
Abstract
DNA vaccine can induce the prolonged immune responses against the encoded antigen with the appropriate adjuvant. To study the immunogenicity of the HIV-1 vif DNA vaccine in inducing the humoral and cellular immune responses and the immunoadjuvant effect of LIGHT, which is a member of TNF superfamily and can stimulate the proliferation of naïve T cells as a co-stimulatory molecule, DNA vaccine plasmid pcDNA-Vif was constructed by inserting HIV-1 vif gene into the downstream of CMV promoter in eukaryotic expression vector pcDNA3.1(+). In vitro expression of HIV-1 Vif in pcDNA-Vif-transfected HeLa cells was confirmed in transcriptional and protein level by RT-PCR and Western blot, respectively. After BALB/c mice were injected muscularly with DNA vaccines for three times, the specific immune responses were analyzed. The data showed that anti-Vif antibody response, Vif-specific T cell proliferation, and CTL activities were induced in the mice that were inoculated with HIV-1 vif DNA vaccine plasmid. Interestingly, stronger humoral and cellular immune responses were detected in mice that were immunized with plasmid pcDNA-Vif and pcDNA-LIGHT together compared to the single immunization with plasmid pcDNA-Vif alone. Together, the results of the study suggest that candidate HIV-1 DNA vaccine can elicit HIV-1 Vif-specific immune responses in mice and that LIGHT plays the role of immunoadjuvant in co-immunization with DNA vaccine.
Introduction
DNA vaccine is a promising technology to elicit both humoral and cellular immune responses in humans for controlling HIV-1 infection (3,17). There are many advantages of DNA vaccine over other vaccines such as subunit vaccine, and live recombinant vector vaccine for prophylactic or therapeutic purpose. First, DNA vaccine can induce prolonged humoral and cellular responses. Second, DNA vaccine is safe because only naked DNA is immunized but not infectious agents. Last, it is cheap and easy to produce. Various routes for DNA vaccine delivery have been used including intramuscular, intradermal, intravenous, intranasal, and epidermal injections (6). The types of immunity elicited by DNA vaccine are influenced by the routes and doses of DNA inoculation routes. For example, intramuscular and intradermal injections induce T helper 1 (Th1) responses characterized by increased production of interferon-γ (IFN-γ) and antigen-specific IgG2a antibody. In contrast, epidermal inoculation elicits T helper 2 (Th2) responses characterized by increased production of interleukin (IL)-4 and IgG1 antibody (5,23).
Since the emergence of gene-based immunization methodologies, various strategies have been used to increase the efficiency of DNA vaccination. In several studies, the co-inoculation of plasmids encoding cytokine or co-stimulatory molecule genes as adjuvants significantly enhanced the immune response elicited by DNA vaccines (2,13,19). LIGHT, which is homologous to lymphotoxins, exhibits inducible expression, and competes with HSV glycoprotein D for herpes virus entry mediator (HVEM), a receptor expressed by T lymphocytes, also named HVEM-L or TNFSF14, is a member of the TNF superfamily (18). LIGHT can induce apoptosis of tumor cells expressing both LTβR and TR2/HVEM receptors (30). LIGHT also can function as a co-stimulatory molecule for human naïve T cells to proliferate (27,31). The mixed lymphocyte reaction (MLR) can be enhanced by inclusion of soluble LIGHT, and MLR can be inhibited by neutralization of LIGHT (9,14). Therefore, LIGHT can be used as a potential immunoadjuvant in DNA vaccination.
In the present study, the immunogenicity of HIV-1 vif DNA vaccine and the immunoadjuvant effect of LIGHT were investigated. The immune responses to Vif were analyzed in mice immunized with DNA vaccine. Specific immune responses to HIV-1 Vif can be elicited by plasmid DNA. Humoral and cellular immune responses elicited by the co-immunization of pcDNA-Vif and pcDNA-LIGHT are stronger than those induced by pcDNA-Vif alone.
Materials and Methods
Plasmid construction
The eukaryotic expression vector pcDNA3.1(+), purchased from Invitrogen Corporation (San Diego, CA), was used as the parental plasmid for constructing the expression plasmids. Plasmid pET32a(+)-Vif harboring the full-length of HIV-1 vif gene was provided by the author's lab. The HIV-1 vif gene was extracted by digesting plasmid pET32a(+)-Vif with EcoR I/Xho I and subcloned into pcDNA3.1(+) predigested with EcoR I/Xho I(TaKaRa, China) and the plasmid was designated as pcDNA-Vif. Plasmid pcDNA-LIGHT was constructed previously (31). Briefly, the PCR products of LIGHT gene from bone marrow (BM)-derived immature DCs were cleaved with restriction endonuclease EcoR I/Xho I(TaKaRa, China) and were cloned into the restriction sites of pcDNA3.1(+) digested with the same endonucleases. Plasmid pcDNA-EGFP expressing enhanced green fluorescent protein was used to detect the transfection efficiency and was provided by the author's lab.
