
Abstract
Porcine respiratory and reproductive syndrome (PRRS) is an economically important disease of pigs worldwide. Currently used PRRSV vaccines provide incomplete protection. Recently, we identified Mycobacterium tuberculosis whole cell lysate (Mtb WCL) as a potent mucosal adjuvant to modified live PRRSV vaccine (PRRS-MLV). In this study, pigs were unvaccinated or vaccinated with PRRS-MLV plus Mtb WCL, intranasally, and challenged with either homologous (strain VR2332) or virulent heterologous (strain MN184) PRRSV; subsequently, euthanized at three time points post-challenge to evaluate lung immune responses. Microscopic examination of lung sections revealed reduced disruption of the lung architecture and less of interstitial pneumonia in vaccinated, compared to unvaccinated MN184 challenged pigs. The restimulated lung and peripheral blood mononuclear cells revealed increased proliferation of CD8+ lymphocytes, and in the lung homogenate increased secretion of nitric oxide was detected in vaccinated MN184 challenged pigs. In summary, the adjuvant effects of Mtb WCL to PRRS-MLV resulted in favorable anti-PRRSV immune microenvironment in the lungs to help better viral clearance.
Introduction
To control PRRS outbreaks, commonly employed vaccines include modified live PRRSV (PRRS-MLV) and killed virus vaccines. Killed vaccine does not provide adequate protection. PRRS-MLV provides satisfactory protection against homologous infections, but protection against heterologous viruses and reinfections is incomplete. There are reports of results obtained using field isolates of PRRSV and PRRS-MLV administered by either parenteral or intranasal route showing suppression of innate NK cell cytotoxic function and IFN-α production (12 –14), weak and delayed cell-mediated and virus neutralizing antibody responses (15,16), associated with prolonged immunosuppression (17). To improve the efficacy of PRRS-MLV in pigs, we explored the benefits of inducing anti-PRRSV mucosal immunity by intranasal co-administration of PRRS-MLV with a suitable mucosal adjuvant (17).
Intranasal immunization of certain vaccines has shown great promise in eliciting protective immunity against various respiratory viral infections such as bovine herpes virus-1, influenza, and parainfluenza-3 (18,19). Mucosal tissues and associated lymphoid organs contain approximately 80% of total immune cells in the body, and stimulation of the mucosal immune system leads to generation of both systemic and mucosal immune responses, resulting in effective prevention of entry and establishment of pathogens (20,21). Recently, we have demonstrated that intranasal co-administration of PRRS-MLV and adjuvant Mycobacterium tuberculosis whole cell lysate (Mtb WCL) can overcome the vaccine induced immunosuppression with concomitant increase in host innate and adaptive immune responses, associated with better protection against a virulent heterologous PRRSV MN184 challenge (12,17,22,23). We have also shown that protective anti-PRRSV immune response elicited in pigs to PRRS-MLV was mediated by the adjuvant effects of Mtb WCL, as pigs vaccinated with only PRRS-MLV failed to elicit adequate immune responses (22). In continuation, this article deals with aspects of microscopic lung pathology, proliferation of PRRSV specific CD8+ lymphocytes, and production of an important innate molecule ‘nitric oxide’ in the lungs of pigs vaccinated (PRRS-MLV+ Mtb WCL) and challenged with PRRSV.
Materials and Methods
Pigs and inoculations
Conventional Large White-Duroc crossbred specific-pathogen-free piglets weaned at 3 to 4 weeks of age were obtained from farms regularly screened and free from PRRSV, porcine respiratory coronavirus, transmissible gastroenteritis virus, and porcine circovirus type 2 were used in the study. Before beginning of our study, pigs were tested to confirm absence of PRRSV antibodies by ELISA. Throughout the duration of the study, pigs received food and water ad libitum. All inoculations including PRRSV challenge was performed by intranasal delivery. Animals were maintained, samples collected, and euthanized in our large animal BSL2 facility as per the protocol approved by the Institutional Animal Care and Use Committee (IACUC) and Institutional Biosafety Committee (IBC), The Ohio State University, Ohio. Forty pigs were allocated to one of five groups: group 1, mock pigs (n=4) inoculated with vehicle (normal saline) and DMEM; other four groups (n=9 per group), groups 2 and 3, unvaccinated; groups 4 and 5, vaccinated [PRRS-MLV (Boehringer Ingelheim Vetmedica, Inc.)+Mtb WCL]. Groups 2 and 4 were challenged with a homologous parenteral PRRSV strain VR2332 (2×106 TCID50 per pig), and groups 3 and 5 were challenged with virulent heterologous PRRSV MN184 (24) (1×106 TCID50 per pig) in 2 mL on day 21 post-immunization. Nine pigs each under groups 2 to 5 were euthanized at three time points at 15, 30, and 60 day post-challenge (DPC) (n=3 per group). Mock inoculated pigs (n=4) were euthanized together prior euthanizing any virus challenged animals.
