
Abstract
This study examined cytokine gene expression patterns associated with fowl adenovirus (FAdV) infection. The selected cytokine mRNA was quantified by quantitative real-time reverse transcription-PCR in spleen, liver, and cecal tonsil during the course of infection of chickens with a serotype 8 FAdV (FAdV-8). Compared to uninfected chickens, infected birds had higher mRNA expression of interleukin (IL)-18 and IL-10 in spleen and liver, respectively. Interferon gamma (IFN-γ) mRNA expressed in spleen and liver of infected chickens was significantly upregulated, while the expression of IL-8 mRNA in spleen and liver of infected chickens was significantly downregulated. There was no significant difference between infected and uninfected groups in terms of cytokine gene expression in cecal tonsil. These results indicate that these four cytokines might play an important role in driving the immune responses following FAdV-8 infection.
Introduction
FAdVs have a worldwide distribution and some are associated with diseases such as inclusion body hepatitis (IBH). IBH is seen mainly in meat-type chickens 3–4 weeks of age (2). However, some FAdVs cause no or mild clinical signs and may be isolated from asymptomatic chickens, and are considered ubiquitous in poultry populations. IBH is characterized by sudden onset of mortality that usually ranges from 2%–10% in affected flocks. Mortality varies and is closely related to the pathogenicity of the virus, secondary infection with other pathogens, and susceptibility of the chickens. At gross examination, the main lesions are seen in the liver, which is swollen and light brown to yellow, occasionally with hemorrhages. Histologically, the basophilic nuclear inclusion bodies present in the hepatocytes are characteristic of infection (3). Most FAdV strains follow the same pattern of infection: after initial multiplication, the virus spreads to virtually all organs. Viremia is detected at 24 hours post-infection (hpi) and has two peaks, the first at 2 and the second at 7 days post-infection (dpi) (4). FAdVs are transmitted both vertically and horizontally. The virus is present in all excretions (e.g., fecal, tracheal, and nasal). Transmission within a flock is usually via the oral–fecal route (2,5,6).
Aviadenoviruses have the largest genome (43-45 kb) among adenoviruses, although genes of a few structural proteins such as V and IX, and early regions E1, E3, and E4 described in mastadenoviruses are not identified for aviadenoviruses (7 –11).
The early gene products of mammalian adenoviruses are responsible for host cell interactions and modulation of the immune response, downregulation of major histocompatibility complex (MHC-I), inhibition of cytolysis induced by tumor necrosis factor (TNF)-α, and prevention of interferon-mediated shut-off of protein synthesis (12,13).
Earlier we determined the pathogenicity of a fowl adenovirus serotype 8 (FAdV-8) isolate on the basis of clinical signs and pathological and histological examination. Viral genome copy numbers in the bursa of Fabricius, cecal tonsil, and liver after infection were also evaluated (8). Virus titer in feces and antibody response were also determined. Despite a lack of clinical signs and pathological changes, virus was identified in all selected tissues and cloacal swabs of all birds. The current work is a continuation of our efforts towards better understanding of the pathogenesis of FAdV infection with the emphasis on the role of interleukins, an area that was not studied so far.
Mammalian adenoviruses modulate cytokine expression by encoding intracellular proteins that counteract TNF-α (12,13). Booth and Metcalf (14) reported type-specific induction of IL-8 in human pulmonary epithelial cells upon infection with human adenovirus (HAdV) 7. Wu et al. (15) showed that HAdV-7 internalization is calcium dependent and is essential for IL-8 induction following infection. Additionally, HAdV-7 infection initiates secretion of IFN-γ inducible protein 10 (IP-10) from macrophages and epithelial cells (15). While extensive knowledge has been accumulated on the immunity to mammalian, primarily human, adenoviruses only the antibody response to FAdVs has been studied (2,16).
Our objective was to investigate the effects of FAdV infection on chicken immune responses, characterized by cytokine gene expression. Hence, the dynamics of cytokine gene expression in FAdV-8 infected chickens was determined. Interferon gamma (IFN-γ), interleukin (IL) 10, IL-18, and IL-8 gene expression was studied by quantitative real-time RT-PCR (qRT-PCR) in spleen, liver, and cecal tonsil at different days after FAdV-8 infection and compared to samples of uninfected chickens.