Antibodies and peptides
Anti-HIV-1 Vif polyclonal antibodies were prepared by immunizing the mice with the purified Vif protein expressed in E.coli (data not shown). Briefly, the prokaryotic expression vector pET32a(+)-Vif was constructed and transformed into E. coli BL21(DE3), the recombinant HIV-1 Vif protein was expressed in E. coli BL21(DE3) under the induction of IPTG at 37°C, and the polyclonal antibodies against HIV-1 Vif were prepared by immunization of Balb/c mice with recombinant HIV-1 Vif protein purified from polyacrylamide gels. Goat anti-mouse IgG-HRP antibodies were obtained from Tiangen Co. Ltd. (Beijing, China). The HIV-1 Vif epitope peptide (WKRLVKHHMYISRKA,25–39) and HIV-1 Nef epitope peptide (GVRYPLTFGWCYKLVP,132–147) were synthesized by GL Biochem Co. Ltd. (Shanghai, China).
Cell line and plasmid transfection
HeLa cells (human cervical epithelial carcinoma cell line), purchased from Shanghai Institute of Biochemistry and Cell Biology of Chinese Academy of Sciences, were cultured in RPMI 1640 supplemented with 10% FBS (Biowest, China). Cells were seeded in a 6-well plate at 3×105 cells and the transfection was performed until the cells were 80% confluent at 37°C in a humidified 5% CO2 air incubator. The DNA–liposome complexes which contained 4 μg of DNA and 6 μg of lipofectamine (Beyotime Biotech, China) in a total 70 μL of serum-free RPMI 1640 medium were incubated for 15 min at room temperature and added to the cells. After incubation for 4 h with serum-free RPMI 1640 medium, the cells were supplemented with 10% FBS for the following 24 h.
Extraction of total RNA and RT-PCR
The total RNA was isolated from the transfected cells by TRIzol (Invitrogen, USA) and cDNA was synthesized using oligo(dT)20 primer, dNTP mixture, and reverse transcriptase ReverTra Ace (Toyobo, Japan). The vif gene was amplified by PCR using the forward (5’-GGAATTCATGGAAAACAGA TGGCAGGTG-3’) and reverse (5’-CCGCTCGAGGTGTCCA TTCATTGTAT-3’) primers. The PCR program was as follows: 95°C for 2 min, 32 cycles of 94°C for 1 min, 64°C for 1 min, 72°C for 1 min and 72°C for 10 min. The PCR products were identified by 1% agarose gel electrophoresis.
Western blot analysis
The total protein was extracted by lysis buffer (10 mM Tris-Cl, pH 7.4, 1 mM MgCl2, 0.5% NP40, 20 μg/mL DNase I) and then separated on 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). After transferring the protein bands to a nitrocellulose membrane (NC), the membrane was blocked with 5% (w/v) nonfat milk for 1.5 h at room temperature and then incubated with 5000-fold dilution of anti-HIV-1 Vif polyclonal antibodies as the primary antibody overnight at 4°C. After being washed three times with TBST (Tris-buffered saline with 0.1% Tween 20), the membrane was incubated with the HRP-conjugated goat anti-mouse antibodies (Tiangen Biotech, China) for 1 h at room temperature. The NC membrane was then stained with enhanced chemiluminescent substrate (Thermo Scientific, Shanghai) and exposed to x-ray film.
Immunization of mice
Female BALB/c (H-2Kd) mice, 6–8 weeks old, were purchased from the Experimental Animal Center of the Second Military Medical University (Shanghai, China) and raised according to the institutional guidelines. The mice were divided into five groups with six mice in each group and were intramuscularly immunized in both left and right quadriceps muscles for three times at 3-week intervals with one of the following plasmids dissolved in 100 μL of PBS: 1) 100 μg of pcDNA-Vif; 2) a mixture of 50 μg of pcDNA-Vif and 50 μg of pcDNA-LIGHT; 3) 100 μg of pcDNA-LIGHT; 4) 100 μg of pcDNA3.1 vector; 5) 100 μL PBS. Half of the dose was given in each leg.
Measurement of anti-HIV-1 Vif antibodies
Sera were collected by tail vein bleeding at day 0 and every 2 weeks after the first immunization and by eye socket bleeding when the mice were sacrificed 2 weeks after the final immunization. The anti-HIV-1 Vif specific antibodies were detected by enzyme-linked immunosorbent assay (ELISA). Each well of the ELISA plate was coated with recombinant HIV-1 Vif protein expressed in E. coli (data not shown) overnight at 4°C, and the diluted sera of the indicated time points were then added to the wells. The horseradish peroxidase-conjugated goat anti-mouse IgG was used to detect the bound immunoglobulins. After the plates were washed and reacted with an OPD solution and stopped with H2SO4, the optical density (OD) was measured at 450 nm with ELISA plate reader.
