
Abstract
Unlike the polymorphic MHC class Ia molecules, MHC class Ib molecules are oligomorphic or nonpolymorphic. We recently discovered a protective CD8 T cell response to mouse polyomavirus (MPyV) in H-2b haplotype mice that is restricted by H2-Q9, a member of the Qa-2 MHC class Ib family. Here, we demonstrate that immunization with a peptide corresponding to a virus capsid-derived peptide presented by Q9 also elicits MHC class Ib-restricted MPyV-specific CD8 T cells in mice of H-2s and H-2g7 strains. These findings support the concept that immunization with a single MHC class Ib-restricted peptide can expand CD8 T cells in MHC class Ia allogeneic hosts.
Polyomaviruses are ubiquitous viruses that persist in a smoldering infectious state in many vertebrate species, including humans (12,32,35). In healthy hosts, polyomavirus infection is clinically silent (6). However, immunosuppression resulting from HIV/AIDS, aging, and immunomodulatory agents may result in unchecked viral replication with life-threatening complications, such as nephropathy in kidney transplant patients, central nervous system demyelination, and cutaneous malignancies (2 –4,26,28,37).
Using MHC class Ia-deficient C57BL/6 mice (B6.Kb-/-Db-/-), we recently discovered a class Ib-restricted CD8 T cell response that confers protection against MPyV infection (31). A peptide derived from amino acids 139–147 of the VP2 capsid protein (VP2.139; HALNVVHDW) binds Q9, a ß2m-associated MHC class I molecule encoded in the mouse Qa-2 locus. H2-Q9 is nonpolymorphic in mice (30). In addition, H2-Q9 shares the peptide binding specificity of the nearly identical Qa-2 family member H2-Q7 (29). In this study, we tested the hypothesis that immunization with the VP2.139 peptide will generate Q9:VP2.139-specific CD8 T cells in mice of non-H-2b haplotypes that express Q9 and/or Q7. H-2 s and H-2 g7 strains express Qa-2 (15,25). We confirmed Qa-2 expression in SJL (H-2s) and NOD (H-2g7) mice by positive staining of splenocytes with Qa-2 antibody (clone 1-1-2, BD Biosciences) which recognizes the Qa-2 antigen expressed by Q6, Q7, Q8, and Q9 (7, 27) (data not shown).
Adult (6–13 wk) female SJL and B6 mice, and male NOD mice, were injected in a hind footpad with 100 μg VP2.139 peptide emulsified in Complete Freund's Adjuvant (CFA) containing 1 mg/mL heat-killed Mycobacterium tuberculosis. Control mice received phosphate buffered saline (PBS) emulsified in CFA. Male NOD mice were used due to the lower incidence of a diabetogenic phenotype than observed in NOD females (1). Two weeks later, mice were boosted with 50 μg VP2.139 peptide emulsified in Incomplete Freund's Adjuvant (IFA) injected subcutaneously (s.c.) at the tail base. Control mice received PBS emulsified in IFA. Two–three weeks after the IFA boost, mice were infected with 2×105 PFU of MPyV (strain A2) intraperitoneally (i.p.). Six days later, samples of spleen, kidney, salivary gland, and heart were snap-frozen. DNA was isolated from these organs and Taqman-based quantitative (q)PCR performed as previously described (14). Single cell suspensions of RBC-lysed splenocytes prepared from each of these mice were incubated with or without 10 μM VP2.139 peptide for 5 h in the presence of Brefeldin A (GolgiPlug, BD Bioscience). Cells were surface stained, then permeabilized (Cytofix/Cytoperm, BD Bioscience) and stained intracellularly, as previously described (11). Alternatively, RBC-lysed splenocytes were treated with FcBlock (BD Biosciences) then co-stained with tetramers (described below) and antibodies, as previously described (11). Samples were run on BD FACSVerse (BD Biosciences). FACS data were analyzed using FlowJo (Tree Star) and statistical analyses were performed using Prism (GraphPad).
Engagement of CD8 coreceptors with Q9 is likely handicapped by an unusual structure and orientation of a CD8-binding loop of the Q9 α3 domain (8). Therefore, a novel Q9 tetramer having the α3 domain of H-2Kb, which efficiently binds CD8 (17), was constructed by The NIH Tetramer Core Facility. This chimeric Q9(Kb):VP2.139 tetramer stains CD8 T cells from MPyV-infected B6.Kb-/-Db-/- mice with 2-fold higher mean fluorescence intensity (MFI) than Q9:VP2.139 tetramers (Fig. 1A). To control for nonspecific tetramer binding, a chimeric Q9(Kb) tetramer containing a Q9-binding peptide from the L19 ribosomal protein (13) was also constructed by the NIH Tetramer Core Facility.