Microscopic lung pathology
Lung tissue samples collected from pigs was fixed in 10% neutral buffered formalin and processed into paraffin blocks, three μm sections were cut and stained with haematoxylin and eosin as described previously (25). The lung sections were examined in a blindfolded fashion by a board certified veterinary pathologist to determine the lung lesion scores by virus-induced pathology (26).
Carboxyfluorescein diacetate succinimidyl ester proliferation assay
To determine PRRSV specific proliferation of lymphocytes, lung mononuclear cells (LMNC) and peripheral blood mononuclear cells (PBMC) were isolated as described (27 –29). Cells were stained with an irreversible cytosolic dye, (carboxyfluorescein diacetate succinimidyl ester, CFSE), as described previously (30). Briefly, five million cells were washed and stained with CFSE at 10 μM concentration in PBS and incubated for 15 min at room temperature. The labeling reaction was stopped using ice-cold RPMI containing 10% FBS, washed twice, and resuspended with the medium and incubated at 37°C for 30 min, and adjusted the cell numbers to 5×105 cells per well in a 96-well flat bottom plate. The cells were restimulated with semi-purified PRRSV antigens (20 μg/mL) of VR2332 and MN184 strains, obtained by ultracentrifugation of clarified PRRSV infected MARC 145 cell culture supernatant. Cells untreated or treated with Concavalin A were used as negative and positive control, respectively. Following 3 days of incubation, cells were washed and stained with R-phycoerythrine conjugated anti-pig CD8 antibody (clone 76-2-11, BD Biosciences). Finally, cells were fixed and 50,000 to 100,000 events acquired using the FACS AriaII (BD Biosciences) flow cytometer and analyzed for CFSE and PE stained lymphocytes. The cell proliferation data was analyzed by FlowJo software (Tree Star, Inc. OR) to obtain the proliferation index (PI) of each sample. The PI denotes the average number of cell divisions the responding lymphocytes underwent after subtracting from the background (initial population of cells at generation ‘0’).
Griess assay
Nitric oxide production in the lungs was determined by measuring nitrite levels, which is formed as nitric oxide breakdown product using a commercial kit (Promega). Lung homogenates were prepared from the lung tissue samples as described previously (13). Samples were treated with a sulfanilamide solution for 5–10 min in the dark, and subsequently with N-1-napthylethylenediamine dihydrochloride (NED) solution. The resulting reaction product was measured at an absorbance between 520 and 550 nm, and the absorbance was calculated by generating a nitrite standard reference curve.
Statistical analysis
All data were expressed as the mean±SEM of three pigs. Statistical analyses were performed using two-sample t-test to establish differences between vaccinated and unvaccinated groups. Statistical significance was assessed as p<0.05.
Results and Discussion
Porcine reproductive and respiratory syndrome virus has evolved into a major swine disease over the last 2 decades. We have recently demonstrated the benefits of intranasal delivery of PRRS-MLV with a potent mucosal adjuvant, Mtb WCL, in stimulating favorable anti-PRRSV immune responses against virulent heterologous (MN184) PRRSV (12,22). As a potent mucosal adjuvant, we used mechanically lyzed endotoxin-free WCL of Mtb which contains predominantly soluble intracellular bacterial proteins and is almost free from toxic cell wall lipid components (31) (Torrelles JB, personal communication). Clinically, pigs administered with Mtb WCL did not show any signs of side effects. In vaccinated (PRRS-MLV plus Mtb WCL) pigs, reduced gross lung pathology and significantly reduced viremia were attributed to adjuvant mediated innate and PRRSV specific adaptive immune responses (22). Here, in the same set of pigs, we are providing additional important information in the lung microenvironment with reference to status of PRRSV specific CD8+ lymphocytes, nitric oxide production, and microscopic lung pathology.
