum from vaccinated mice, and Lane 3, pooled serum from mock-vaccinated mice. Approximate molecular masses of mosquito salivary proteins recognized by mouse antibodies are indicated by arrows at right.
WNV-neutralizing antibody titers in mouse sera following WNV infection by mosquito-bite from two independent experiments were assayed by PRNT80 and averages are shown in Figure 3. At 4 days after mosquito feeding, mice vaccinated with Cx tarsalis salivary gland extract had increased neutralizing antibody titers compared to mock-immunized mice. By day 8, a sharp rise in anti-WNV antibody titers was noted in both immunized and control mice (Fig. 3); however, immunized mice had significantly higher neutralizing antibody titers compared to mock-immunized mice (p<0.0008). This correlated with a significantly reduced virus load within the CNS of immunized mice, as shown in Table 2.
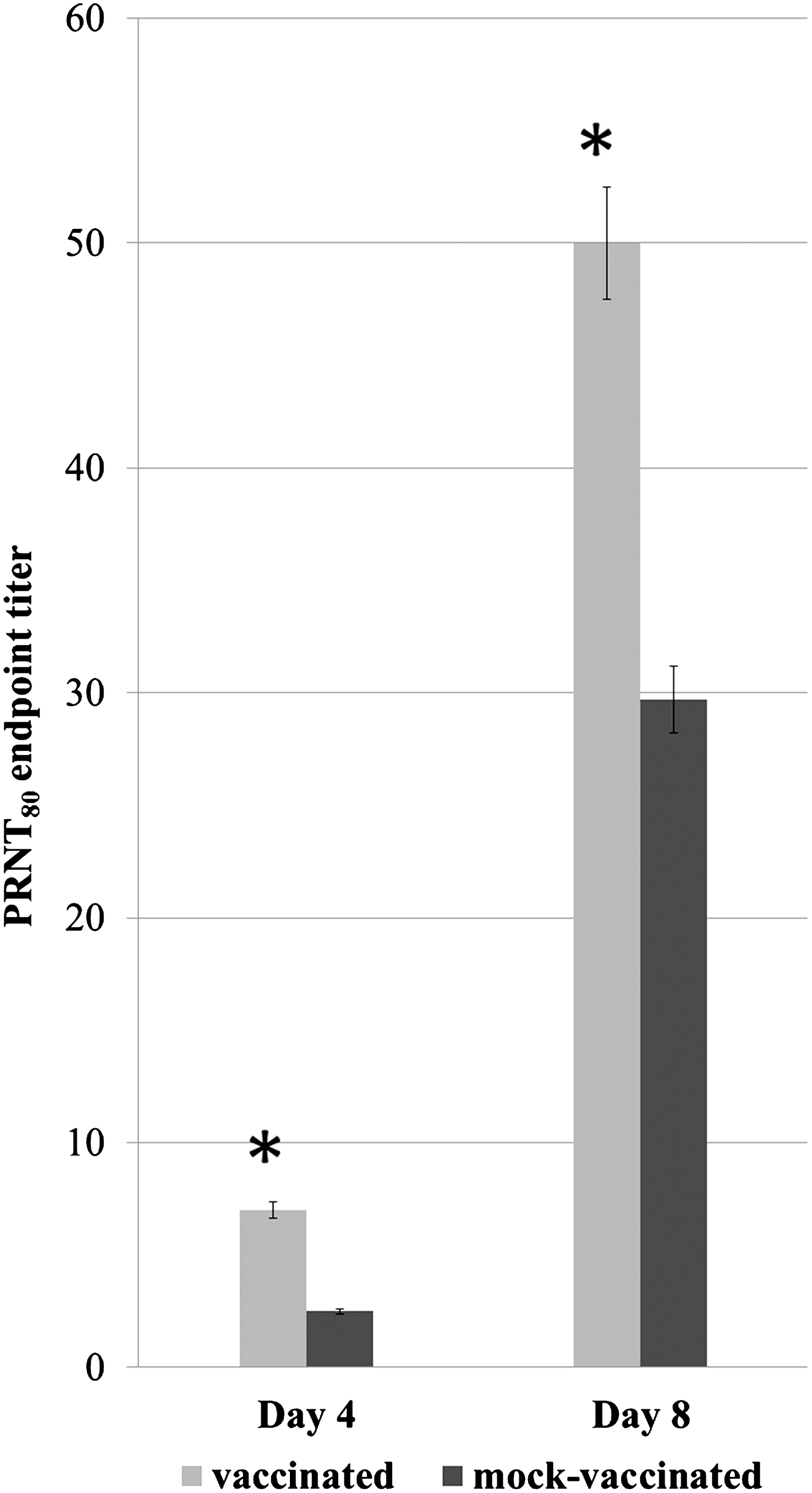
WNV neutralizing antibody titers (PRNT80) in vaccinated and mock-vaccinated mice at days 4 and 8 post WNV challenge. Sera from vaccinated and mock-vaccinated mice were assayed for WNV neutralizing antibodies by 80% plaque-reduction neutralization test (PRNT). Error bars represent the standard error of the mean of two independent experiments, N=8 mice per group. *indicates a significant difference between groups (p<0.0008).
