
Abstract
Bacteriophages are viruses that infect bacteria. It was shown that bacteriophage therapy is an effective method of combating bacterial infections, including infections caused by antibiotic-resistant bacterial strains. One of the main obstacles to widespread use of phage preparations is limited knowledge regarding the influence of bacteriophages on human organisms. In our study, we evaluated whether application of phage preparations impair bactericidal activities of human phagocytes (granulocytes and monocytes). In our study, we used preparations of phages T2 and T4 specific to Escherichia coli and A3 phage specific to Staphylococcus aureus. We found that bacteriophage preparations do not influence intracellular killing of bacteria by human phagocytes. The effect is irrespective of phage preparation type (lysate, purified phage preparation), phage titer of the preparation, and whether bacteria phagocytosed by phagocyte cells are sensitive or insensitive to phage (bacteriophages homologous and heterologous to bacteria). Although the results of our study are preliminary, they support previous data indicating safety of therapeutic application of phages.
Introduction
Application of bacteriophages in medicine, biotechnology, and industry constantly increases (food safety, protection of biomedical equipment, plant protection, control of water-borne pathogens, and control of environmental microflora) (10,11). It is significant that lately, the number of companies involved in the commercialization of phage preparations has remarkably increased (12,13). A few phage products have been classified by the FDA (Food and Drug Administration) as GRAS (Generally Regarded as Safe), registered by the EPA (Environmental Protection Agency), and approved for use by the USDA (United States Department of Agriculture) (12).
One of the reasons why application of phage preparations was limited after their discovery was the obscure nature of these viruses. Nowadays, knowledge regarding phage biology and interactions with prokaryotic cells and eukaryotic organisms is much more extensive. Bacteriophages are believed to have no natural tropism to eukaryotic cells; however, studies show that such interactions may occur (14 –22). Unfortunately, the knowledge regarding interactions between bacteriophages and immune cells is still insufficient.
In our work, we evaluated the influence of phage preparations on intracellular killing (ICK) of bacteria by human phagocytes (granulocytes and monocytes). Phagocytes are cells that play a crucial role in human defense mechanisms. Our studies concern in particular patients subjected to phage therapy, who often have phagocyte dysfunctions (23,24). Determining whether phage preparations have a negative impact on these cells is of great importance.
The study was conducted in two experimental models: in the first one, the influence of phages homologous to bacteria phagocytosed by phagocytes was studied (bacteria sensitive to phage), and in the second, the influence of phages heterologous to bacteria was studied (bacteria insensitive to phage). The second experimental model reflects infection caused by more than one bacterial strain, when the patient receives a preparation of phage sensitive to only one bacterial strain (e.g., when it is not possible to select a phage active toward the second pathogen). It also concerns the situation when bacteria are not de facto sensitive to phage received by the patients, for example, in the case of phage resistance or in the case of infection which has not been already detected.
The results presented in our article indicate that phage preparations do not influence ICK of bacteria by human phagocytes irrespectively of phage preparation type, phage titer in the preparation, and whether the phage is homologous or heterologous to bacteria phagocytosed by phagocytes. This may confirm data indicating safety of phage therapy. Although the results of our studies are preliminary, we believe that they will contribute to propagation of phage usage.
Material and Methods
Bacteriophages
Bacteriophages T2, T4, and A3 were obtained from the Polish Collection of Microorganisms in the Institute of Immunology and Experimental Therapy (IIET), Polish Academy of Sciences (PAS). Phages were propagated with Escherichia coli B and Staphylococcus aureus (IIET Bacterial Collection). Bacteriophage lysates were prepared according to the method described by Carlson and Miller (25). In most experiments, we used preparations with phage titer 1×109 pfu/mL. To determine whether phage titer correlates with phages' influence on ICK of bacteria by phagocytes, in further experiments we used lysates with phage titer 1×1010 pfu/mL; 1×108 pfu/mL; 1×106 pfu/mL, and 1×104 pfu/mL.
We also used purified phage preparations of phages T2, T4, and A3. Purified preparations were obtained according to the procedure described by Boratynski et al. (26). Endotoxin level in preparations was determined with Limulus Amebocyte Lysate Test Kit (LAL) (BioWhittaker, Radnor, PA). Additionally, we used a purified preparation of A3 phage obtained from the Institute of Biotechnology, Sera and Vaccines, BIOMED S.A. Both purified A3 phage preparations had the same phage titer (1×109 pfu/mL) but they differed in lipopolysaccharide concentration (A3 IIET–23 Eu/mL; A3 BIOMED–3 Eu/mL), purification technique (the procedure of A3 phage (BIOMED) purification is protected by patent) and enterotoxin level.
Phage titer in all preparations was determined with the double agar method ( 27).
Phagocytes
Granulocytes and monocytes were isolated from buffy coats obtained from blood of volunteer donors from the Blood Donor Centre at the Military Clinical Hospital with Policlinic in Wroclaw. Vitality of isolated cells was >98%.
Bacterial strains
Standard E. coli B and Staphylococcus aureus strains used for phage propagation were from Polish Collection of Microorganisms (Institute of Immunology and Experimental Therapy, Poland). Clinical strains of E. coli, Pseudomonas aeruginosa, S. aureus, and E. faecalis were isolated from patients qualified to the phage therapy in Phage Therapy Unit in Wroclaw under the protocol of “Experimental phage therapy of drug-resistant bacterial infections, including MRSA infections” (23).
Lipopolysaccharide
Lipopolysaccharide (LPS) of E. coli B was isolated by Dr. Kinga Switala–Jelen according to the procedure described by Westphal (28). LPS was used as a control of the potential activity of bacterial residues in phage preparations.
Preparation of bacteria
Standard bacterial growth curve
Serial dilutions (10−1–10−9) of 20-h bacterial culture were prepared. The number of bacteria was determined with the standard plate count technique, and simultaneously, the absorbance of each sample was measured. The standard bacterial growth curve was determined: f(N)=A (N – number of bacteria in sample per mL; A, absorbance).
Preparation of bacterial suspension
20-h bacterial culture on agar slant was suspended in 3 mL of NaCl (0.9%) and centrifuged (∼1300 g, 4°C, 5 min). Supernatant was discarded and cells were washed twice in 10 mL of NaCl (0.9%). Finally, bacteria were suspended in 1 mL of NaCl (0.9%) and optical density (ODx) of the sample was measured. The number of bacteria in the sample was determined according to a standard bacterial growth curve. A suspension of 1010 bacteria/mL was prepared.
Opsonization of bacteria
Human serum collected during phagocyte isolation was added to the cell suspension to a final concentration of 10%. The suspension was incubated for 25 min (37°C). After incubation, bacteria were washed three times with NaCl (0.9%) (∼1300 g, 4°C, 10 min) and finally suspended in 1 mL of NaCl (0.9%).
Preparation of phagocyte suspension
Phagocytes were isolated from buffy coats (enriched with 2500 UI/mL heparin) of volunteer human blood donors on a double gradient of Histopaque 1.119 g/mL and Histopaque 1.077 g/ml (Sigma-Aldrich, Poland) according to the procedure described by Boyum (29). The fraction of granulocytes and/or mononuclear cells was collected and washed in NaCl (0.9%) (∼600 g, 4°C, 5 min). Lymphocytes have no phagocytic activity, so in the experiments a complete suspension of mononuclear cells was used, assuming that only monocytes reveal phagocytic activity. The average content of monocytes in the suspension of mononuclear cells determined with FACSCalibur (BD Biosciences, San Jose, CA; marker CD14) was 14%. The deposit of phagocytes was suspended in 1 mL of NaCl (0.9%). The number of cells was determined by counting with trypan blue in a Bürker chamber. A suspension of 1×107 cells/mL was prepared.
Influence of phage preparations on ICK of bacteria by human granulocytes and monocytes
The following samples were prepared:
N: sample in which phagocytes and bacteria were suspended only in NaCl (0.9%) (without serum in suspension)—in these conditions the process of ICK of bacteria occurs in suboptimal conditions;
S: sample in which phagocytes and bacteria were suspended in NaCl (0.9%) with serum (10%)—in these conditions the process of ICK of bacteria occurs in optimal conditions;
A: sample in which phagocytes were incubated with phage preparation or control preparations: control medium (CM), phosphate-buffered saline (PBS), or LPS solution before introducing the bacteria (serum in suspension);
A+BP: control of phage activity: bacteriophages and bacteria (without phagocytes and serum in suspension);
200 μL of phage preparation was added to 200 μL of phagocyte suspension (in control groups instead of phage preparation 200 μL of LPS, CM, or PBS was added, depending on the experimental model) and incubated for 1 h (37°C). In samples N and S, cells were incubated only with NaCl (0.9%). A phage activity control sample (A+BP) was also prepared: 200 μL of phage preparation and 200 μL of NaCl (0.9%). Table 1 presents the pre-incubation scheme in all samples.