Histological examination has revealed reduced lung lesions (Fig. 1), indicated by reduced infiltration of mononuclear cells and interstitial pneumonic changes in vaccinated compared to unvaccinated MN184 challenged pigs (Fig. 1A). Further, in vaccinated, MN184 challenged pigs reduced (although not statistically significant) trend in lung lesion scores was detected at DPC 15 and 30 compared to unvaccinated pigs (Fig. 1Bi). Due to the low virulence nature of VR2332 strain, the inflammatory changes in the lungs were lesser compared to MN184 challenged pigs (Fig. 1Bi and Bii).
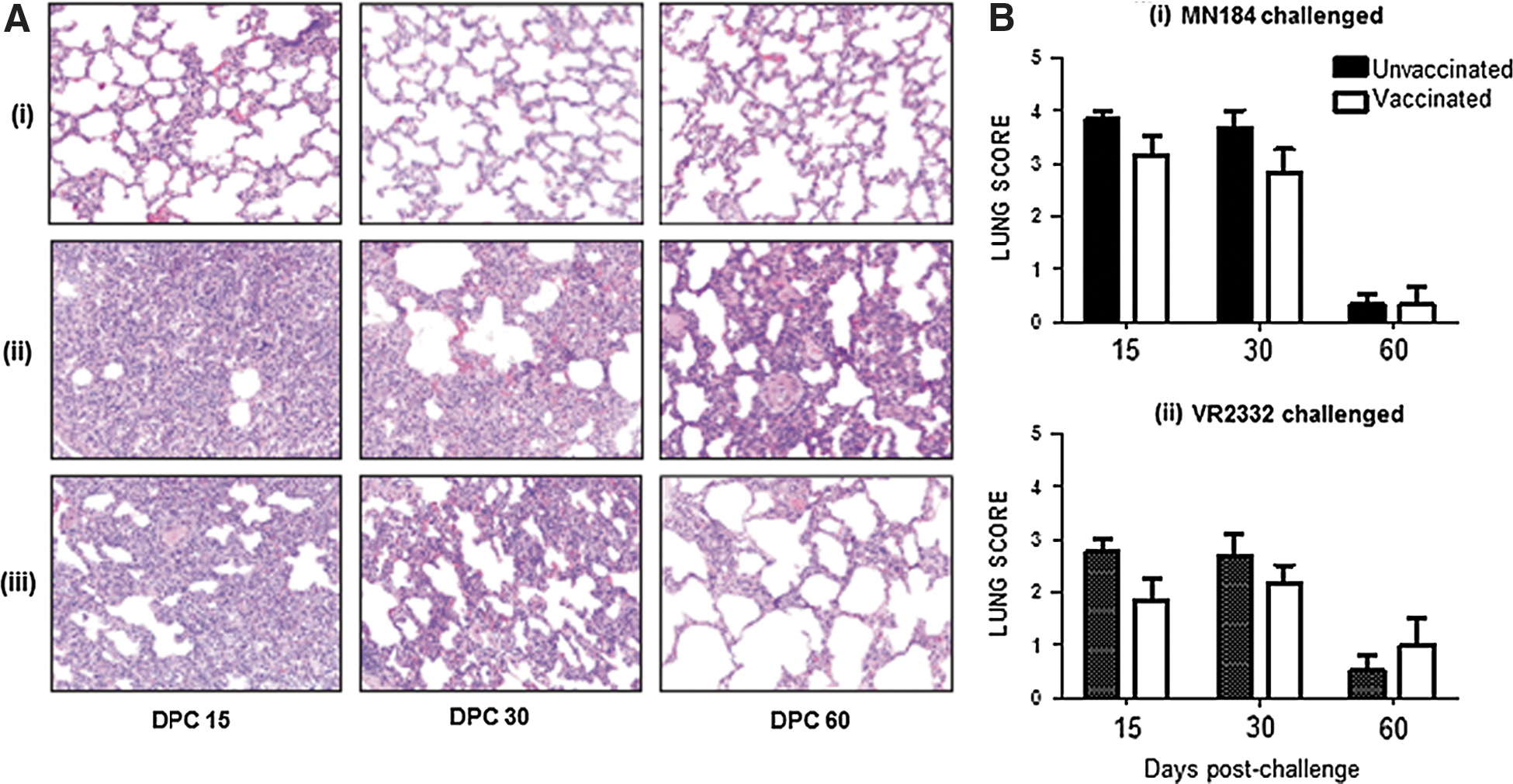
Reduced infiltration of inflammatory cells and lung pathology in vaccinated PRRSV challenged pigs. Pigs were unvaccinated or vaccinated with PRRS-MLV plus Mtb WCL intranasally and challenged with PRRSV strain MN184 or VR2332, and euthanized at DPC 15, 30, and 60. Lung sections were stained with H&E:
The cell-mediated immunity against virus infections is contributed by different lymphocyte subsets, in particular contributed significantly by CD8+ cytotoxic T lymphocytes (CTL) and activated Th/memory CD4+CD8+ T cells (19,32). PRRSV-specific lymphocytes present in the PBMC of infected pigs proliferate from 4-week post-infection (33,34). However, so far, knowledge related to the status of CD8+ lymphocytes in the lungs of PRRSV infected and/or vaccinated pigs is limited. Since the PRRSV replicates in the lungs of infected pigs, such information is important. In our study, increased proliferation of total PRRSV-specific lymphocytes in the lungs (LMNC) and blood (PBMC) of vaccinated compared to unvaccinated, and either PRRSV