Splenocyte proliferation assay
Two weeks after the final immunization, the splenocytes were prepared through sterile stainless steel mesh and washed twice with RPMI 1640. The erythrocytes were removed by NH4Cl solution. After being washed twice with RPMI 1640, the splenocytes were re-suspended in RPMI 1640 medium supplemented with 10% FBS. Cell numbers were counted with a hemocytometer by trypan blue dye and the cell viability exceeded 95%. Splenocytes were grown in triplicate in 96-well flat-bottom microtiter plates (2×105 cells per 100 μL RPMI 1640 medium). 10 μg/mL of HIV-1 Vif peptide was added to the wells, while HIV-1 Nef peptide was served as irrelevant peptide control. The equal volume of the medium and ConA (10 μg/mL) were used as the blank control and positive control, respectively. After the splenocytes were incubated at 37°C in a CO2 incubator for 72 h, 10 μL of MTT solution (5 mg/mL) were added to each well and incubated for another 4 h. Then 100 μL of formazan solvent was added to each well and incubated until formazan was solved. The absorbance was detected in an ELISA plate reader at 570 nm and the results were expressed as a stimulation index (SI, SI=OD570 of peptide stimulation/OD570 of blank control), which indicates significant proliferation if SI>2.
Cytotoxic T lymphocyte assay
The cytotoxic T lymphocyte (CTL) assay was performed as previously described (11). The splenocytes were prepared and cultured in a complete medium with 10 U/mL IL-2 and 5 μg/mL ConA at 37°C in a humid atmosphere with 5% CO2 for 1 or 2 days as the effector cells. The splenocytes were harvested from naïve mice and were pulsed with final concentration of 20 μg/mL of HIV-1 Vif peptide for another 4 h and then treated with 80 μg/mL of Mitomycin C (Genview, USA) for 2 h. The cells were washed four times with RPMI 1640 and used as the stimulator cells. The effector cells (4×107) were incubated with stimulator cells at an effector-stimulator ratio of 10:1 for 4 days at 37°C in 5% CO2. P815 cells (mouse mastocytoma cell line, Shanghai Institute of Biochemistry and Cell Biology of Chinese Academy of Sciences) were prepared as the target cells after pulsed with HIV-1 Vif peptide for 4 h at 37°C in 5% CO2 and treated for 2 h with 80 μg/mL of Mitomycin C. The ability of effector cells to kill P815 target cells was detected by a Nonradioactive Cytotoxicity Assay Kit (Promega, USA). The ratio of the effector and target cells was 100:1, 50:1, 25:1, and 12.5:1 in triplicate, respectively. The supernatant was collected and measured according to the instructions. The percentage of specific lysis of P815 target cells for a given effector cell sample was calculated as follows: specific lysis=(OD of experimental LDH release – OD of effector cell spontaneous LDH release − OD of target cell spontaneous LDH release)/(OD of maximum target cell LDH release − OD of target cell spontaneous LDH release) x 100%.
Statistical analysis
Data were reported as mean±SD and were analyzed by professional statistical software SPSS. Significance was set at a value of p<0.05.
Results
In vitro expression of HIV-1 Vif DNA vaccine plasmid
Vif gene was digested from pET32a(+)-Vif with EcoR I/Xho I and cloned into pcDNA3.1(+) predigested with Eco R I/Xho I. Then the insertion of vif was identified by the enzyme digestion of the recombinant plasmid with restriction endonuclease and gene sequencing (data not shown). The transfection efficiency was confirmed by transfecting plasmid pcDNA-EGFP into HeLa cells and GFP-positive cells were visualized under a fluorescent microscope (data not shown).
The transcription of HIV-1 vif gene in pcDNA-Vif-transfected HeLa cells was detected by RT-PCR (Fig. 1A). The extracted RNAs were used to synthesize the cDNA, and PCR was performed with the specific primers of HIV-1 vif gene. A specific DNA band between 500 and 750 bp was shown in pcDNA-Vif-transfected cells, while there were no specific DNA bands detected in vector plasmid-transfected cells. In order to exclude the contamination of plasmid DNA of pcDNA-Vif in the transfected cells, the extracted total RNAs were treated with DNase (TaKaRa, China) according to the manufacturer's protocol. The expression of HIV-1 Vif in protein level was detected by Western blot, and the data are shown in Figure 1B. A specific protein band between 20 and 26 kDa was shown only in pcDNA-Vif-transfected cells but not in vector plasmid-transfected cells when anti-HIV-1 Vif antibodies were used.