+=presence in experimental model; −=absence in experimental model.
After pre-incubation, suspensions (except A+BP) were centrifuged (∼200 g, 4°C, 5 min), supernatant was gently removed and cells were suspended in 1200 μL of NaCl (0.9%). To each sample (excluding sample N), serum was added to a final concentration of 10%. Cell suspensions were thoroughly mixed and bacteria were added (200 μL of bacterial culture prepared as indicated above for each sample). The ratio of phagocyte:bacteria was 1:1000.
Three minutes after bacteria addition (t0), 500 μL of each sample (N, S, A) was removed to a new test tube, and 700 μL of cool NaCl (0.9%) (4°C) was added to stop ICK of bacteria. Both initial and collected samples were centrifuged (∼200 g, 4°C, 5 min) in order to remove nonphagocytosed bacteria. Collected samples were washed three times in cool NaCl (0.9%) and then suspended in 1 mL of 0.01% bovine albumin in order to disrupt cells and release intracellular bacteria. Serial dilutions of obtained suspension were prepared. The number of bacteria was estimated with the standard plate count method.
Initial samples were washed twice in NaCl (0.9%) (room temperature) and then suspended in 1 mL of NaCl (0.9%) with serum (exception: sample N—without serum). Serum final concentration in suspensions was 10%. Samples were incubated at 37°C. Total incubation time (t1) in the case of granulocytes was 1 h and in the case of monocytes 1.5 h (it was determined that after this time ICK of bacteria by these cells is optimal). Total incubation time was measured from the moment of bacteria addition. After t1, 500 μL of the each suspension was removed to a new test tube, and 700 μL of cool NaCl (0.9%) (4°C) was added to stop ICK of bacteria by phagocytes. The collected sample was centrifuged (∼200 g, 4°C, 5 min). The deposit was suspended in 1 mL of 0.01 % albumin. Serial dilutions of bacterial suspension were prepared. The number of bacteria was determined with the standard plate count technique.
Sample A+BP was centrifuged at no stage of the experiment. From the moment of bacteria addition, it was incubated at 37°C. At t0 and t1, 200 μL of suspension was collected and serial dilutions were prepared. The number of bacteria was determined with the standard plate count method.
Studies of ICK of clinical strains of bacteria by healthy donors
Blood samples were obtained from six healthy blood donors. Study of the intracellular killing of clinical strains by phagocytes isolated from healthy donors was prepared similar to procedure for sample N for in vitro experiments. Mean intracellular killing [expressed in %], and standard error of the mean (SE) were calculated. Differences between groups were analyzed using t-test (p<0.05 was considered significant).
Determination of proportional ICK of bacteria by phagocytes
Diluted bacterial suspensions were seeded on agar plates in two repetitions. After 20-h incubation, bacterial colonies were counted. The number of bacteria in t0 and t1 samples was determined:
CB=NC×2×10×D
CB, number of bacteria in sample (per ml)
NC, number of bacterial colonies
D, inversed dilution, from which the number of colonies was counted
2, 10, dilution factors
ICK (%) of bacteria by phagocytes was determined according to equation:
ICK (%), proportional ICK of bacteria by phagocytes (%)
Ct0, Ct1, the number of bacteria in samples t0 and t1 (determined as indicated above).
Results
Influence of phage preparations on ICK of bacteria by human granulocytes
Phages homologous to bacteria phagocytosed by granulocytes (bacteria sensitive to phages)
Table 2 presents experimental models applied in studies of phage influence on ICK of bacteria by granulocytes (phages homologous to bacteria).
ICK (%) of bacteria killed by granulocytes was determined according to the procedure described in Material and Methods. ICK (%) of bacteria killed by granulocytes incubated with phage lysates T2 (T2L) and T4 (T4L), purified preparations of phages T2 (T2P) and T4 (T4P), A3 phage lysate (A3L), and purified preparation of A3 phage (A3P) is presented on Figures 1 –3. The influence of two A3 purified phage preparations (A3(IIET), A3(BIOMED)) on ICK of bacteria by granulocytes was evaluated (Fig. 4a and 4b). Differences between preparations A3 (IIET) and A3 (BIOMED) have been described in Material and Methods. All figures below present the arithmetic mean from three experiments and standard deviation. Descriptions of samples are as indicated in Material and Methods.
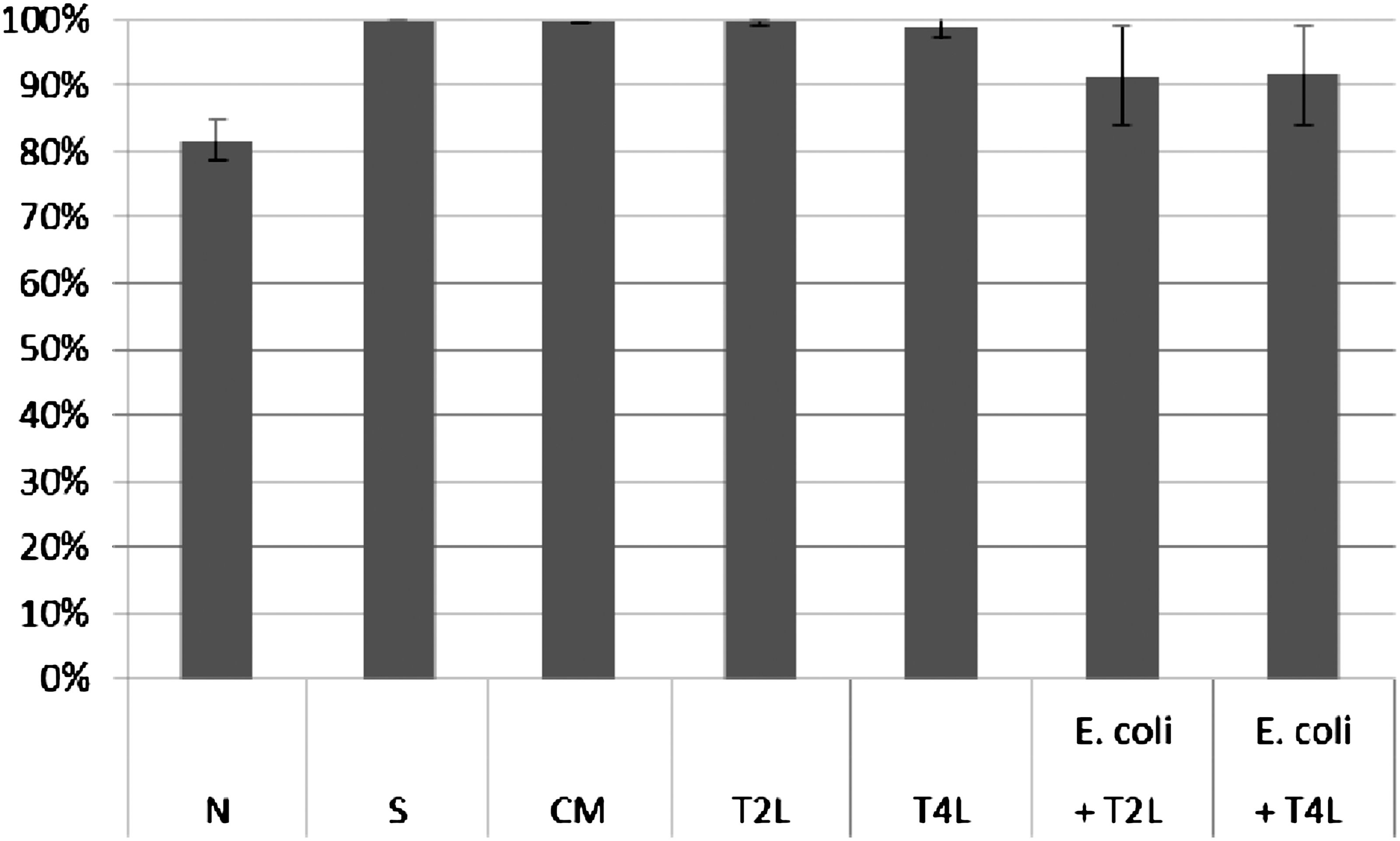
Influence of T2 and T4 lysates on ICK of bacteria E. coli by human granulocytes. Figure presents ICK (%) of bacteria by granulocytes after 1 h. Last two columns on the right present proportional lysis of bacteria E. coli (%) by T2 and T4 phage lysates (no phagocytes in suspension) after 1 h.