Quantitation of mRNA for Th1- or Th2-associated cytokines in the spleens of vaccinated and mock-vaccinated mice by RT-qPCR indicated that expression of Th1-associated cytokines IFNγ and TNFα was significantly higher in immunized mice (p<0.001) by day 4 after virus challenge (Fig. 4A and B). IFNγ remained significantly higher in spleens of immunized mice at 8 days post challenge (Fig. 4A). The Th2 cytokine IL-4 mRNA levels were slightly higher at 4 days and significantly higher at 8 days post challenge in immunized mice (Fig. 4C), whereas IL-10 mRNA levels were significantly higher in immunized mice at day 4 and significantly decreased relative to mock-immunized mice by day 8 post mosquito challenge (Fig. 4D).
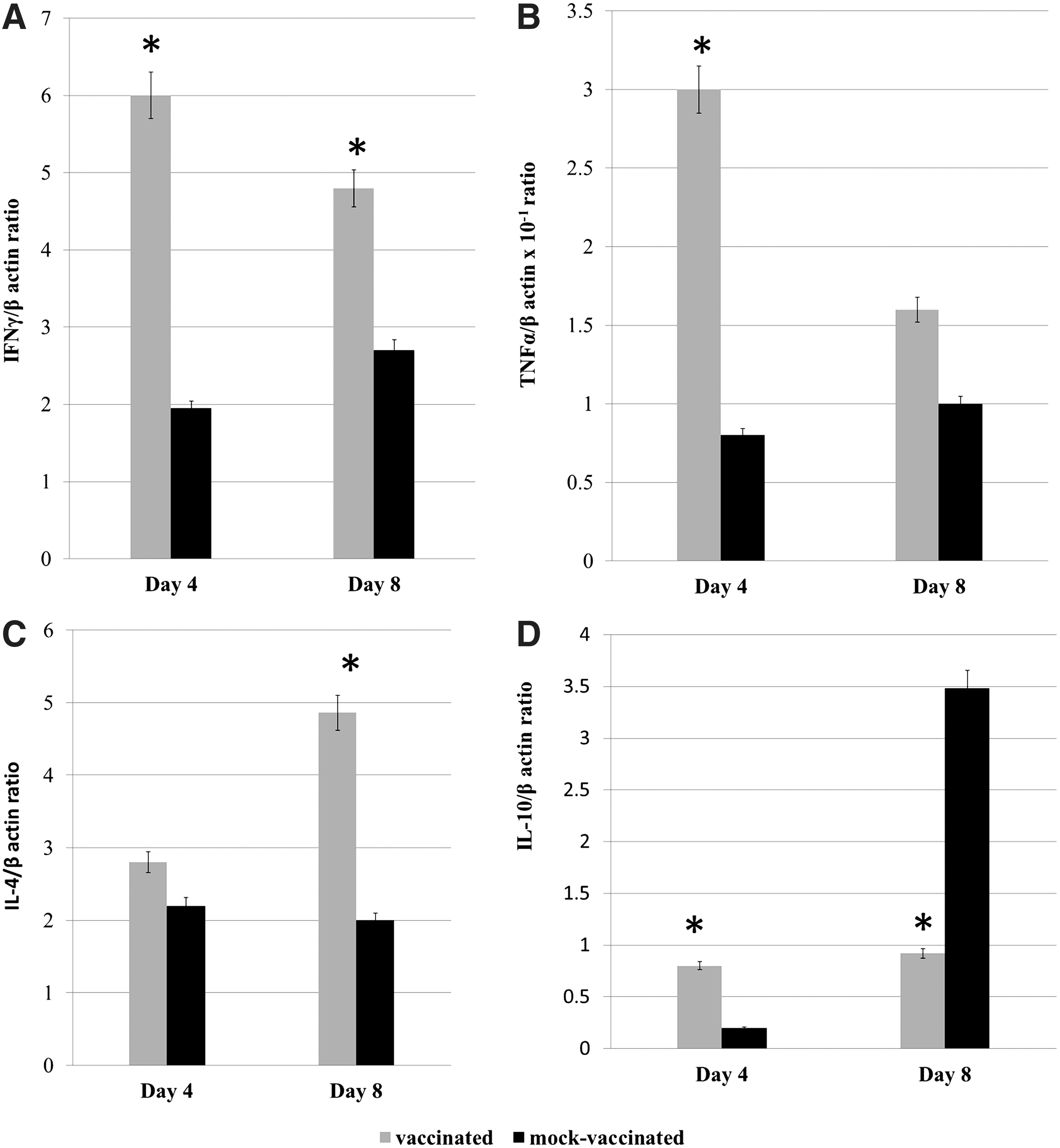
Splenic cytokine mRNA production in vaccinated and mock-vaccinated mice determined by RT-qPCR at days 4 and 8 post-WNV challenge; The Y-axis depicts the ratio of amplified cytokine cDNA to the β-actin gene of each sample. A, IFNγ; B, TNFα; C, IL-4; and D, IL-10. The bars indicate the standard error of the mean and the data are representative of 2 independent experiments, N=8 mice per group and time point. * indicates a significant difference (p<0.05) between groups.