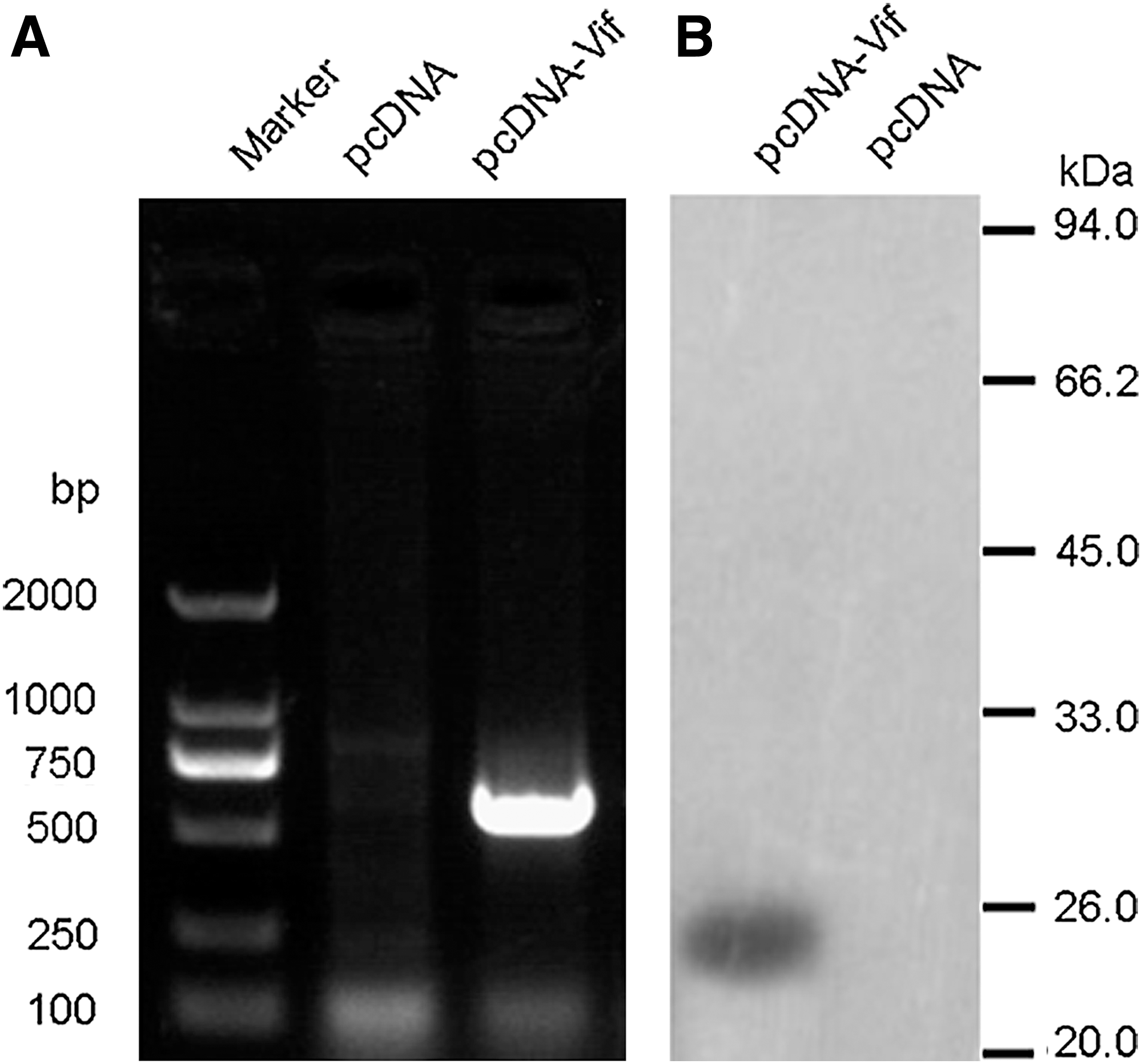
In vitro expression of HIV-1 Vif.
Measurement of HIV-1 Vif-specific antibody
The serum antibodies were detected by ELISA in 96-well polystyrene microtiter plates coated with recombinant HIV-1 Vif protein. The kinetics of antibody production are shown in Figure 2A.The result showed that anti-HIV-1 Vif antibodies were detectable at week 2 and the antibody titers increased following the booster immunization both in single immunization mice with pcDNA-Vif alone and in co-immunization mice with pcDNA-Vif plus pcDNA-LIGHT. Interestingly, the antibody titers in co-immunization mice were significantly higher than those in single immunization mice from week 4 (p<0.01). The sera collected from the sacrificed mice were also detected with ELISA after 1000-fold dilution; the data are shown in the OD value at 450 nm (Fig. 2B). Anti-HIV-1 Vif antibodies in the group immunized with HIV-1 vif DNA vaccine plasmid were significantly higher than the control group (p<0.01), and the mice immunized with pcDNA-Vif plus pcDNA-LIGHT had higher antibody level than the mice immunized with pcDNA-Vif alone (p<0.01).