Materials and Methods
Virus and cells
FAdV-8 was isolated by the Animal Health Laboratory (AHL) at the University of Guelph and characterized in our laboratory (8). Virus propagation and titration were performed in CH-SAH cells as described by Alexander et al. (17).
CH-SAH cells were maintained in Dulbecco's Modified Eagle's Medium/Nutrient Mixture F-12 Ham (DMEM-F12) with 10% non-heat inactivated fetal bovine serum (FBS) as described (17).
Experimental design
Specific pathogen-free (SPF) eggs were obtained from the Canadian Food Inspection Agency (CFIA) (Ottawa, ON, Canada), hatched at the Arkell Poultry Station, University of Guelph, and the chicks were housed at the University Isolation Unit. Ten-day-old chicks were randomly divided into two groups, each group containing 50 birds. Group I was assigned for intramuscular (im) administration of the virus, each bird received 2×108 plaque forming units (pfu). Group II, the uninfected chicks (negative control), received PBS by im administration. Following inoculation, five chicks from each group were randomly drawn for necropsy at 1, 2, 3, 4, 5, 6, 7, 10, and 14 dpi. Birds were assigned for necropsy by simple random sampling of their identification numbers stratified by cage, using the statistic software (MINITAB 14). Euthanasia was performed according to the University of Guelph Animal Care Committee regulations (Isolation Unit, University of Guelph, ON, Canada) and the spleen, cecal tonsil, and liver were collected, and immediately submerged in RNA stabilization reagent (RNAlater, Qiagen, Mississauga, ON, Canada) and stored at −80°C until RNA extraction.
RNA extraction and cDNA synthesis
Total RNA was prepared from triplicates at each time point using TriZol (Invitrogen, Canada Inc., Burlington, ON, Canada) as per the manufacturer's protocol. Briefly, tissues were homogenized in 1 mL TriZol, followed by chloroform extraction, isopropanol precipitation, and a 75% ethanol wash of the RNA. The pellet was dissolved in 20 μL of RNAse-free water and the RNA concentration was measured by NanoVue spectrophotometer. To remove possible contaminating DNA, the samples were treated with a DNase based inactivation reagent (Invitrogen). The cDNA was synthesized with Random Primer (Invitrogen) and use of SuperScript™ II Reverse Trancriptase (Invitrogen), according to the manufacturer's instructions. The cDNAs were stored at −80°C until analyzed by quantitative real-time PCR.
Preparation of constructs for producing standard curves
Preparation of constructs and creation of standard curves for all cytokine genes, which were used in this study, as well as the β-actin gene were as described (18,19).
Quantitative real-time PCR
The expression of β-actin and cytokine genes from the cDNA samples was measured by quantitative real-time PCR. The reaction was prepared in a total volume of 20 μL of Qiagen Quantitect SYBR Green PCR kit (Qiagen) containing hot start Taq polymerase and cDNA. All reactions were carried out in 96-well plates (Roche Diagnostics GmbH, Mannheim, Germany). Each assay was run along with no-template control and three dilution series from the standard. The sequences and other features of specific primers for chicken IFN-γ, IL-18, IL-10, IL-8, and β-actin have been described (18,19) and given in Table 1.
The primers were synthesized by Laboratory Services, University of Guelph. The qRT-PCR conditions for different segments of each cycle were optimized for all target genes because the thermal cycling parameters differ for each gene (19). All samples were amplified in a LightCycler (LC480) instrument (Roche Diagnostics GmbH, Mannheim, Germany).
Data analysis
The efficiency of qRT-PCR was calculated by 10(-1/slope of the standard curve) (20). The mRNA expression was calculated relative to β-actin gene expression using the equation (Etarget)ΔCPtarget(control–sample)/(Eref) ΔCPref(control–sample) (21). In this equation, CP (crossing point) is the point at which the fluorescence rises significantly above the background. The difference in cytokine expression between groups was assessed by Mann-Whitney test and comparisons were considered significant at p≤0.05.