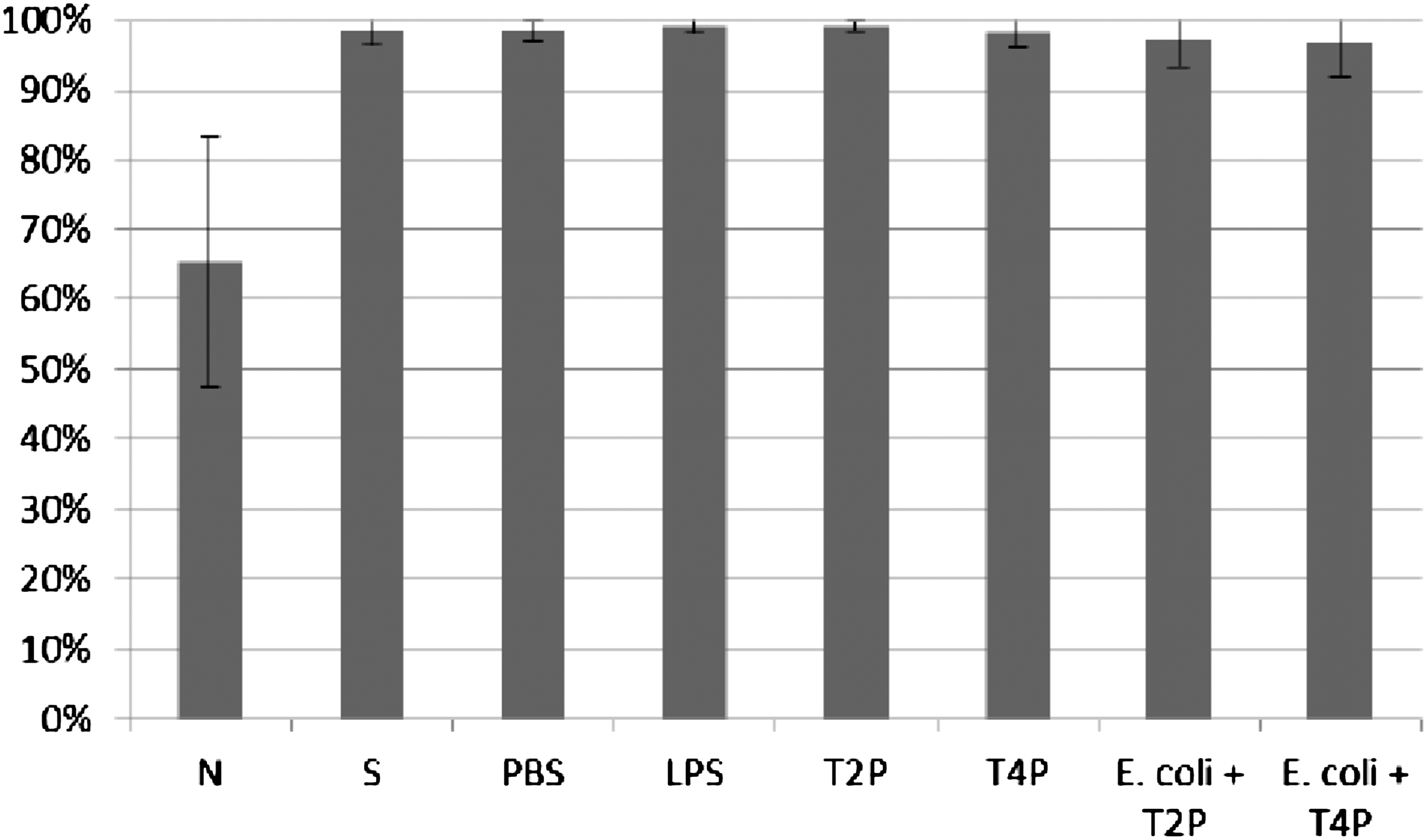
Influence of T2 and T4 purified preparations on ICK of bacteria E. coli by human granulocytes. Figure presents ICK (%) of bacteria by granulocytes after 1 h. Last two columns on the right present proportional lysis of bacteria E. coli (%) by T2 and T4 purified preparations (no phagocytes in suspension) after 1 h.
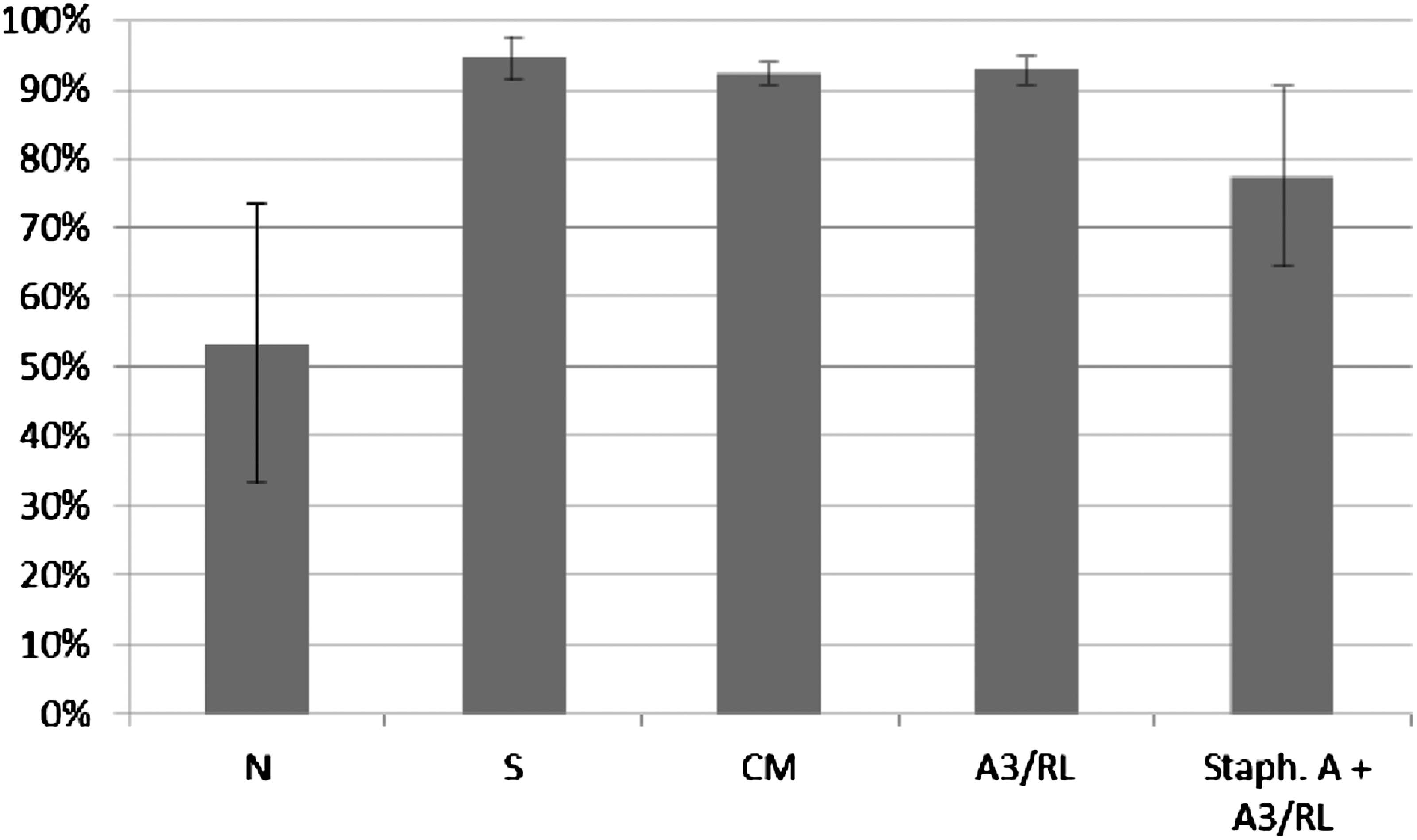
Influence of A3 lysate on ICK of bacteria Staphylococcus aureus by human granulocytes. Figure presents ICK (%) of bacteria by granulocytes after 1 h. Last right column presents proportional lysis of bacteria Staphylococcus aureus (%) by A3 lysate (no phagocytes in suspension) after 1 h.