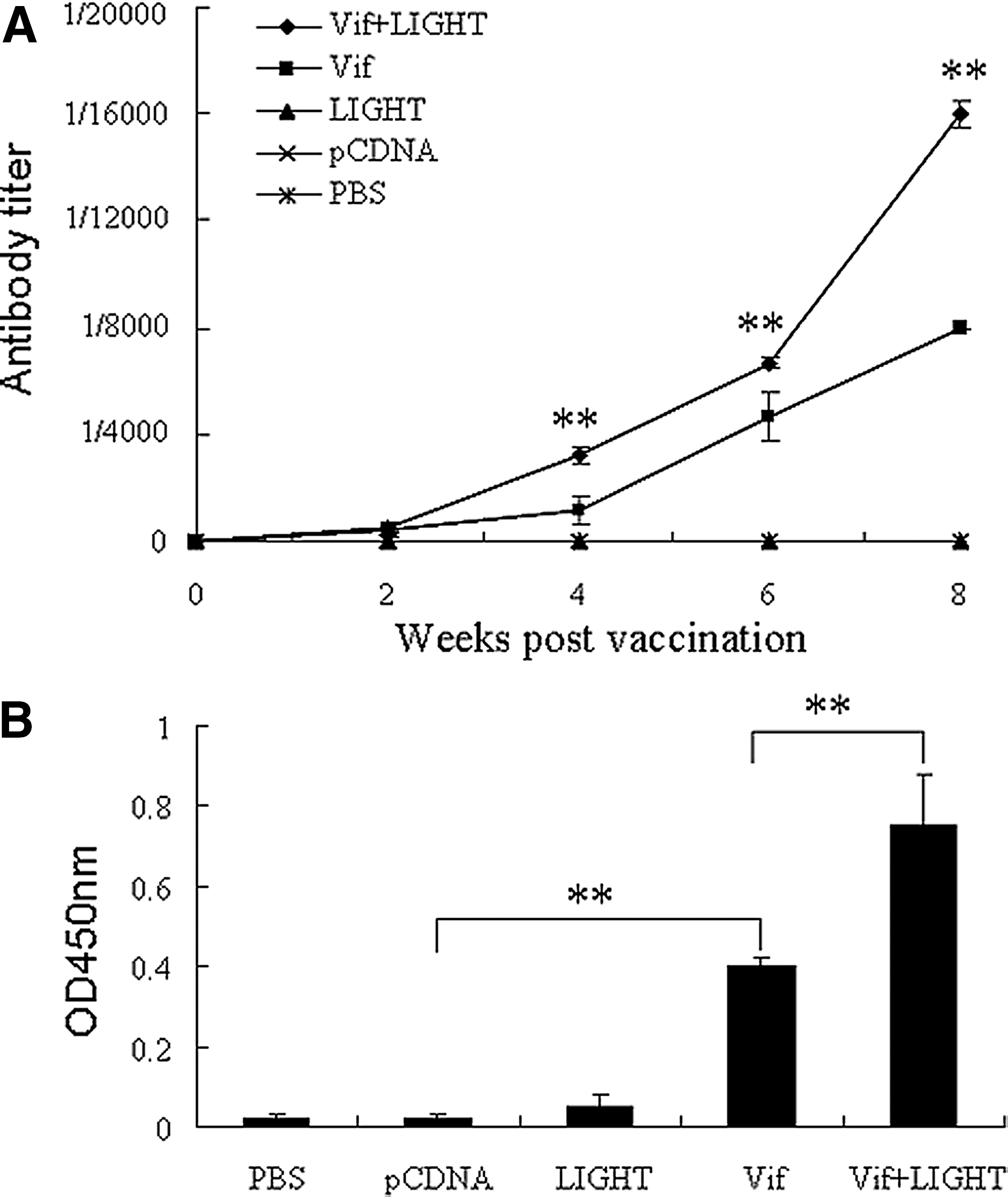
ELISA analysis of anti-HIV-1 Vif antibodies. 6–8 week old female BALB/c mice were administered i.m. with HIV-1 vif DNA vaccine or co-immunized with LIGHT gene, respectively. Mice inoculated with pcDNALIGHT, vector plamid pcDNA3.1(+), or PBS were used as controls. Sera from mice were collected at 2 week intervals and detected by ELISA. The results were expressed as the highest dilution while the ratio (OD450 of experimental group/OD450 of negative control) was more than 2
HIV-1 Vif-specific T cell responses
To determine whether HIV-1 vif DNA vaccine could elicit Vif-specific T cell responses and whether LIGHT played the costimulatory effects on DNA vaccine, a splenocyte proliferation assay was performed by MTT method. As shown in Figure 3, significant splenocyte proliferation was detected in the single immunization group with pcDNA-Vif as compared with the control groups (p<0.01). Higher SI was shown in the co-immunization group with pcDNA-Vif plus pcDNA-LIGHT (p<0.05). No significant proliferation was observed in the negative control in the presence of irrelevant peptide in all the groups.