Results
The cytokine gene expression in the spleen, liver, and cecal tonsil of chickens infected with FAdV-8 was evaluated by a quantitative real-time reverse transcription-PCR.
Gene expression in spleen
The expression of IFN-γ, IL-18, IL-10, and IL-8 in spleen of FAdV-8 infected and uninfected control chickens is shown in Figure 1. The expression of selected interleukins was detected at all time points. There was a statistically significant increase (p≤0.05) in IFN-γ gene expression in spleen of infected chickens on 3 dpi. Also, at this time point, there was a significant (p≤0.05) decrease in IL-10 gene expression. However, when IL-18 gene expression was compared to that of uninfected controls, a statistically significant (p≤0.05) increase in expression was seen among infected chickens at 1 dpi. Expression of this cytokine changed on 7 dpi., when it was significantly downregulated (p≤0.05) in the infected group. IL-8 gene expression showed similar patterns to those of other cytokines in that it was expressed across all time points in the spleen of chickens of both infected and uninfected groups. Expression of this cytokine was significantly downregulated (p≤0.05) in the infected group at days 7 and 10 pi.
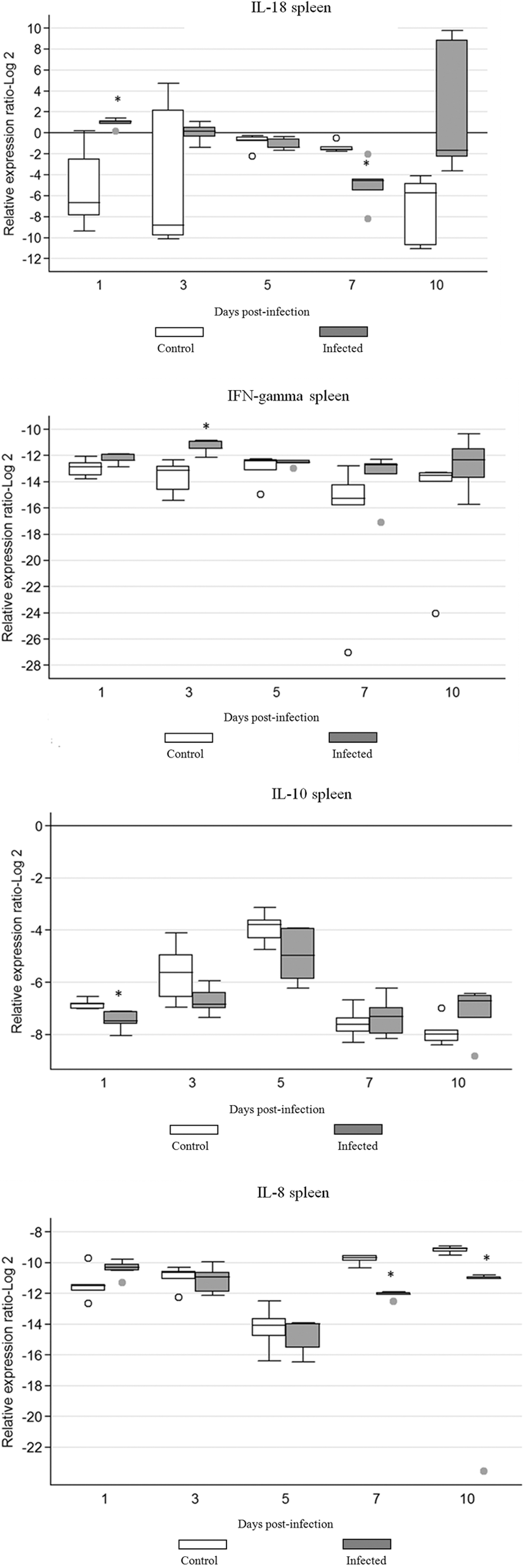
Cytokine mRNA expression in spleen of chickens infected with FAdV-8. The groups were as follows: uninfected (negative control) and FAdV-8 infected chickens. Each group contained 50 birds. Target and reference gene expression in spleen was quantified by real-time RT-PCR using SYBR Green. Target gene expression is presented relative to β-actin expression and normalized to a calibrator. The results are diagrammed as whisker boxes with medians. Boxes represent inter-quartile ranges and whiskers indicate extreme values. The difference in cytokine expression between groups was assessed by Mann-Whitney test and comparisons were considered significant at p≤0.05 (*). ○, ● symbols represent values which were identified as outliers. Numbers 1, 3, 5, 7, and 10 indicate days post-infection when the samples were collected.