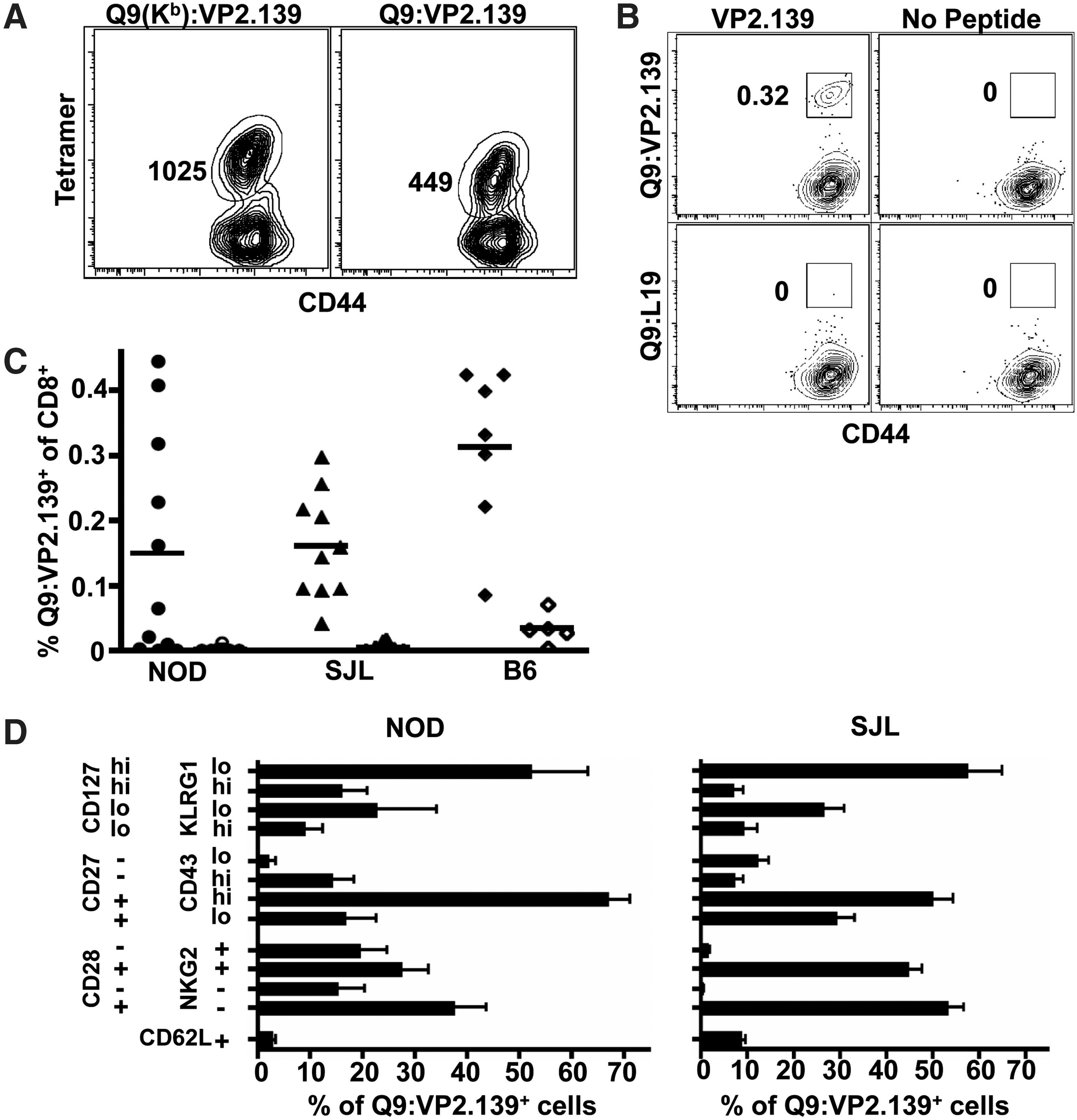
VP2.139 peptide-immunized NOD, SJL, and B6 mice generate Q9:VP2.139-specific CD8 T cell responses.