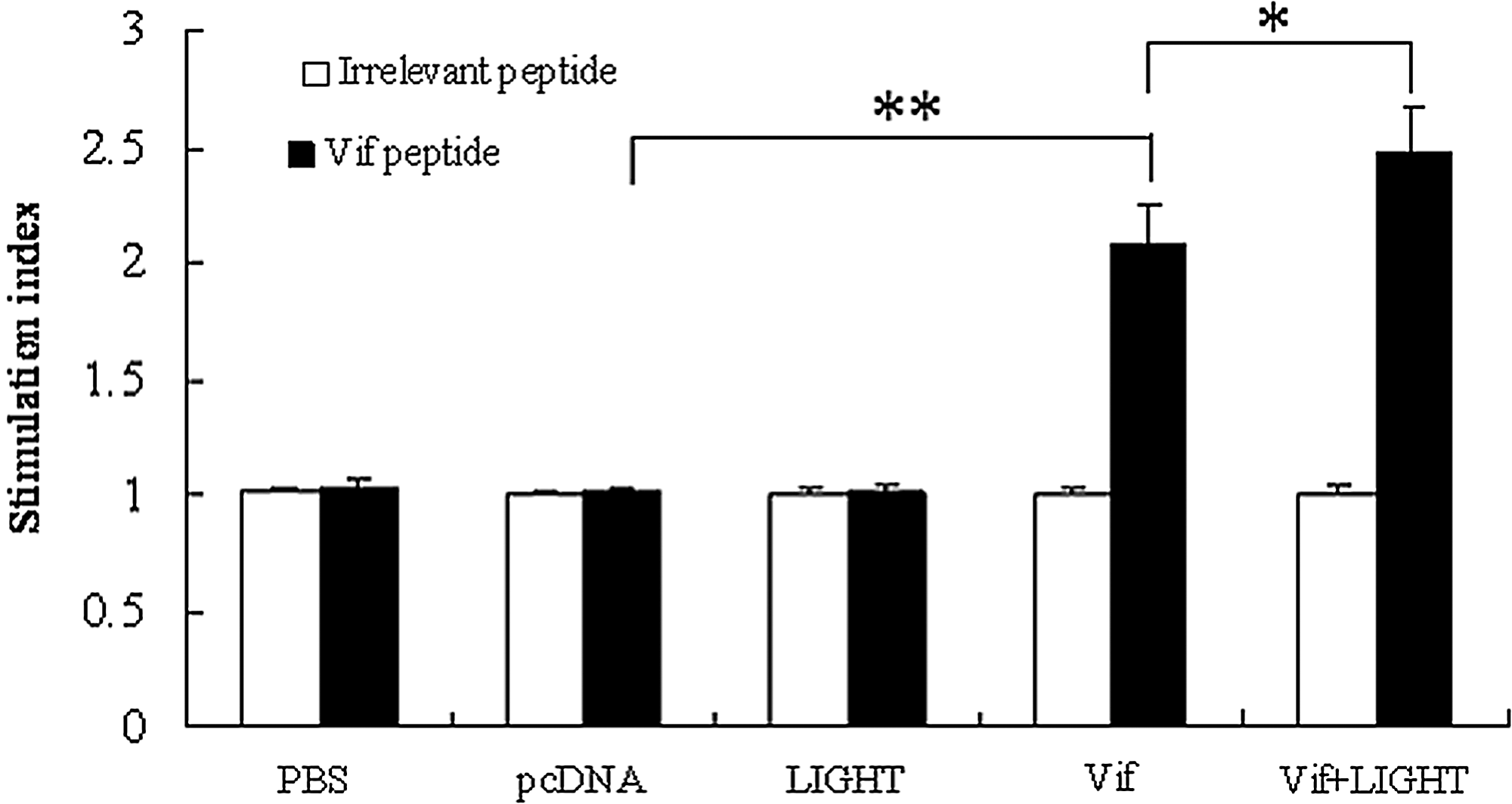
Splenocyte proliferation assay. Splenocytes were obtained from the mice 2 weeks after the last immunization and pulsed with Vif peptide (10 μg/mL) or irrelevant peptide (10 μg/mL) for another 4 days before the MTT assay was performed. The data are the mean±SD from six mice; *p<0.05; **p<0.01.
Induction of CTL responses in mice immunized with HIV-1 DNA vaccine
Splenocytes derived from the immunized mice 2 weeks after the final immunization were re-stimulated by the naïve splenocytes pulsed with HIV-1 Vif peptide for 4 days. P815 cells pulsed with HIV-1 Vif peptide were used as target cells. The specific lysis was detected by nonradioactive LDH release assay; the data is shown in Figure 4. HIV-1 Vif-specific CTL activity was detectable in the group immunized with HIV-1 DNA vaccine and the group co-immunized with HIV-1 DNA vaccine and LIGHT plasmid as compared with the control groups. The co-immunization group could induce stronger CTL responses than the group injected with HIV-1 DNA vaccine alone, which indicated that co-delivery of LIGHT could enhance the HIV-1 Vif-specific CTL response.