Gene expression in cecal tonsil
There were no significant differences (p>0.05) in expression of mRNA for any of the tested cytokine genes, compared with the values for the appropriate uninfected controls (Fig. 2). Increased levels of IFN-γ, IL-18, and IL-10 mRNA were evident in infected birds at 1 dpi although the results were not statistically significant. Patterns of mRNA expression changed after 1 dpi for IFN-γ, IL-18, IL-10, showing a downward trend over the course of the infection. IL-8 gene expression was downregulated and although the expression of IL-8 was detected at all time points in both groups, there was no significant difference in the expression between groups (Fig. 2).
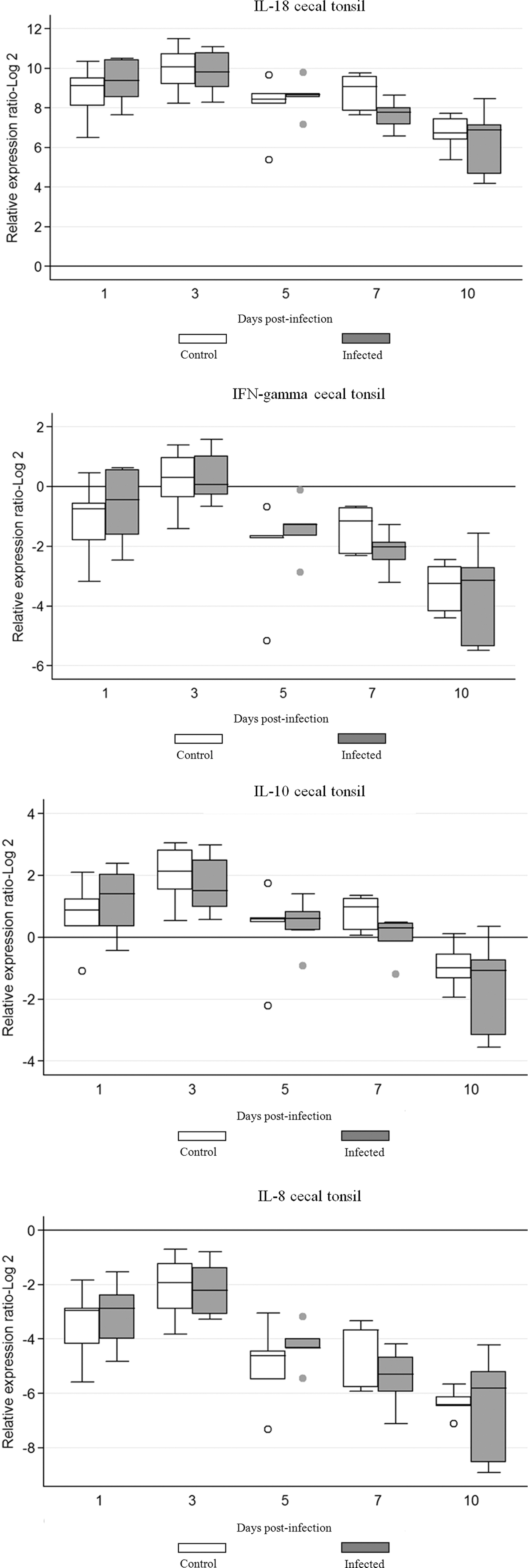
Cytokine mRNA expression in cecal tonsil of chickens infected with FAdV-8. The groups were as follows: uninfected (negative control) and FAdV-8 infected chickens. Each group contained 50 birds. Target and reference gene expression in cecal tonsil was quantified by real-time RT-PCR using SYBR Green. Target gene expression is presented relative to β-actin expression and normalized to a calibrator. The results are diagrammed as whisker boxes with medians. Boxes represent inter-quartile ranges and whiskers indicate extreme values. The difference in cytokine expression between groups was assessed by Mann-Whitney test and comparisons were considered significant at p≤0.05 (*). ○, ● symbols represent values that were identified as outliers. Numbers 1, 3, 5, 7, and 10 indicate days post-infection when samples were collected.