As shown in Figure 1B and C, Q9:VP2.139-specific CD8 T cells were readily detected in VP2.139-immunized mice at d 6 p.i. PBS-immunized mice showed no tetramer binding above background staining by Q9:L19 tetramers (Fig. 1B). However, the frequency of responders varied among mice of these inbred strains. Approximately 60% of NOD mice, but >80% of SJL and B6 mice had Q9:VP2.139-specific CD8 T cell responses to acute MPyV infection (Fig. 1C). It is important to note that Q9:VP2.139-specific CD8 T cells are detected in only a third of B6 mice infected with MPyV, with Q9:VP2.139 tetramer+ CD8 T cells first detected at day 8 p.i. (31). In this experimental setup, most VP2.129 peptide-immunized mice produced a Q9:VP2.139-specific CD8 T cell response by 6 days after MPyV inoculation.
Q9:VP2.139-specific CD8 T cells in VP2.139 peptide-primed NOD and SJL mice express a phenotypic profile suggestive of a memory recall response after initial priming under high inflammatory conditions. The Q9:VP2.139-specific CD8 T cells were predominantly CD62L-, as expected for a secondary effector response (Fig. 1D). Most of the Q9:VP2.139 tetramer+ CD8 T cells expressed high levels of CD127, which may be a consequence of CFA-based immunization (10). In addition, 16% (SJL) or 25% (NOD) of the Q9:VP2.139-specific CD8 T cells were KLRG-1hi, which may similarly reflect the highly inflammatory CFA priming environment (20,22). The majority of the tetramer+ cells were CD27+CD43hi, which has been associated with potential for antigenic recall (10). The Q9:VP2.139-specific CD8 T cells were predominantly CD28+, as expected for activated T cells (34). Approximately half of the tetramer+ cells expressed CD94-NKG2A/C/E (clone 20d5). In B6 mice, NKG2A is the predominant NKG2 isoform expressed by this heterodimeric receptor (36), and has been interpreted as a T cell activation marker (16,18,19). These data are consistent with our previous observations that CD94-NKG2A expression increases while CD28 expression decreases on Q9:VP2.139-specific CD8 T cells over the course of MPyV infection (11).
We previously observed that Q9:VP2.139-specific CD8 T cells mediate protection against MPyV infection in MHC class Ia knockout mice. However, viral titers were equivalent between control mice and those receiving VP2.139 peptide (Fig. 2A). Three explanations may account for this discrepancy. First, competition with conventional MHC class Ia-restricted antiviral CD8 T cells may curtail the magnitude of expansion of effector Q9:VP2.139-specific CD8 T cells needed to control acute MPyV infection. Approximately 2% of CD8 T cells in VP2.139-primed B6 mice were specific for the immunodominant Db:LT359 epitope (14) at day 6 p.i. (data not shown). Moreover, the immunization regimen used here may be insufficient to drive maximal expansion of these MHC Ib-restricted T cells. Third, Q9:VP2.139-specific CD8 T cells are defective in producing IFN-γ (31), which we recently demonstrated to be a central anti-MPyV effector activity (38). As shown in Figure 2B, over 50% of the Q9:VP2.139 specific CD8 T cells, based on tetramer staining, produce IFN-γ after VP2.139 peptide (10 μM) stimulation; this comprises just 10,000 cells in the spleen at day 6 p.i. The low magnitude Q9:VP2.139-specific CD8 T cell response elicited by the CFA-prime, IFA-boost strategy, compounded by this marked deficit in a critical antiviral cytokine, likely explains the inability of these cells to control MPyV infection.
