
Abstract
The largest population of people at risk for HCV-infection is injecting drug users (DU). We hypothesize that recurrent exposure to HCV, by continuing risk behavior, influences the development of an HCV-specific T-cell response. Therefore, we studied the association between repeated exposure to and the height and focus of the HCV-specific T-cell response in HCV antibody-positive injecting DU (n=18) with ongoing risk behavior (‘high-risk’), 9 with and 9 without detectable HCV-RNA), and 9 never-injecting DU (‘low-risk’, HCV-RNA+). Both total HCV-specific T-cell response, as well as the T-cell response against HCV nonstructural proteins, were significantly higher in injecting compared to never-injecting DU. Interestingly, the high-risk HCV-RNA¯ had no measurable CD4+ T-cell response to Core protein, compared to detectable responses to Core in the HCV-RNA+ group. Thus, both ongoing risk behavior and presence of HCV-RNA affect the HCV-specific T-cell response in both magnitude and specificity, which may have implications for vaccine development.
Introduction
We included HCV-antibody positive DU from the Amsterdam Cohort Studies (ACS) among DU (third generation commercial microparticle EIA system test from AxSym HCV version 3.0; Abbott, Wiesbaden, Germany) (14). The ACS is an open, ongoing cohort study initiated to study epidemiology and risk factors for HIV-1 infection. We included 18 injecting DU who reported recent injecting drug use (i.e., in the past 6 months). Most injecting DU reported at least daily injecting drug use (72.2%) and the median number of shared needles was 10 (range 5–700) in the 6 months preceding ACS entry. The median time since first injection was 7.3 years (IQR 3.5–9.1 years). From these injecting DU, 55.6% (10/18) were HCV-RNA positive, of which one patient resolved the infection (19777) (Table 1). Next to these, we included 9 HCV-infected self-reported never-injecting DU that were previously identified (13) from a total of 22 HCV-antibody positive never-injecting DU, of whom 15 (68.2%) were HCV-RNA+, remained chronically infected, and 9/15 had PBMC available for T-cell analyses.
p<0.05.
In DU with detectable HCV-RNA, genotype 3a was most common (7/9 never-injecting DU and 4/10 injecting DU). In the HCV-RNA¯ injecting DU, no individuals were HIV-positive, while in the two groups where HCV-RNA could be detected, the HIV prevalence was 20%–22%. Other general characteristics were similar in the three groups (Table 1).
To study the effect of ongoing risk behavior and presence of HCV-RNA on the HCV-specific T cell response, we measured HCV-specific CD4+ T-cell responses in DU with high-risk behavior with and without detectable HCV viral load and never-injecting DU using short-term culture of peripheral blood mononuclear cells (PBMC) (4,8). In short, PBMC were thawed and cultured for 12 days in the presence of recombinant HCV genotype 1 proteins [Core (C22–3), NS3 (C33c), NS4 (C100–3), and NS5A/B (NS5)] (kindly provided by Chiron/Novartis) and recombinant interleukin-2 (rIL-2) (Chiron, Uxbridge, UK). After 12 days, cells were washed, pooled, and rested overnight. On day 13, cells were restimulated using overlapping peptide pools corresponding to HCV genotype 1a (18-mers, 11 overlapping amino acids, Mimotopes, Australia) and the HCV-specific response was quantified by analysis of IFN-γ production using intracellular cytokine staining (ICCS) (15). Briefly, PBMC were stimulated for 6 h with HCV Core/E1/E2/p7, NS2, NS3, NS4, and NS5 peptide pools (2 μg/mL), and both αCD28 (2 μg/mL) and αCD49d (2 μg/mL) as co-stimuli, after 1 h 1:1,000 Brefeldin A (Golgiplug, BD Biosciences (BD), San José, CA) was added. As a negative control, PBMC were stimulated with medium and co-stimulation alone. As a positive control, PBMC were stimulated with 10 ng/mL PMA, 2 μg/mL ionomycin and co-stimulation. After stimulation, cells were washed, permeabilized (FACS Permeabilizing Solution, BD), washed again, and stained with antibodies specific for CD3, CD4, CD8, and IFN-γ (BD). After fixation (Cellfix, BD) min. 50,000 events were acquired on LSRII flow cytometer (BD). Lymphocytes were gated by forward and sideward scatter and data analyzed using FACSDiva software (BD). Results were expressed as the percentage of IFN-γ producing CD4+ T-cells, after subtracting the percentage of IFN-γ producing T-cells in unstimulated conditions. Subsequently, a more absolute number of HCV-specific effector T cells was calculated by including the proliferation rate. (total number of cells recovered after 12 days of expansion divided by the total number of cells put in to culture at day 0) (8,15). Because this assay is more sensitive compared to assays used in previous reports indicating very low or no HCV-specific T-cell responses in cHCV patients (1 –3,9,10,12), we were able to measure HCV-specific T-cell responses in most patients. Nonparametric tests (Kruskal-Wallis) were used to compare medians between groups; if there was a significant difference, Dunns' post-test was used to compare medians between groups. All performed statistical tests were two-sided. A p value ≤0.05 was considered to be statistically significant. All statistical analyses were performed using Graphpad and SPSS (Graphpad version 5.0, SPSS version 15.0.1, SPSS Inc.).