strain challenged pigs was detected (Fig. 2A and data not shown). Further, significantly increased proliferation of CD8+ lymphocytes in the PBMC of MN184 challenged and LMNC of VR2332 challenged pigs restimulated with MN184 antigens was detected (Fig 2B, Cii and Ciii). In pigs, apart from CD8+ T cells, CD4+CD8+ T lymphocytes are abundantly present, and they possess memory, activated T-helper and cytolytic properties (32,35 –37). Since the antigen-specific proliferation occurs only by CD4+ and CD8+ T cells, our results indicated that PRRSV-specific proliferation occurred in both CD8+ and CD4+CD8+ T lymphocytes.
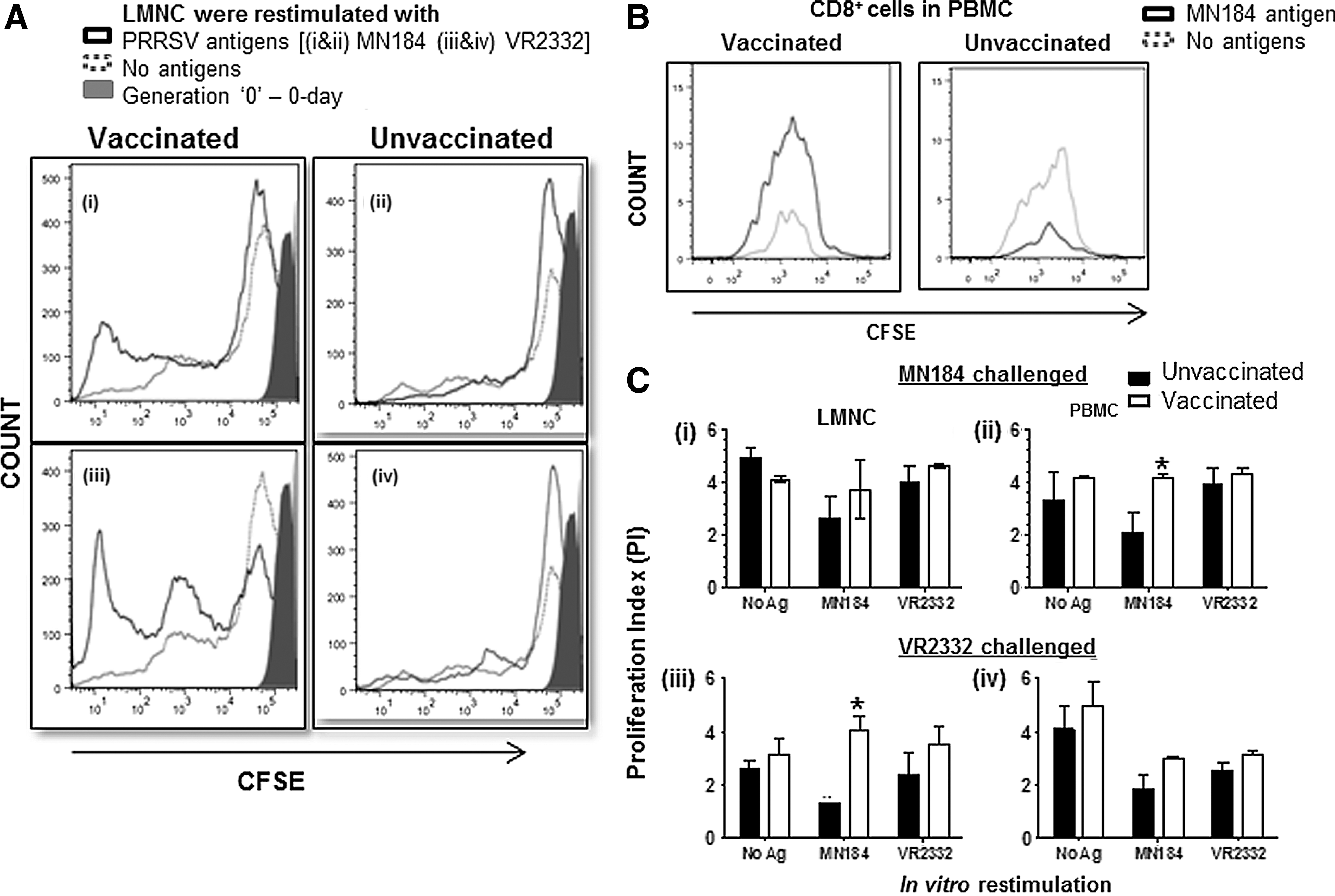
Vaccinated pigs had enhanced proliferation of CD8+ lymphocytes.
Nitric oxide (NO’) is an important immune molecule involved in both innate and adaptive immunity. It helps in the clearance of intracellular pathogens by inhibiting viral replication and plays a major role in lung immunity and virus clearance. It has been shown to inhibit replication of influenza virus and herpes simplex virus type1 (38 –40). A large amount of pulmonary NO’ is produced by the epithelial lung inducible nitric oxide synthase (41), which acts on L-arginine to produce NO’. To determine the production of NO’ in the pig lung, lung homogenates (which represent the lung parenchyma) were prepared and used in the analyses. A significant increase in the production of NO’ in the lungs of pigs received adjuvanted vaccine and challenged with virulent PRRSV MN184 at DPCs 30 and 60 was detected (Fig. 3A). Levels of NO’ production in PRRS-MLV alone vaccinated and MN184 challenged pigs was high and it was comparable to unvaccinated virus challenged animals at DPC 30 (data not shown).