In experiments in which the influence of phages homologous to bacteria phagocytosed by granulocytes was studied, ICK (%) of bacteria by granulocytes was comparable to ICK of bacteria in the positive control (S), CM in the case of bacteriophage lysates, and PBS and LPS in the case of purified phage preparations. No differences in activity of A3 (IIET) and A3 (BIOMED) purified preparations were observed. Preparations of phages homologous to bacteria do not influence ICK of bacteria by human granulocytes.
Phages heterologous to bacteria phagocytosed by granulocytes (bacteria insensitive to phages)
Table 3 presents experimental models applied in studies of phage influence on ICK of bacteria by granulocytes (phages heterologous to bacteria).
ICK (%) of bacteria killed by granulocytes was determined according to the procedure described in Material and Methods. ICK (%) of bacteria killed by granulocytes incubated with phage lysate T4 (T4L) and purified preparation of phage T4 (T4P) is presented on Figures 5 and 6. Figures present the arithmetic mean from three experiments and standard deviation. Descriptions of samples are as indicated in Material and Methods.
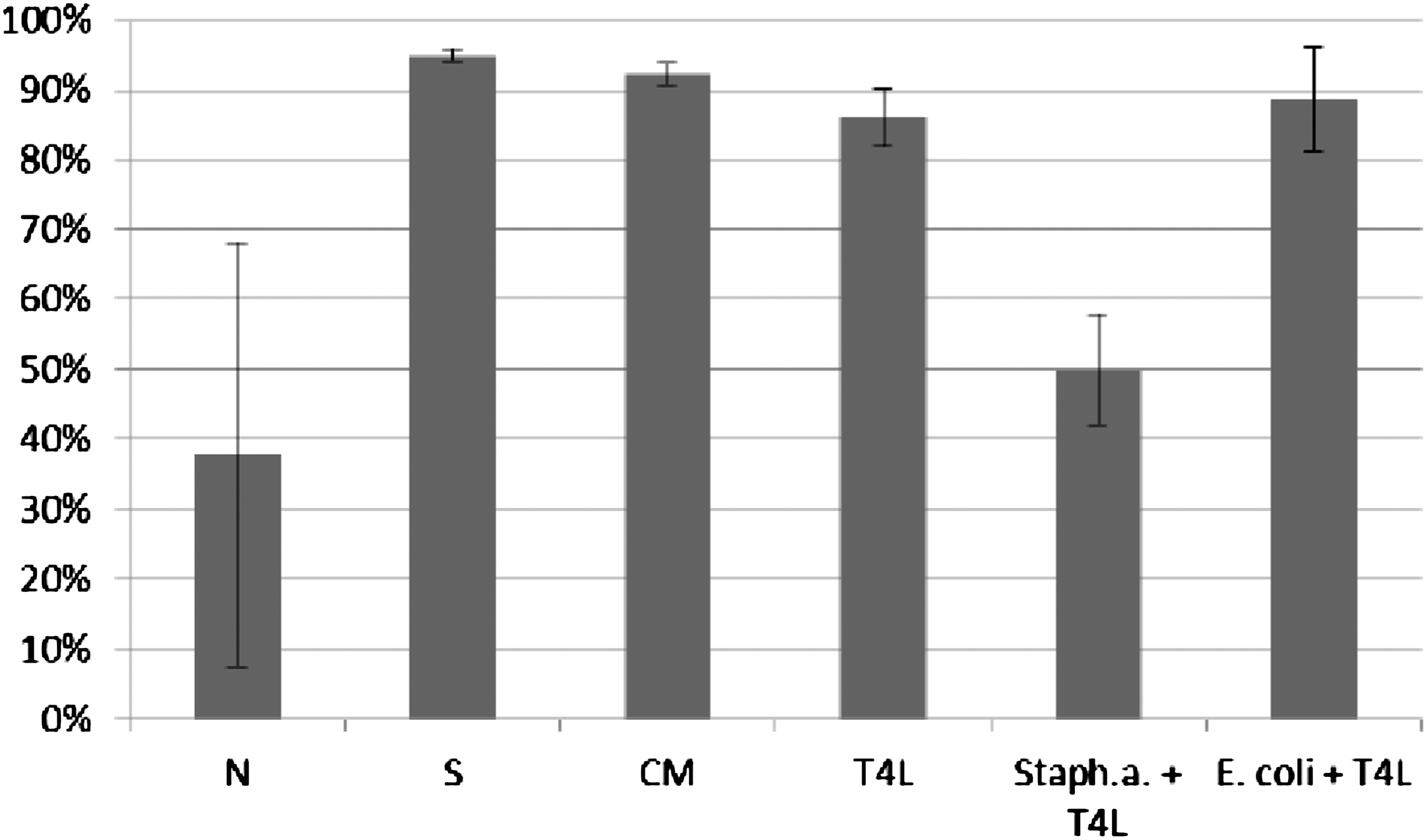
Influence of T4 lysate on ICK of bacteria Staphylococcus aureus by human granulocytes. Figure presents ICK (%) of bacteria by granulocytes after 1 h. Last two columns on the right present proportional lysis of bacteria E. coli and Staphylococcus aureus (%) by T4 lysate (no phagocytes in suspension) after 1 h.
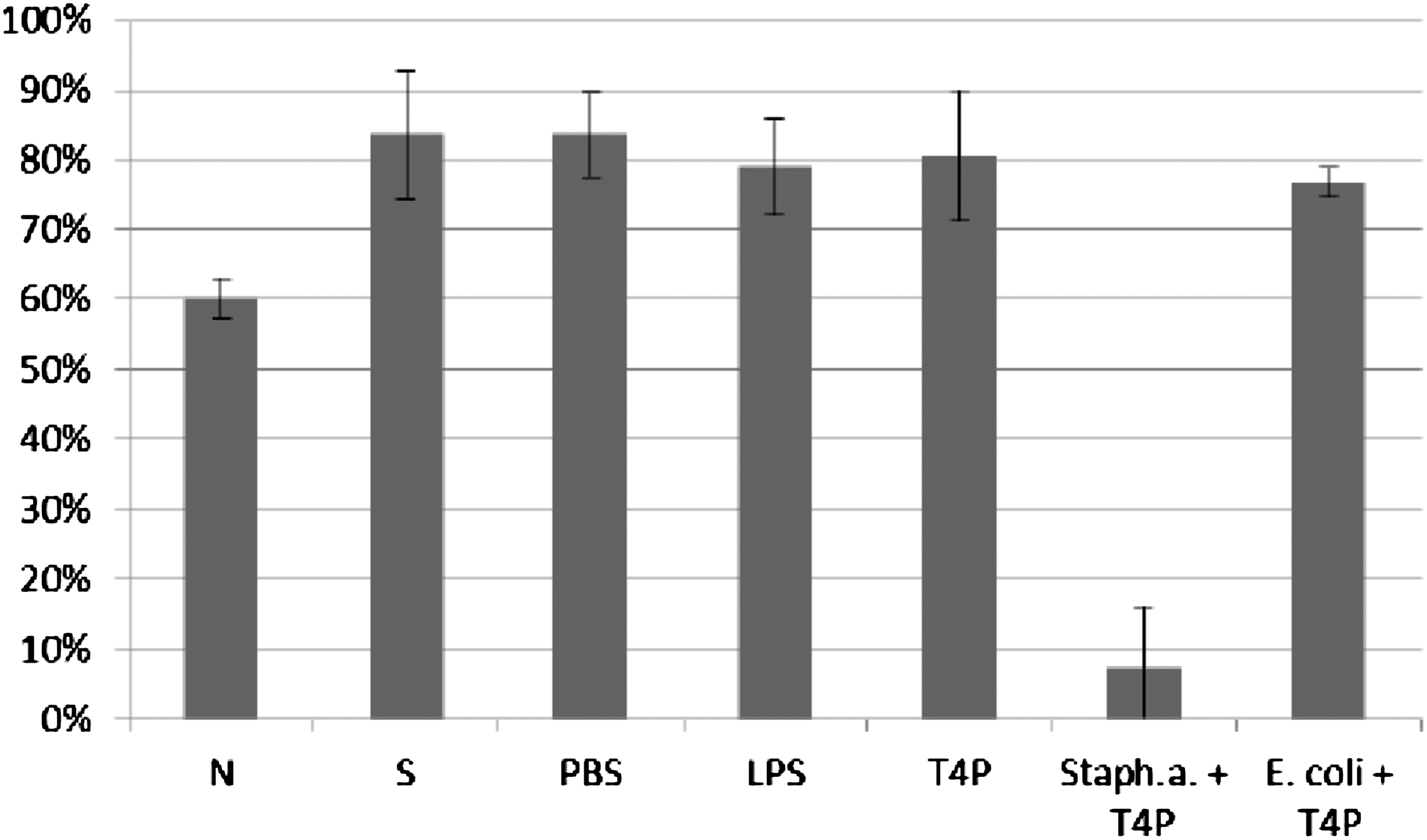
Influence of T4 purified preparation on ICK of bacteria Staphylococcus aureus by human granulocytes. Figure presents ICK of bacteria (%) by granulocytes after 1 h. Last two columns on the right present proportional lysis of bacteria E. coli and Staphylococcus aureus (%) by T4 purified preparation (no phagocytes in suspension) after 1 h.