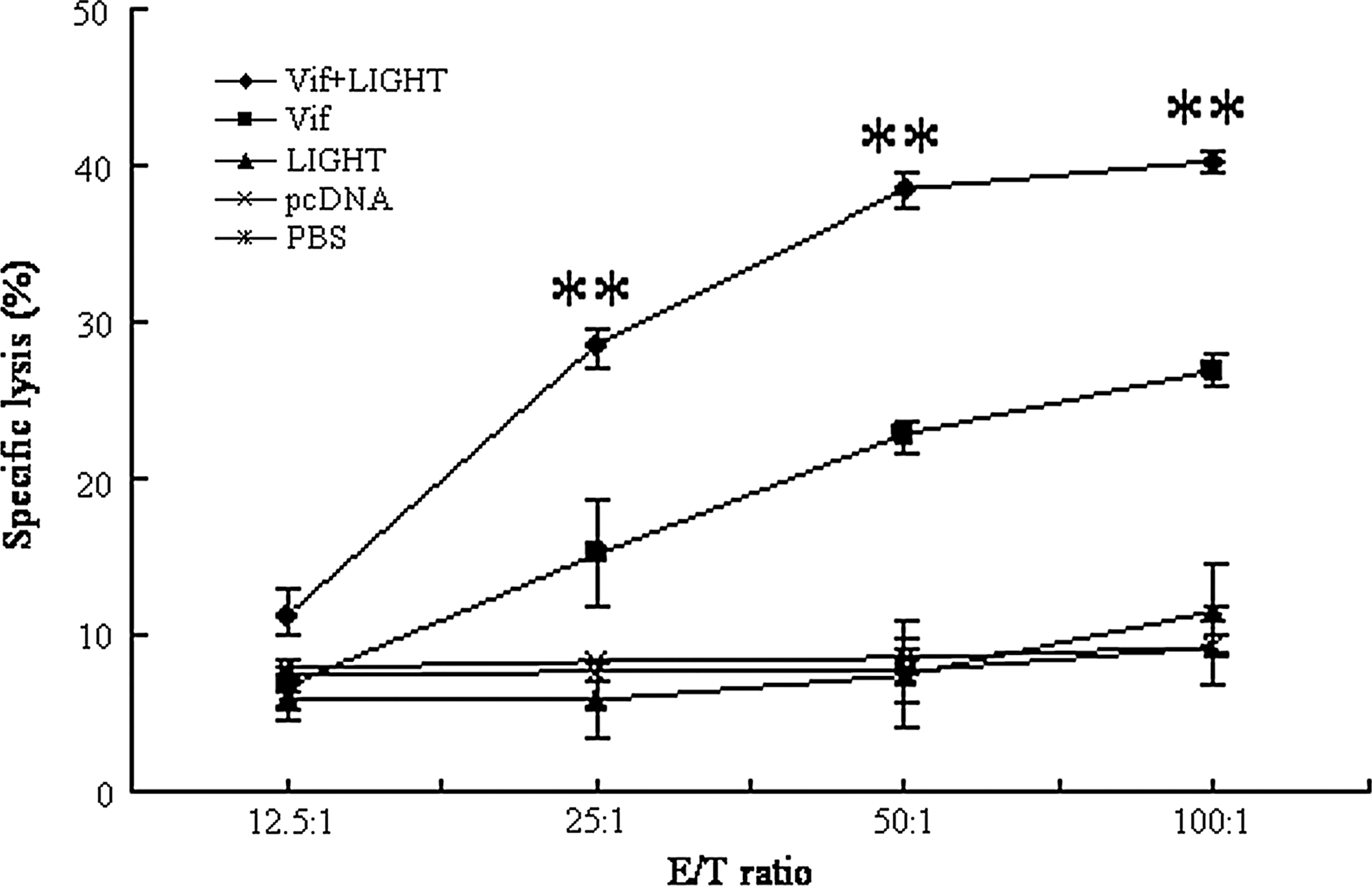
CTL activity. BALB/c mice were immunized 3 times with HIV-1 Vif DNA vaccine or co-injected with LIGHT gene adjuvant every 3 weeks. The parental plasmid pcDNA3.1(+), pcDNA-LIGHT, or PBS was inoculated as the controls. Splenocytes from the mice were prepared as the effector cells and the specific lysis on target cells was performed. The percentage of specific lysis is reported on the vertical axis. The data are the mean±SD from six mice; **p<0.01.