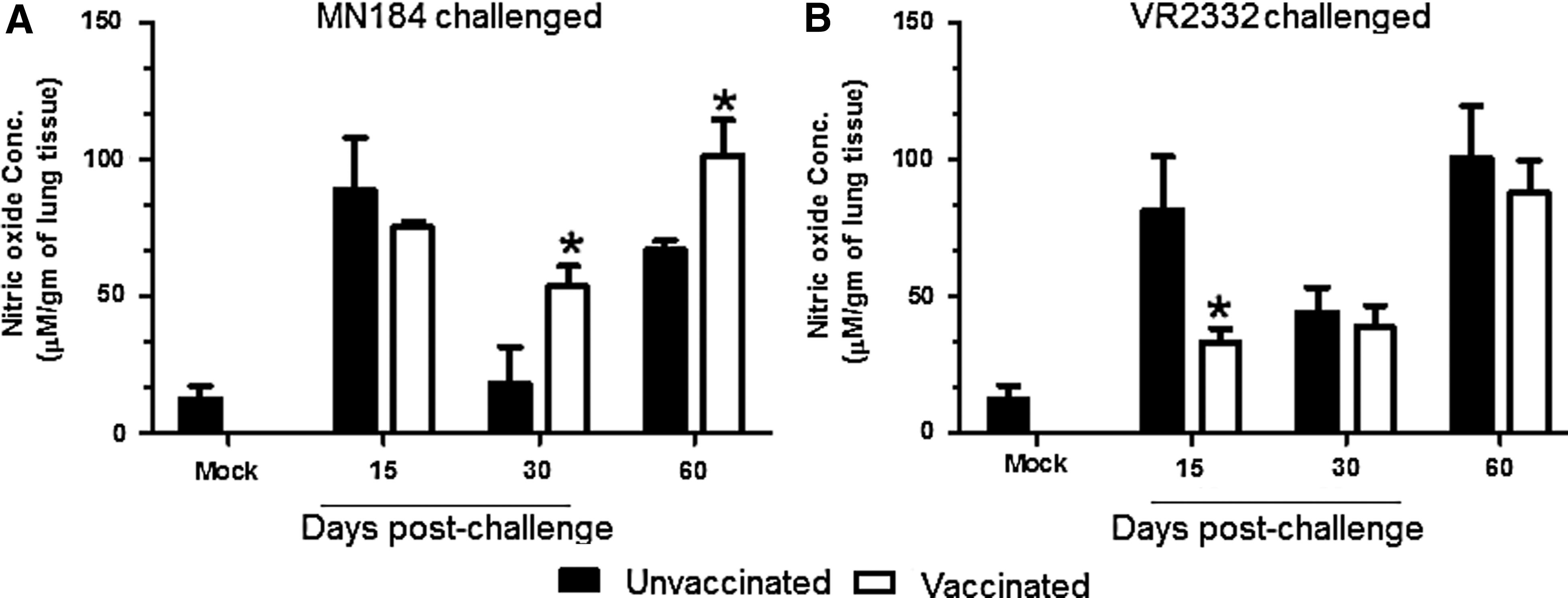
Elevated nitric oxide production in vaccinated pigs. Pigs were unvaccinated or vaccinated (PRRS-MLV+Mtb WCL) intranasally and challenged with PRRSV strain:
In the lungs of pigs vaccinated and challenged with VR2332 strain, an opposite trend in the NO’ production was detected at DPC 15, and at DPC 30 and 60 the levels were comparable between the groups (Fig. 3B), which could be due to differential immune responses elicited by the avirulent VR2332 strain (42). Earlier reports have demonstrated the absence of NO’ in bronchoalveolar lavage fluid of a mild PRRSV strain SD23983 infected pigs (43), and production of inducible nitric oxide synthetase (iNOS) in alveolar macrophages (44). However, pulmonary epithelial cells in the respiratory viral infections contribute significantly in the production of NO’ (45). In our study, we used lung homogenates, and in pigs vaccinated with PRRS-MLV plus Mtb WCL and challenged with a virulent heterologous PRRSV strain MN184, significantly increased NO’ production was detected in the lung homogenate.
Earlier, we have shown an increased secretion of reactive oxygen species (ROS) by bronchoalveolar lavage fluid cells and PBMC of unvaccinated compared to vaccinated (PRRS-MLV plus Mtb WCL) MN184 challenged pigs at three DPCs (23). While ROS and reactive nitrogen species (RNS) are needed for antimicrobial activity, their overproduction leads to bystander cell death, inflammation, and tissue injury (46,47). NO’ is one of the RNS, and it reacts with oxygen radicals, leading to production of more ROS