In experiments in which the influence of phages heterologous to bacteria phagocytosed by granulocytes was studied, ICK (%) of bacteria by granulocytes was comparable to ICK of bacteria in the positive control (S), CM in the case of bacteriophage lysate, and PBS and LPS in the case of purified phage preparation. Preparations of phages heterologous to bacteria do not influence ICK of bacteria by human granulocytes.
Influence of phage preparations on ICK of bacteria by monocytes
Phages homologous to bacteria phagocytosed by monocytes (bacteria sensitive to phages)
Table 4 presents experimental models applied in studies of phage influence on ICK of bacteria by monocytes (phages homologous to bacteria).
ICK (%) of bacteria killed by monocytes was determined according to the procedure described in Material and Methods. ICK (%) of bacteria killed by monocytes incubated with phage lysate T4 (T4L) and purified preparation of phage T4 (T4P) is presented in Figures 7 and 8. Figures present the arithmetic mean from three experiments and standard deviation. Description of samples are as indicated in Material and Methods.
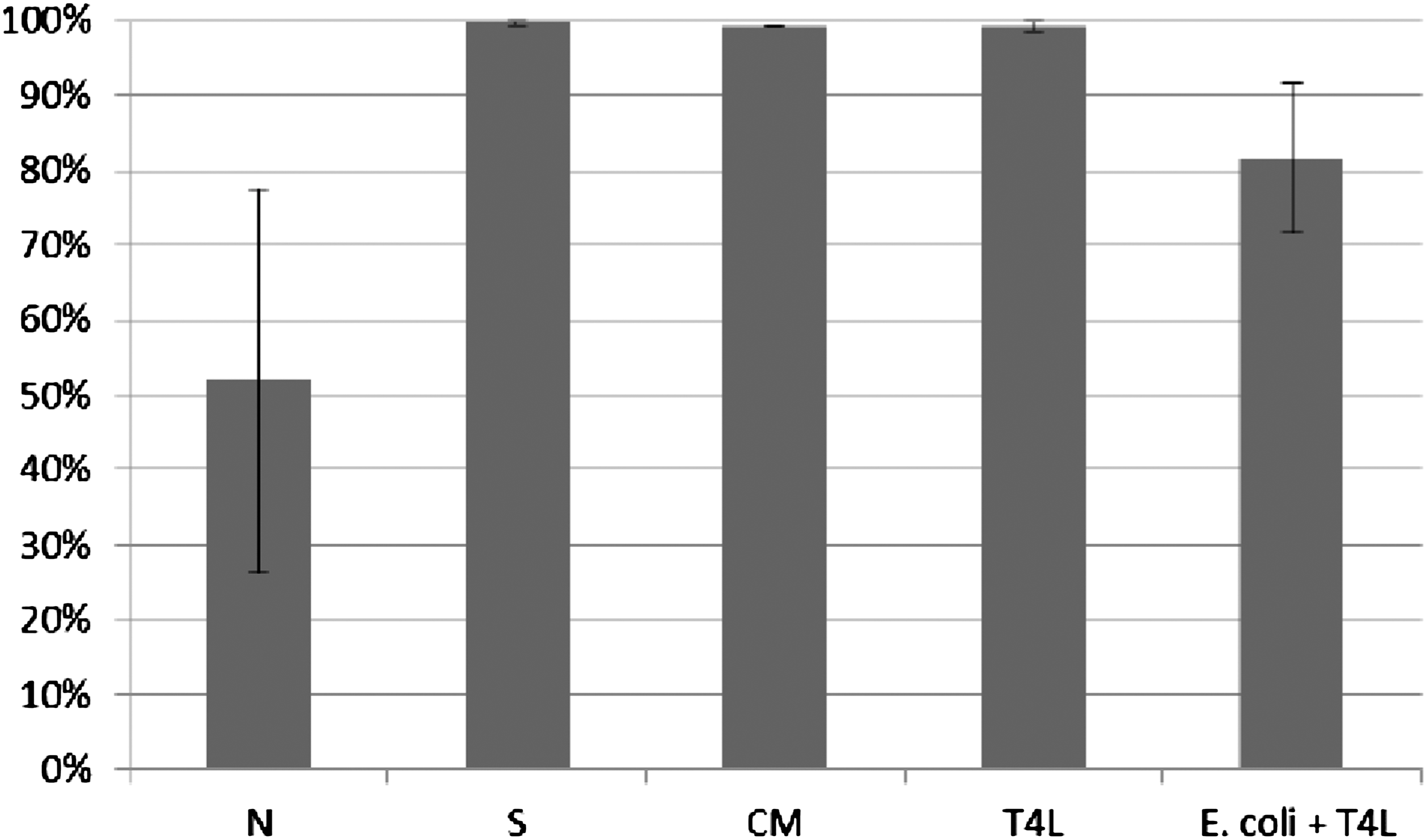
Influence of T4 lysate on ICK of bacteria E. coli by human monocytes. Figure presents ICK (%) of bacteria by monocytes after 1.5 h. Last column on the right presents proportional lysis of bacteria E. coli (%) by T4 lysate (no phagocytes in suspension) after 1.5 h.