Discussion
The AIDS epidemic has always been a health threat since the first description in 1981 (20). There have been several candidate HIV-1 vaccines which elicited varying degrees of protective responses in nonhuman primate models such as DNA vaccines, subunit vaccines, live vectored recombinant vaccines, and various prime-boost combinations (7). DNA immunization could stimulate both humoral and cellular immune responses in animal models, and the potential utility of plasmid DNA as a component of an AIDS vaccine is currently an area of active investigation (21,29). HIV-1 Vif protein is a 23-kDa accessory protein encoded by 609-nucleotides and critical for virus infectivity because of the capacity to counteract with the APOBEC3G (10). There is extensive amino acid conservation in the Vif sequences and Vif mutants leading to 25% reduction in the infectivity (25,28). CTL responses against Vif may be effective for virus suppression and the Vif epitopes may be important for the vaccine-induced immunity. In the current study, DNA vaccine plasmid containing HIV-1 vif gene was constructed and the expression in vitro was detected at the transcriptional and protein levels, respectively. The results showed that DNA vaccine plasmid containing HIV-1 vif gene could express Vif protein in HeLa cells, which lay the basis for further animal experiments to detect the immunogenicity of HIV-1 vif DNA vaccine.
T cell responses play an important role in delaying the progression of AIDS and controlling viral replication and slowerring the decline of CD4+ T cells. To test cellular immune responses against HIV-1, the spleen cells were harvested and evaluated for antigen-specific T cell proliferation and CTL activity. The data show that HIV-1 Vif-specific T cell proliferation and CTL response can be detected in the splenocytes from the mice immunized with HIV-1 Vif DNA vaccine plasmid as compared with the control groups (p<0.01). In order to improve the immunogenicity of DNA vaccine in animal trials, the candidate DNA vaccine have been co-inoculated with the adjuvants such as genetic adjuvants that express cytokines or co-stimulatory molecules. GM-CSF, IL-6, and IFN-α improve the CTL activity and T cell proliferation induced by HIV-1 DNA vaccines (11,12,29). LIGHT is a member of the tumor necrosis factor superfamily and can function as a co-stimulatory molecule to increase T-cell proliferation and cell-mediated immune response in vivo (27). Although the exogenous administration of LIGHT to the airways induced fibrosis and smooth muscle hyperplasia (4), which indicated that LIGHT involved in inflammation, it was reported that the inflammation was good to improve the immunogenicity of DNA vaccine because the inflammation increased the recruitment of antigen-presenting cells at vaccination sites (22). In the present study, plasmid pcDNA-LIGHT containing the LIGHT gene was co-inoculated with HIV-1 DNA vaccine, and T cell responses were also investigated. The data show that the stronger T cell proliferation as well as the higher specific lysis were detected in the co-immunization group with pcDNA-Vif plus pcDNA-LIGHT as compared with the single immunization group with pcDNA-Vif plasmid alone, the result was similar to our previous studies (31). Gorgian Mohammady and colleagues also evaluated the effect of LIGHT on the kinetics of the CTL response induced by HSV-1 glycoprotein B-based DNA vaccine; the results showed that the CTL response in LIGHT adjuvanted group of HSV-1 DNA vaccinated mice was enhanced but not accelerated after booster, and the amount of granzyme B induced by the LIGHT containing vaccine was significantly higher than that induced by the vaccine without the adjuvant (8).
Antibodies usually play an important role in preventing infection. In order to detect the humoral immune responses in the five groups, the total antibodies against HIV-1 Vif elicited by HIV-1 DNA vaccine were analyzed by ELISA. The result indicated that the high levels of antibodies were obtained in the mice immunized with HIV-1 DNA vaccine, and a higher titer of anti HIV-1 Vif antibodies was detected in the mice co-immunized with HIV-1 DNA vaccine and LIGHT, which showed that LIGHT can increase the antibody responses induced by DNA vaccine. Unfortunately, we do not know if LIGHT can enhance the level of the neutralizing antibodies, because the neutralizing antibodies were not measured in the current study.
In summary, a DNA vaccine plasmid expressing HIV-1 Vif was constructed and the eukaryotic plasmid pcDNA-Vif could express the protein of interest in vitro. The inoculation of HIV-1 vif DNA vaccine could induce anti-HIV-1 Vif antibodies and T cell responses, whereas stronger humoral and cellular immune responses could be induced by co-administering the LIGHT gene. These results strongly suggest that HIV-1 vif DNA vaccine can be a candidate HIV-1 vaccine, and LIGHT can be a potential immunoadjuvant in DNA vaccination by enhancing the antibody production and T cell responses.
Footnotes
Acknowledgments
This work was supported by grants from National Natural Science Fund of China (30400070, 81072459), Program for New Century Excellent Talents in University (NCET-12-0179) and Shanghai Rising-Star Program (06QA14017).
Author Disclosure Statement
No competing financial interests exist.