Gene expression in liver
The expression of IFN-γ, IL-18, IL-10, and IL-8 in liver of infected and control chickens is illustrated in Figure 3. Marked differences in expression of mRNA between infected and uninfected chickens were evident for all studied cytokines. There was a statistically significant increase (p≤0.05) in IFN-γ gene expression in liver of infected chickens on 3, 5, and 7 dpi. Infected chickens also had a significantly higher IL-10 gene expression compared to chickens in the uninfected control group at 1, 3, and 5 dpi. Additionally, adenoviral infection altered the expression levels of IL-8 and IL-18 and resulted in an increased expression (p≤0.05) of IL-18 at 3 dpi. On the other hand, the expression of IL-8 in the liver was downregulated and a decrease in expression became more prominent and statistically significant (p≤0.05) at 5, 7, and 10 dpi.
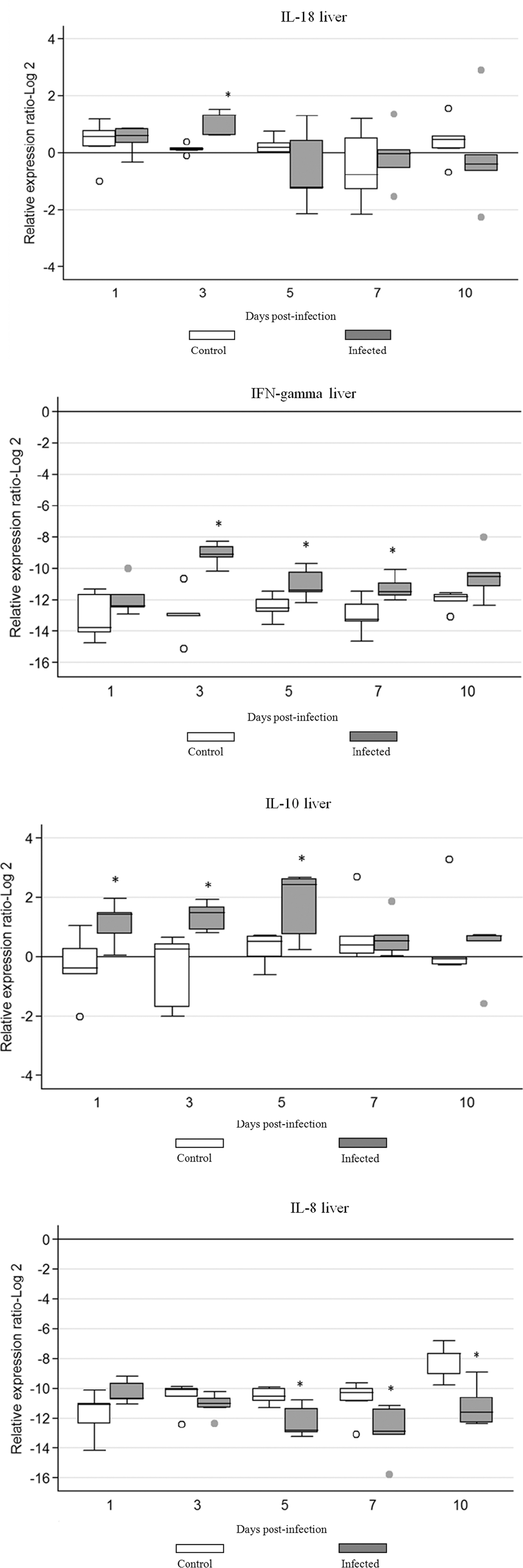
Cytokine mRNA expression in liver of chickens infected with FAdV-8. The groups were as follows: uninfected (negative control) chickens, and FAdV-8 infected chickens. Each group contained 50 birds. Target and reference gene expression in liver was quantified by real-time RT-PCR using SYBR Green. Target gene expression is presented relative to β-actin expression and normalized to a calibrator. The results are diagrammed as whisker boxes with medians. Boxes represent inter-quartile ranges and whiskers indicate extreme values. The difference in cytokine expression between groups was assessed by Mann-Whitney test and comparisons were considered significant at p≤0.05 (*). ○, ● symbols represent values that were identified as outliers. Numbers 1, 3, 5, 7, and 10 indicate days post-infection when samples were collected.