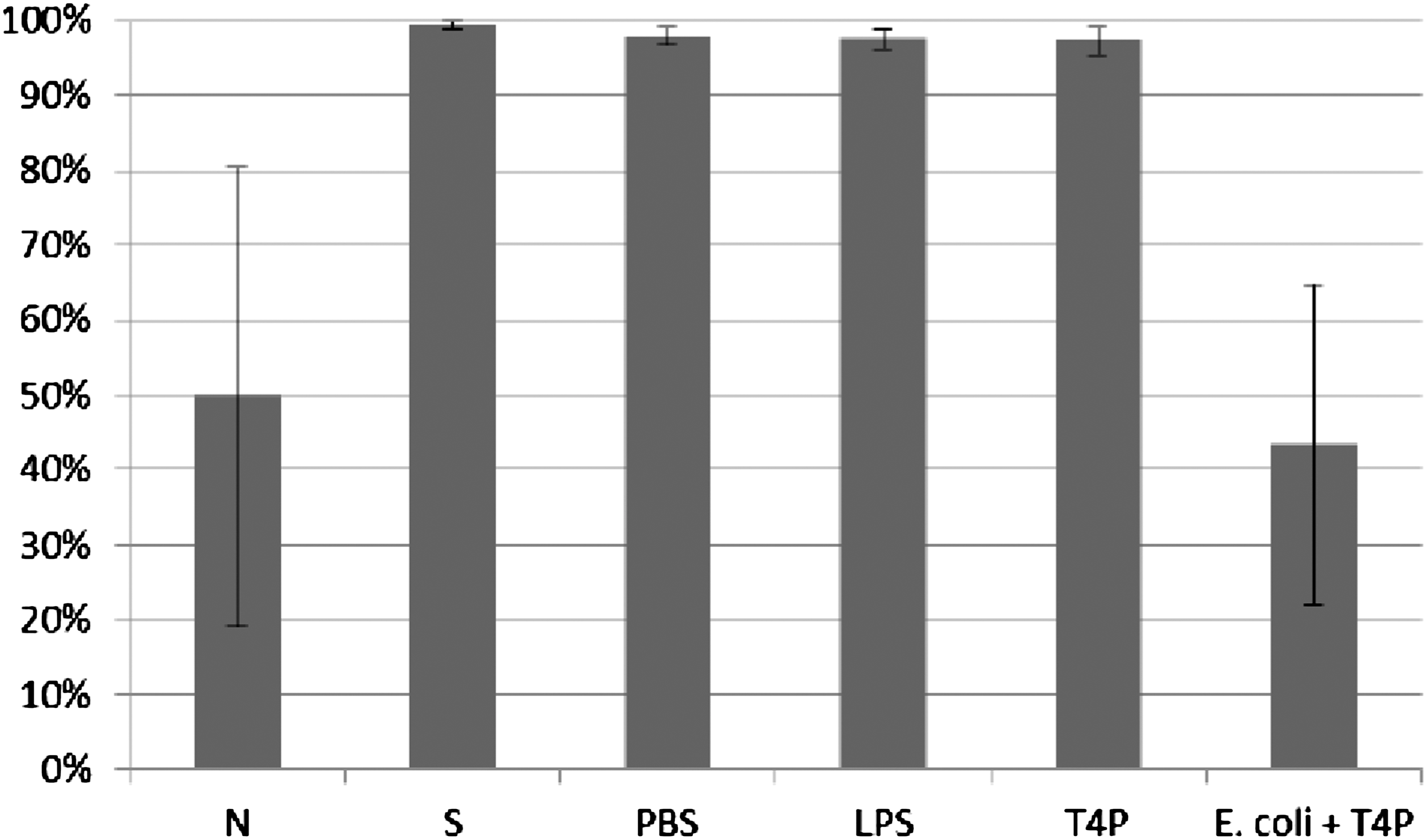
Influence of T4 purified preparation on ICK of bacteria E. coli by human monocytes. Figure presents ICK (%) of bacteria by monocytes after 1.5 h. Last right column presents proportional lysis of bacteria E. coli (%) by T4 purified preparation (no phagocytes in suspension) after 1.5 h.
In experiments in which the influence of phages homologous to bacteria phagocytosed by monocytes was studied, ICK (%) of bacteria by monocytes was comparable to ICK of bacteria in the positive control (S), CM in the case of bacteriophage lysate, and PBS and LPS in the case of purified phage preparation. Preparations of phages homologous to bacteria do not influence ICK of bacteria by human monocytes.
Phages heterologous to bacteria phagocytosed by monocytes (bacteria insensitive to phage)
Table 5 presents experimental models applied in studies on phage influence on ICK of bacteria by monocytes (phages heterologous to bacteria).
ICK (%) of bacteria killed by monocytes was determined according to the procedure described in Material and Methods. ICK (%) of bacteria killed by monocytes incubated with T4 phage lysate (T4L) and purified preparation of phage T4 (T4P) is presented on Figures 9 and 10. Figures present the arithmetic mean from three experiments and standard deviation. Descriptions of samples are as indicated in Material and Methods.
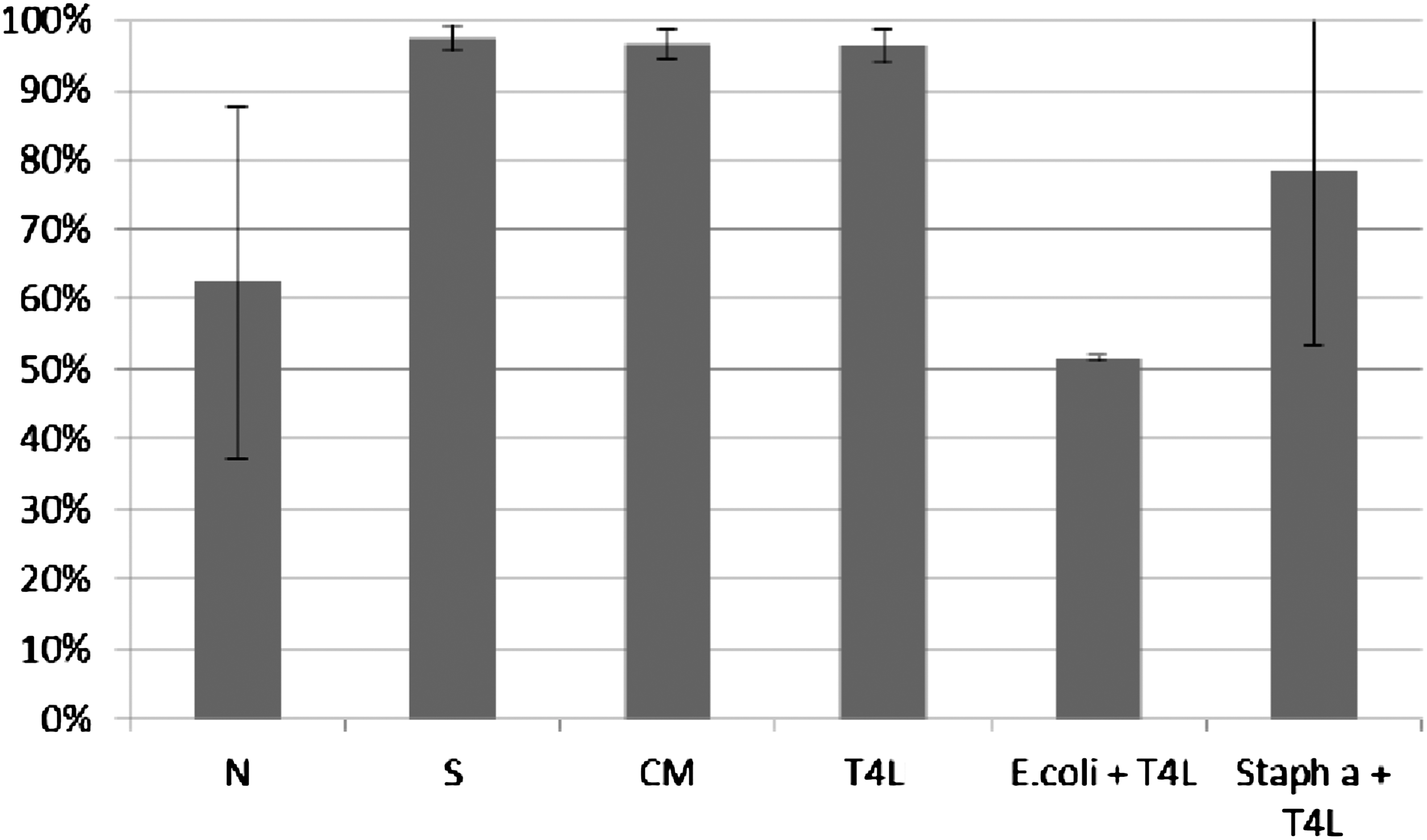
Influence of T4 lysate on ICK of bacteria Staphylococcus aureus by monocytes. Figure presents ICK (%) of bacteria by monocytes after 1.5 h. Last two columns on the right present proportional lysis of bacteria E. coli and Staphylococcus aureus (%) by T4 lysate (no phagocytes in suspension) after 1.5 h.