Discussion
Saliva of several different vectors has been shown to enhance infection by arthropod-transmitted viruses in naïve animal models (7,17,27,28). In general, arthropod saliva contains proteins capable of regulating inflammatory mediators and modulating T cell-derived cytokines (8). Inflammation inhibitors and certain modulators of cytokine expression are known to prevent vertebrate hosts from triggering an effective immune response. This was demonstrated when mice were either injected with a synthetic mosquito salivary peptide or fed upon by uninfected mosquitoes, each of which significantly affected the Th1/Th2 cytokine balance, downregulating IFNγ production while upregulating IL-4 and IL-10 in flavivirus-susceptible mice (42).
In the current study, we immunized outbred NIH-Swiss mice with Cx. tarsalis SGE proteins in the presence of an adjuvant to determine the effect of immunity to mosquito salivary proteins on WNV pathogenesis after infection by the bite of Cx. tarsalis. Experimental models for other vectors demonstrated that previous exposure to sand fly saliva protected mice against cutaneous leishmaniasis (13,29). Vaccination of hamsters with a single protein from Phlebotomus papatasi (SP15) resulted in generation of specific antibodies against the natural protein and elicitation of a strong delayed-type hypersensitivity reaction, resulting in protection against Leishmania infection. The induction of both humoral and cellular immunity resulted in smaller lesions and significantly lower parasite burden in this model (10). In this study, we found that immunization of mice with Cx. tarsalis SGE modulated WNV infection in mice by limiting virus replication within the CNS, resulting in decreased mortality. Immunized mice exhibited a significant delay in virus dissemination to the brain in addition to significantly lower virus titers on day 8 after infected mosquito challenge, whereas mock-immunized mice challenged by WNV-infected mosquitoes demonstrated higher virus titers in brains at both 4 and 8 days post challenge. Although there was a significant delay in dissemination to the CNS in immunized mice, the brains of all mice eventually became infected; however, lower virus titers within the CNS correlated with a lack of mortality in the SGE-immunized mice by day 8 post mosquito challenge.
Schneider et al. (24) demonstrated potentiation of WNV encephalitis following challenge by infected Aedes aegypti in mice that had been exposed to repeated feeding by uninfected A. aegypti 4–6 weeks prior to infectious bite, and Styer et al. (28) found that enhancement of WNV infection by mosquito saliva still occurred in mice with antibodies to Culex spp. mosquito saliva elicited by previous mosquito feeding. In contrast, we found that mice immunized in the presence of an adjuvant with high doses of SGE proteins from Cx. tarsalis, a natural WNV vector responsible for a high proportion of human and equine infections in western North America (31,32), exhibited decreased signs of disease and mortality. It is known that both Aedes spp. and Culex spp. mosquito feeding can modulate the balance between Th1 and Th2 cytokine responses, greatly increasing production of Th2 related cytokines while inhibiting elaboration of Th1 cytokines (25 42). It is also apparent that the Th1 cytokine response is important in protecting mice during early WNV infection (26,36). We found that SGE-immunized mice showed decreased disease and mortality while maintaining splenic IFNγ and TNFα production on days 4 and 8 after challenge by infected mosquitoes. In mock-vaccinated mice, significant increases in IL-10 production on day 8 post-infection concomitant with decreases in IFNγ production appeared to correlate with higher virus titers in the CNS and resultant mortality. Although IL-4 mRNA levels were significantly higher at 8 days post challenge in immunized mice, immunization with Cx tarsalis SGE appeared to decrease or delay the shift to Th2 cytokine expression induced by mosquito feeding and consequently arm the murine host to both resist the effects of immune modulation by feeding mosquitoes and provide at least some resistance to virus replication, dissemination, and resultant pathogenesis. The upregulation of IL-10 expression in mock-vaccinated mice could have been responsible for lower production of IFNγ due to tightly inter-related control of these two cytokines (12). Interleukin-10 is known as a potent immunosuppressive and anti-inflammatory cytokine that deactivates macrophages and dendritic cells (19). Increased production of IL-10 during WNV infection has proven to be detrimental to a protective antiviral immune response due to deregulation of signaling in antigen-presenting cells and T lymphocytes at the site of infection (2,22), resulting in increased early viral replication