Discussion
Fowl adenoviruses are widely distributed in poultry operations and infect a wide range of tissues, but their ability to cause disease depends on numerous factors such as the type and age of the chickens, virulence of the virus, and secondary infection. The host defense mechanism in healthy birds usually functions effectively. However, some isolates of certain serotypes can cause considerable morbidity and mortality, especially among nutritionally or immunologically compromised birds, for example, those infected with chicken anemia virus or infectious bursal disease virus (22). Due to an increased interest in adenoviruses as vaccine and gene delivery vectors, it is important to understand the molecular aspects of infection and the host responses to these viruses (23). To our knowledge, this is the first study examining cytokine gene expression patterns associated with FAdV infection. We determined that IFN-γ, IL-18, IL-10, and IL-8 were activated early after FAdV-8 infection. The above cytokines represent chicken Th1, regulatory and pro-inflammatory cytokines, all of which have previously been shown to be involved in immunity to viral infections.
In the present study, we discovered that the expression of cytokine genes was spatially regulated. While there was no change in gene expression between infected and control animals in cecal tonsils, there were significant alterations in gene expression in spleen and liver of infected birds compared to uninfected controls. The reason for the absence of such alterations in cecal tonsils is perhaps due to the absence of histological changes in cecal tonsil recorded in our previous study (8).
The potent chemotactic cytokine IL-8 is known as an important mediator of inflammation that recruits and activates mammalian neutrophils to sites of infection (24). Overproduction of cellular IL-8 in viral infections is a reactive response of the host cells aimed at reducing viral load (25). In chickens (26), as in mammals (27,28), one of the roles of IL-10 is to repress the Th1 response. IL-10 aborts T cell responses to viral infections and can inhibit ongoing T cell activity. Several researchers (29 –32) have shown that IL-10 alters cytokine production and prevents maturation, depressing T cell activation.
In our study, a significant induction of the IFN-γ gene was noted in the spleen of infected chickens at 3 dpi. In the liver, expression of IFN-γ was significantly higher at 3, 5, and 7 dpi. IFN-γ is involved in inflammatory and immune responses and is an essential cytokine in the host defense against various pathogens. In addition to IFN-γ, IL-18 expression was also upregulated in spleen and liver. IL-18, an important regulator of innate and acquired immune responses, is expressed at sites of inflammation (33). Although originally identified as an inducer of IFN-γ production, IL-18 also enhances T and natural killer cell maturation, cytokine production, and cytotoxicity (34 –37). Therefore, it is plausible that the combination of IFN-γ and IL-18 expression in spleen and liver indicates polarization of T cells towards a Th1-like phenotype. Although the cellular source of cytokine was not determined in this study, it is possible that in addition to CD4+ Th1 cells, other cells, including CD8+ T cells and NK cells, can produce IFN-γ. It is also important to note that during viral infection, IL-18 activates CD8+ T cells and is important for viral clearance (38).