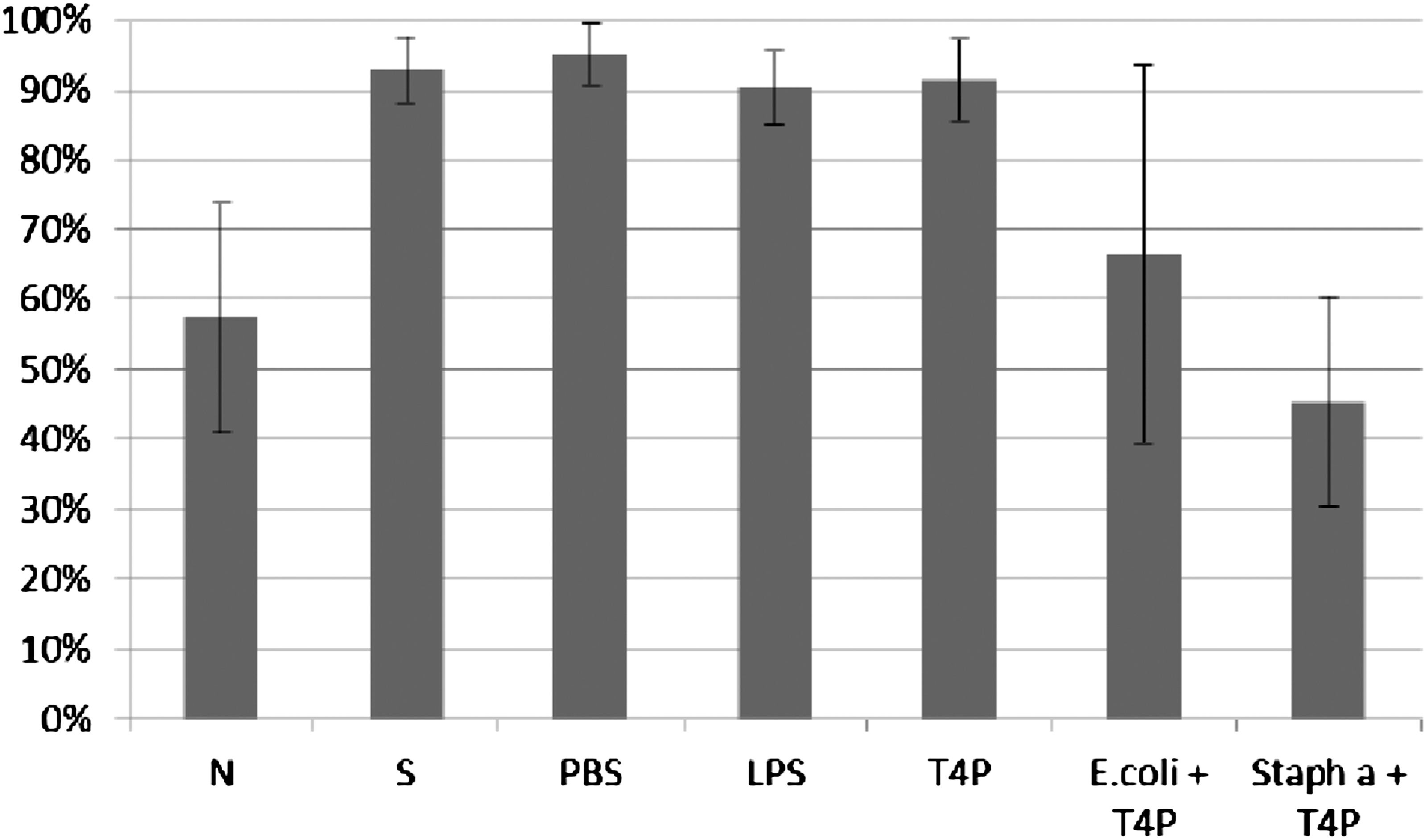
Influence of T4 purified preparation on ICK of bacteria Staphylococcus aureus by human monocytes. Figure presents ICK (%) of bacteria by monocytes after 1.5 h. Last two columns on the right present proportional lysis of bacteria E. coli and Staphylococcus aureus (%) by T4 purified preparation (no phagocytes in suspension) after 1.5 h.
In experiments in which the influence of phages heterologous to bacteria phagocytosed by monocytes was studied, ICK (%) of bacteria killed by monocytes was comparable to ICK of bacteria in the positive control (S), CM in the case of bacteriophage lysate, and PBS and LPS in the case of purified phage preparation. Preparations of phages heterologous to bacteria do not influence ICK of bacteria by human monocytes.
Influence of phage preparations with different phage titers on ICK of bacteria by human phagocytes
In the presented experiments, we used preparations with phage titer 1×109 pfu/mL because phage preparations with comparable phage titer are most commonly applied in phage therapy. The results of our studies showed that phage preparations with phage titer 1×109 pfu/mL do not influence ICK of bacteria by human granulocytes and monocytes; thus, we investigated whether any effect would be observed also for preparations with different phage titers. The influence of T4 lysates with phage titers 1×1010 pfu/mL, 1×108 pfu/mL, 1×106 pfu/mL, 1×104 pfu/mL on ICK of bacteria by human phagocytes was studied. Lysates of T4 phage did not influence ICK of bacteria by human phagocytes irrespectively of phage titer in the preparation.
Influence of ICK of clinical strains of bacteria by healthy donors
We did not observe any significant differences between mean ICK of E. coli B, as well as clinical strains, neither for granulocytes nor for monocytes in polymorphonuclear cells fraction isolated from healthy donors. The results are presented on Figures 11 and 12.
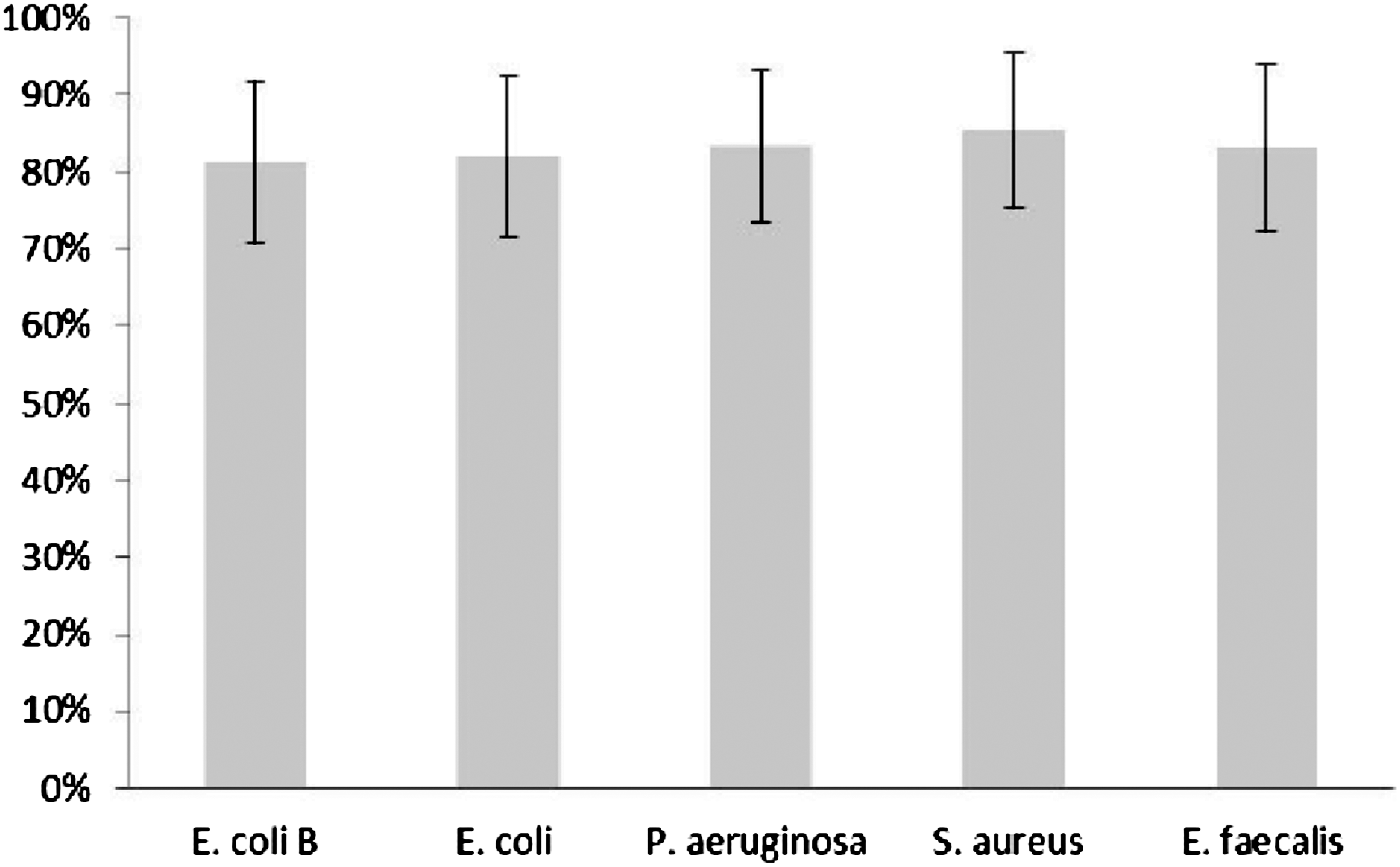
The mean intracellular killing of standard E. coli B strain and clinical bacterial strains by PMNs isolated from healthy donors` blood. Bars represent±SE. No significant differences between studied groups were observed.