and RNS. A balance between NO’ and ROS will influence the outcome (48). Thus, it is possible that PRRS-MLV plus Mtb WCL vaccination optimally increased the NO’ levels in the lungs to virulent MN184 challenge, which however did not lead to lung injury, rather helped in better anti-PRRSV immune response compared to control pigs (22).
In conclusion, intranasal co-administration of PRRS-MLV and Mtb WCL elicits favorable immune responses against a virulent heterologous PRRSV challenge; indicated by reduced gross lung pathology, early lung viral clearance, activation of virus specific immune cells, early virus neutralizing antibody generation, increased Th1, and reduced immunosuppressive cytokine production (22). In addition, in this study in the lungs of pigs vaccinated and MN184 challenged reduced microscopic lesions, increased proliferation of virus specific CD8+ T cells, and enhanced NO’ production were detected. Thus, our results further supported the favorable anti-PRRSV immune responses in the pig lung microenvironment mediated by the adjuvant Mtb WCL to PRRS-MLV.
Footnotes
Acknowledgments
This research was supported by National Pork Board and USDA-NIFA PRRS CAP2 award 2008-55620-19132 to GJR. Salaries were provided by the state and federal funds appropriated to OARDC, The Ohio State University. We would like to thank Ms. Ruth Patterson for her help in the preparation of immune cells. Dr. Juliette Hanson, Todd Root, Mathew Weeman, Drs. Mahesh Khatri and Hadi Yassine for their help in animal studies. Drs. Eric Nelson and Michael Murtaugh provided PRRSV reagents.
Author Disclosure Statement
No competing financial interests exist.