Concomitant with an upregulation in IFN-γ expression, we also noted an upregulation in IL-10 expression in the liver of infected chickens. IL-10 is considered a regulatory cytokine that is inversely correlated with the expression of Th1 cytokines such as IFN-γ. However, under certain circumstances such as high inflammatory responses to pathogens, it might be produced by Th1 cells (39). Furthermore, IL-12 stimulates human CD4 and CD8 T cells for increased production of both IFN-γ and IL-10 (40). Simultaneous expression of IFN-γ and IL-10 genes has also been reported by Johensen et al. (41), following infection of piglets with porcine reproductive and respiratory syndrome virus. Increased expression of IFN-γ and IL-10 genes in liver of infected chickens corresponds to the results of our previous study (8) where the highest number of viral copies in liver was recorded at 3, 5, and 7 dpi Moreover, histopathological examination of the liver showed mononuclear cell infiltration, degeneration, and necrosis of hepatocytes (data not shown) that could be positively correlated with increased expression of IFN-γ and IL-10 genes in liver. Similar to our findings, Diaz et al. (42) showed that HAdV-3 and HAdV-7 induce predominantly IFN-γ, which mediates cellular immunity. The IFN-γ increased production has been related to severity of infection since it activates macrophages and monocytes and induces TNF-α secretion that has antiviral function and activities.
We examined IL-8 expression, which is a pro-inflammatory cytokine. Expression of IL-8 mRNA was significantly downregulated in spleens of infected chickens at 7 and 10 dpi; similarly, significant downregulation was detected in the liver at 5, 7, and 10 dpi. These results are in accordance with the findings of Zhou et al. (43) and Lesokhin et al. (12). Interestingly, the GAM-1 protein, encoded by the Gam 1 gene (ORF8) at the right end of the FAdV-1 genome, has mastadenoviral E1 functions such as inhibition of apoptosis (7,44). A homologue of FAdV-1 ORF8 has been also identified for the FAdV-8 genome (8). Therefore, we speculate that some parts of the FAdV-8 genome share functions similar to mastadenoviral E1, E3, and E4 regions, despite a lack of obvious and high-sequence homology. Adenovirus encodes numerous products that counteract host defenses, ensure prolonged survival in the infected host, and consequently facilitate viral transmission. Zhou et al. (43) have demonstrated that porcine adenovirus 3 early region 1Blarge (E1Blarge) protein downregulates induction of proinflammatory cytokine IL-8 by inhibiting the NF-κB dependent gene transcription from human IL-8 promoter. The same authors suggested that the E1Blarge protein acts as a IkappaB (IκB) homolog and retains the ability to bind and inactivate NF-κB. Moreover, Lesokhin et al. (12) reported that human adenovirus E3 genes can drastically reduce the level of IL-8 mRNA. Inhibition of IL-8 mRNA by E3 is directly related to decreased secretion of IL-8 protein as measured by ELISA. The E3 region of HAdV encodes several immunoregulatory proteins. For example, four of six proteins of the E3 region are involved in immunoregulation (13). Bennett et al. (45) showed that E3-19K retains MHC-I in the endoplasmic reticulum and inhibits tapasin processing of peptides that bind to MHC-I. The E3-10.4K and E3-14.5K complex stimulates the internalization of proapoptotic receptors from the cell surface (46,47), while E3-14.7 inhibits TNF-α-induced apoptosis (48,49). Additionally, E3-14.7K inhibits TNF-α-induced production of arachidonic acid (50). It is unclear which E3 protein interactions are important in the inhibition of IL-8. The E3 region is placed between the pVIII and fiber protein genes, and deletion of this region results in an increased inflammatory response (51). We recently showed that FAdV-8, along with FAdV-1, FAdV-4, and FAdV-9, lacks mastadenoviral E1, E3, and E4 regions (8).
In conclusion, the results presented here showed that FAdV-8 infection in chickens is associated with enhanced expression of IFN-γ, IL-18, and IL-10 and reduced expression of IL-8 in spleen and liver. There was no significant difference between infected and uninfected groups in cytokine gene expression in cecal tonsil. Further work is necessary to shed more light on the role of cytokines in elicitation of the immune response to FAdV infections.
Footnotes
Acknowledgments
This work was supported by the Natural Sciences and Engineering Research Council of Canada (NSERC), the Canadian Poultry Research Council (CPRC), and the Ontario Ministry of Agriculture, Food and Rural Affairs (OMAFRA). The authors thank the Isolation Unit personnel for their professional animal care and assistance.
Author Disclosure Statement
No conflicting financial interests exist.