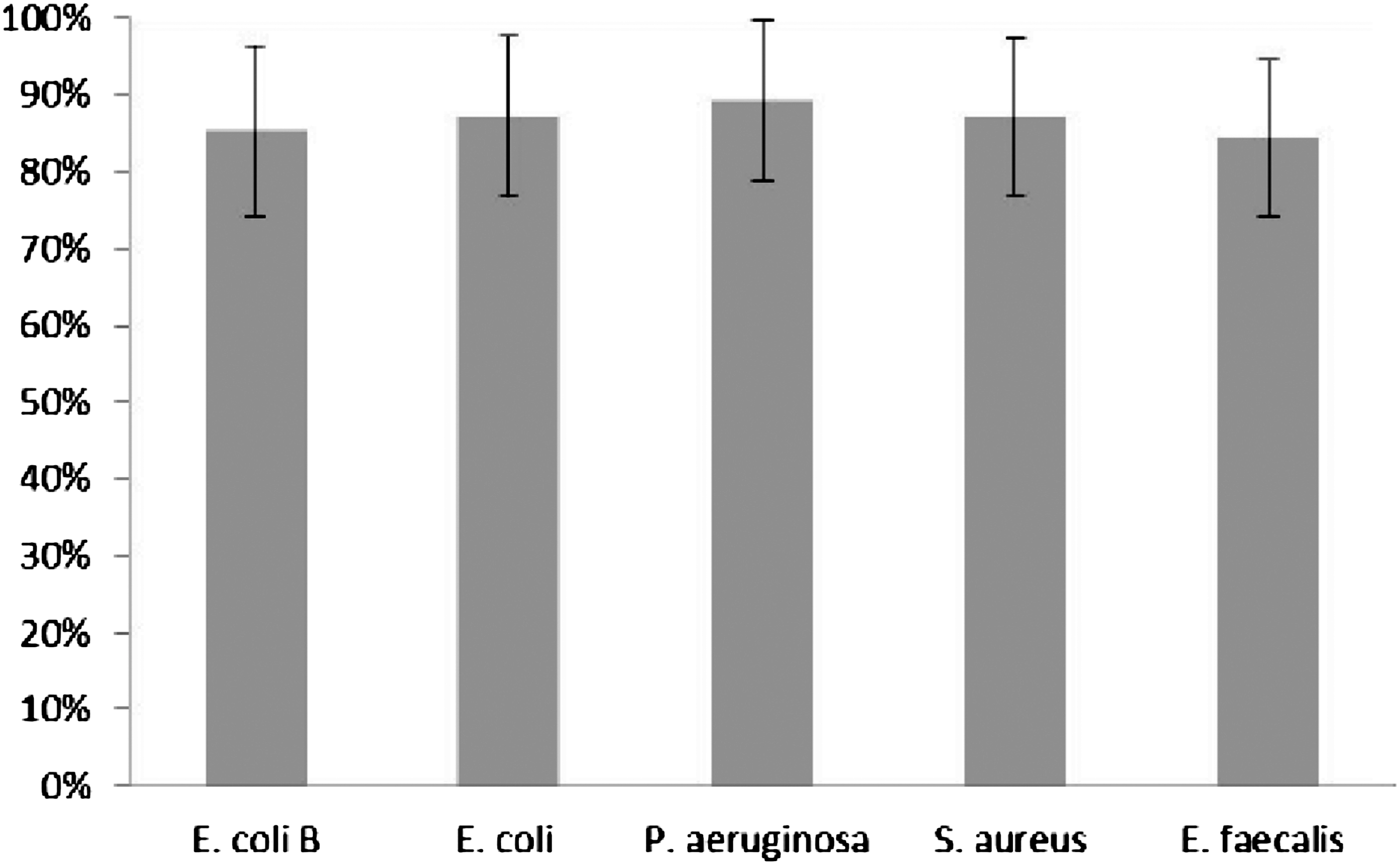
The mean intracellular killing of standard E. coli B strain and clinical bacterial strains by PBMCs isolated from healthy donors` blood. Bars represent±SE. No significant differences between studied groups were observed.
Discussion
In our study, we determined the influence of selected bacteriophage lysates and selected purified phage preparations on ICK of bacteria by human granulocytes and monocytes. We also examined whether the influence of the phage preparation is conditional upon phage titer.
Our study concerned interactions between phages and phagocytes, cells playing a crucial role in organisms' defense mechanisms. Killing abilities are one of the most important features of phagocytes determining their effective struggle with pathogens. Dysfunctions of bactericidal abilities of phagocytes may entail illness such as chronic infections (30). We found that phage preparations do not influence ICK of bacteria by human granulocytes and monocytes. The observed effect is irrespective of phage preparation type (lysate, purified phage preparation), phage titer in the preparation (phage titer range: 1×104 pfu/mL–1010 pfu/mL) and experimental model (bacteria sensitive or insensitive to phages).
Our previous studies showed that most phage preparations do not influence migration of human granulocytes and mononuclear cells (Kurzepa-Skaradzinska et al., data not shown), which might be in line with the presented data indicating that bacteriophages have no impact on phagocyte activity.
The results of our study have obvious consequences for phage therapy, during which bacteria are targets of both bacteriophages and phagocytes, so their mutual interactions are of great importance. Our study was significant in context of the fact that patients subjected to phage therapy often have phagocyte dysfunctions. It concerns almost 50% of patients who come to the Phage Therapy Unit at IIET, PAS) (23, 24). Those dysfunctions are frequently the consequence of long-term antibiotic therapy (31 –34), but they also may result from mechanisms protecting bacteria from elimination (35 –39). Phagocyte dysfunctions may also result from basic illness. In all these cases, combating bacterial infections with antibiotics is hampered; thus bacteriophage therapy is a vital method of treatment. Determining whether phage preparations have any impact on phagocyte activity is thus of great practical value.
It was found that antibiotics may have a negative influence on activity of immune cells (40 –46). Tetracycline inhibits phagocytosis and ICK of bacteria by leukocytes, and also decreases their migration rate at the site of infection. Sulfonamides affect bactericidal activity of these cells. In experiments on mice, it was observed that antibiotics may affect intestinal innate defense mechanisms (47). Lack of a negative influence of phage therapy on phagocyte activity might indicate its safety in comparison with at least some antibiotics.
Our results have also great practical value for patients with impaired defense mechanisms and increased susceptibility to bacterial infections as a consequence of taking immunosuppressive drugs (e.g., allotransplant recipients). It was found that neutrophils of these patients demonstrate dysfunctions of bacterial killing (48), which may result from the direct impact of immunosuppressive drugs on those cells (49). Our results indicating that phage preparations do not impair ICK of bacteria by granulocytes and monocytes support the view that phage therapy does not present an additional risk for patients with defense dysfunctions secondary to immunosuppressive treatment.
We would like to emphasize the original character of our study. We have not found any reports in the literature regarding the influence of phage preparations on ICK of bacteria by human phagocytes. We would also like to emphasize that our study is preliminary; further studies are required, particularly studies in vivo, determining the direct influence of phage therapy on activity of phagocytes of patients subjected to phage therapy. It is fully reasonable considering the results of our previous studies showing that phage therapy may support renewal of granulocytes and decrease their phagocytic activity (19).
In our study, we took up the challenge to extend the knowledge regarding phage–phagocyte interactions. We believe that the results of our study contribute valuable knowledge concerning biological activity of bacteriophages and their influence on the human organism, contributing to optimization of phage therapy and allowing identification of new research directions.
Footnotes
Acknowledgments
Experiments on the ICK of clinical strains by phagocytes isolated from healthy donors` blood samples are part Ewa Jończyk-Matysiak's doctoral study. They were financially supported by the European Regional Development Fund within the Operational Program Innovative Economy, 2007–2013, Priority axis 1. Research and Development of Modern Technologies, Measure 1.3 Support for R&D projects for entrepreneurs carried out by scientific entities, Submeasure 1.3.1 Development projects as project No. POIG 01.03.01-02-003/08 entitled “Optimization of the production characterization of bacteriophage preparations for therapeutic use”.
Author Disclosure Statement
No competing financial interests exist.