and higher acute viremia titers. Thus, lower production of IFNγ as a result of an increase in IL-10 expression leads to an inability to successfully control infectious pathogens (12). Interestingly, in our study, IFNγ production was maintained through days 4 and 8 after infected mosquito challenge in vaccinated compared to mock-vaccinated animals. IFNγ is known to be a critical factor in virus susceptibility and pathogenesis and it has been observed that a lack of IFNγ production results in increased vulnerability to WNV infection of the CNS and greater viral replication in lymphoid tissues (26). IFNγ production has been attributed to a number of cell types, including γδT-cells, the same cell type that facilitates adaptive immunity against WNV infection in mice (37,38). It is possible, although not examined in these studies, that antibodies produced in response to immunization with SGE affected either γδT cell production of IFNγ or neutralized some factor involved in the dysregulation of IFNγ production modulated by vector feeding. Future studies will be needed to answer this question.
Expression of mRNA for TNFα was also measured in these studies and was significantly upregulated in immunized compared to mock-vaccinated mice. TNFα upregulation was observed in vaccinates at 4 days post challenge; this early increase in TNFα could be another important factor in our observation that vaccination against mosquito salivary proteins greatly improves the cellular immune response very early in the course of infection. TNFα was previously reported to be an important cytokine in the control of infections by other flaviviruses such as dengue virus (20), and TNFα production was increased when dengue virus-infected myeloid dendritic cells had been previously exposed to mosquito saliva (1). This finding is similar to our observation that immunization of mice with salivary gland components leads to early production of TNFα in the face of mosquito-transmitted WNV. Although previous reports indicated that TNFα could be in part responsible for early WNV neuroinvasion due to increased permeability of the blood-brain barrier (39), this was not corroborated in this study. Our data align with other reports in which early production of TNFα is a protective factor against WNV neuroinvasion (26). We noted a higher level of IL-10 in immunized mice on day 4 post-infection; nevertheless, the TNFα increase, along with a significant increase in IFNγ correlated with lower viral loads and protection against early viral invasion of the CNS, corroborating previous reports suggesting synergy between IFNγ and TNFα in conferring antiviral immunity (18).
Our study also demonstrated that anti-WNV antibody titers were significantly higher in SGE-vaccinated as compared to mock-vaccinated mice at days 4 and 8 post challenge. Production of antibodies is important in limiting WNV infection (5). Development of WNV-specific neutralizing IgM has been observed beginning on day 4 after subcutaneous needle-delivered infection. Mice lacking IgM antibodies were susceptible to lethal WNV infection and exhibited sustained viremia, as well as earlier WNV entry into the CNS (5). The importance of a sustained antibody response was also demonstrated by passive transfer of serum from wild-type to immune deficient mice, resulting in significant protection from lethal WNV infection. This was confirmed in B cell-deficient mice, in which viral dissemination to the brain could be demonstrated as early as 4 days post infection (5). In the study presented here, significantly higher neutralizing antibody responses in vaccinated mice correlated with delayed dissemination to and lower viral titers in the CNS compared to control mice.
Based on our data, vaccination with high doses of mosquito SGE in the presence of adjuvant appears to limit WNV replication after challenge by infected mosquitoes. Our study indicated elicitation of a more robust immune response during early infection of vaccinated mice. Although late virus dissemination to the brain was not prevented by immunization with Cx tarsalis SGE, no deaths occurred by 8 days after challenge in immunized animals, a significantly different outcome compared to mock-immunized animals. These data provide information that could be used in developing vector-derived immunogens for use as vaccines. Identification of individual Cx. tarsalis salivary antigens and testing their efficacy as vaccines in modulating subsequent WNV infections may provide a rational strategy for induction of potent immune protection to WNV disease.
Footnotes
Acknowledgments
This research project was supported in part by contract N01-AI-25489 from the NIH/National Institute of Allergy and Infectious Diseases.
Author Disclosure Statement
No competing financial interests exist for any of the authors.