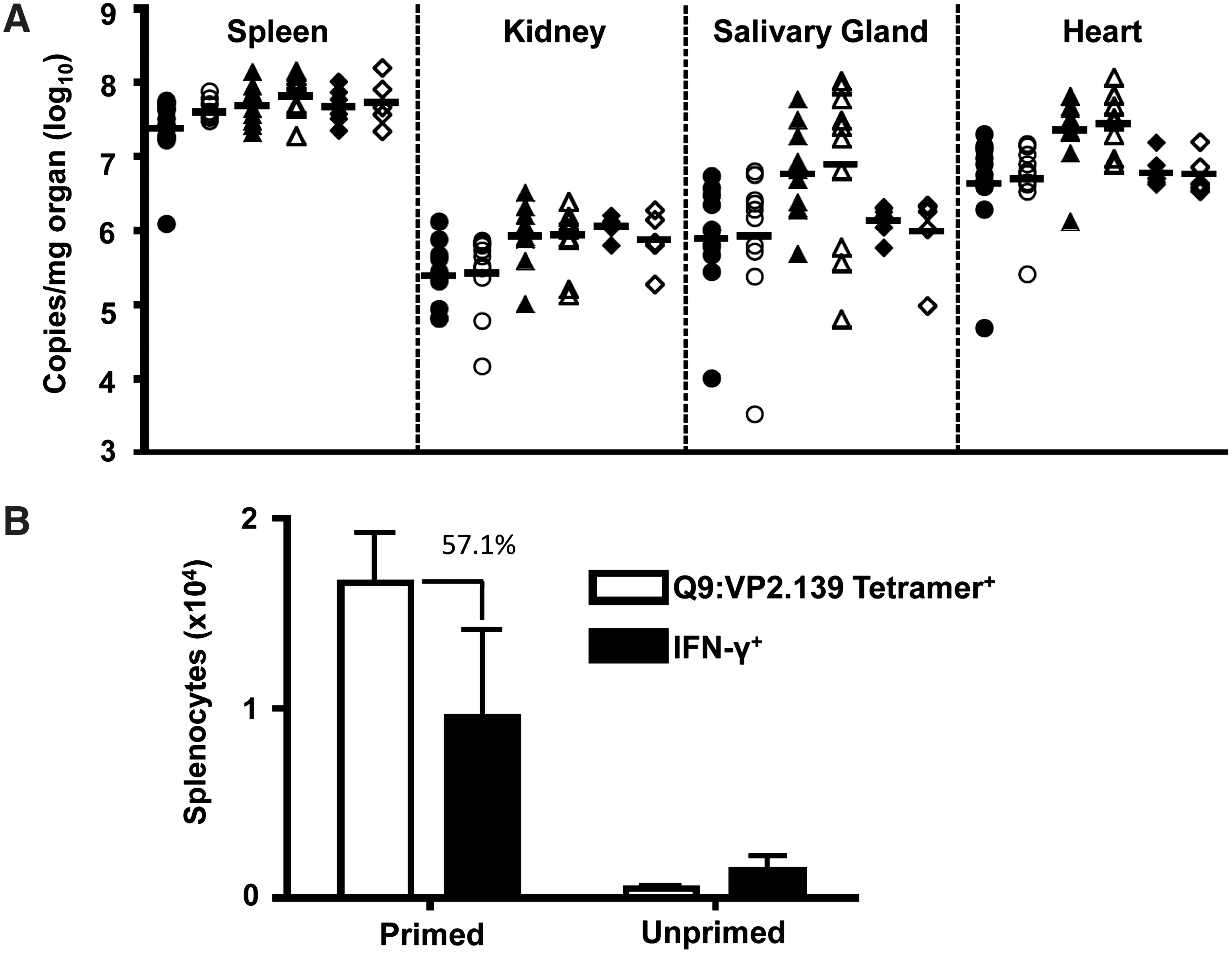
VP2.139 peptide immunized NOD mice do not control acute MPyV infection.
The diversity and level of expression of Qa-2 MHC class Ib molecules varies among inbred mouse strains due to differences in their germline representation of H2-Q6, -Q7, -Q8, and -Q9 (5). In addition, H2-Q7 and -Q9 differ by only a single amino acid outside their peptide-binding grooves and have overlapping peptide-binding repertoires (13,29,33). In contrast to B6 mice, which express both Q7 and Q9 (39), NODs express either Q7 or Q9 (15). There is also evidence that SJLs express less surface Qa-2 than B6 mice (23). It is possible that differences between B6, SJL, and NOD mice in the repertoire and expression of Qa-2 genes may affect the level of Q9:VP2.139 epitope density and explain the differences in magnitude of antigen-specific CD8 T cells observed in these mouse strains.
In summary, evidence is presented here that immunization with a single peptide presented by an MHC class Ib molecule can elicit a CD8 T cell response in mice of allogeneic MHC class Ia haplotypes. Although we were unable to demonstrate control of MPyV infection, this does not preclude the possibility that a more functionally competent and higher magnitude virus-specific CD8 T cell response restricted by other MHC class Ib molecules would confer protection against infection. In this connection, HLA-E-restricted CD8 T cells specific for cytomegalovirus, Mycobacterium tuberculosis and Salmonella typhi have been identified (9,21,24). Data described in this study support the concept that vaccines using single antigenic peptides may be useful in generating virus-specific, MHC class Ib-restricted CD8 T cell responses in individuals differing in HLA class Ia haplotypes. Work presented here underscores the need for further efforts to identify MHC class Ib-restricted CD8 T cells in humans.
Footnotes
Acknowledgments
This work was supported by National Institutes of Health Grant R01 CA139220. The authors would like to thank the NIH Tetramer Core for production of the Q9 tetramers.
Author Disclosure Statement
No competing financial interests exist.