An HCV-specific CD4+ T-cell response could be detected in 20/27 DU (using a cut-off value of 150 IFN-γ producing CD4+ T-cells/million PBMC, based on previous experiments (data not shown). HIV-infected individuals had the lowest HCV-specific CD4+ T-cell response. As shown in Figure 1A, there was no difference in the height of HCV-specific CD4+ T-cell response between DU infected with HCV genotype 1 compared to those infected with another genotype. Irrespective of HCV-RNA presence, injecting DU had HCV-specific CD4+ T-cell responses of significant higher magnitude compared to never-injecting DU (p=0.012). Especially, the NS5-specific CD4+ T-cell response was significantly higher in injecting DU compared to never-injecting DU (p=0.0006). An HCV-specific CD4+ T-cell response was present in 6/9 HCV RNA+ never-injecting DU (66.7%), in 9/10 (90%) HCV RNA+ injecting DU, and in 5/8 HCV RNA¯ injecting DU (62.5%). The median height of the total HCV-specific CD4+ T-cell response was 294 IFN-γ producing CD4+ T-cells/million PBMC among RNA+ never-injecting DU and was significantly higher (1372 IFNγ producing T cells) among RNA+ injecting DU and RNA¯ injecting DU (1010 IFN-γ producing T cells), respectively (p=0.034) (Fig. 1B). The breadth of the HCV-specific CD4+ T-cell response (i.e., the number of different antigens targeted), was not different between groups (Fig. 1A). All 4 DU that were HIV co-infected had detectable HCV-RNA. Only one of them had a low, but detectable, HCV-specific CD4+ T-cell response targeting NS4 (237 HCV NS4-specific CD4+ T cells/million PBMC) (Fig. 1A, HIV-infected DU indicated with asterisk). CD4 counts ranged from 210 to 900 CD4+ T cells/mm3.
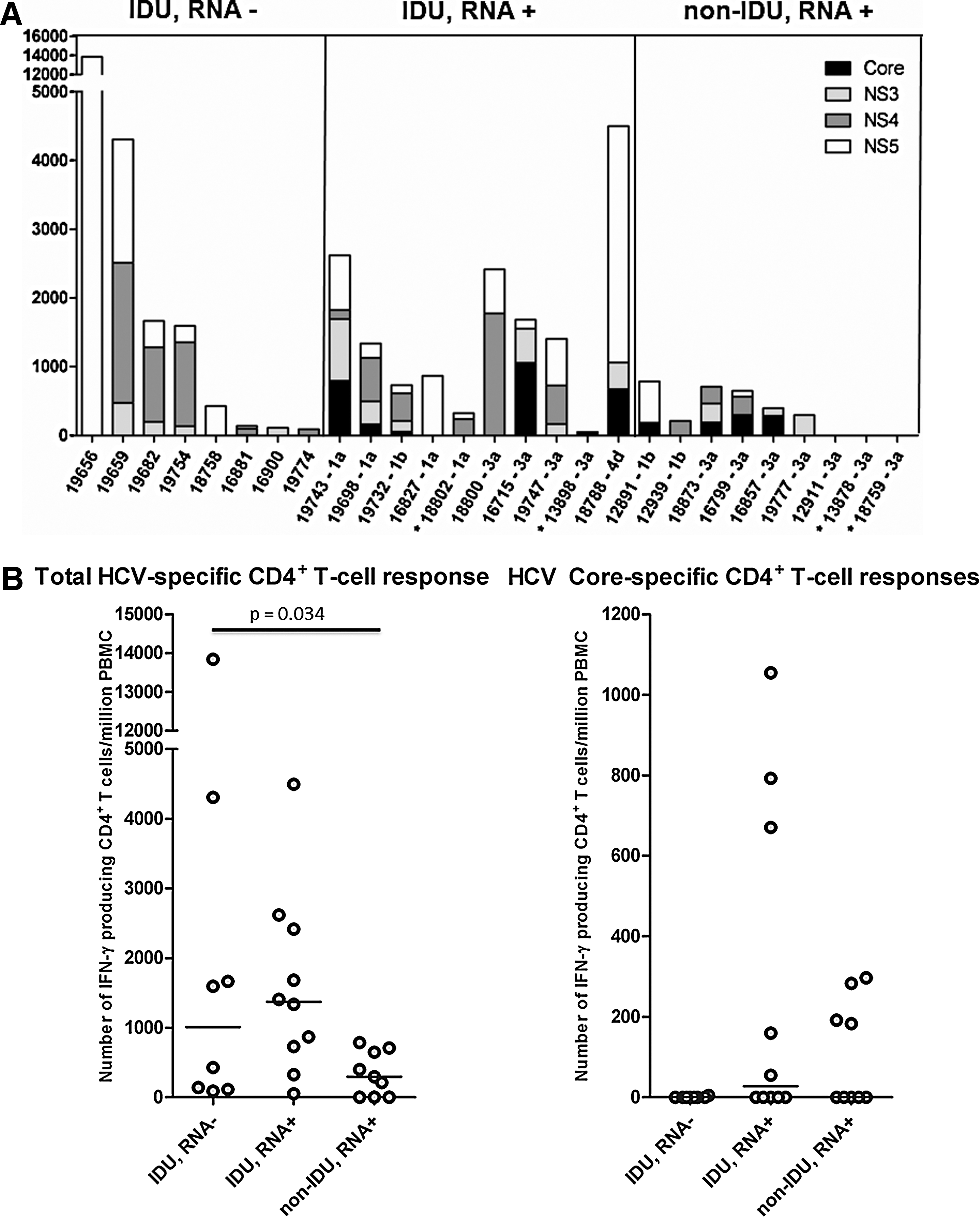
Interestingly, when we compared the specificity of the T-cell response between drug users with and without detectable HCV-RNA, we observed that none of the HCV-RNA¯ injecting DU had detectable HCV-specific T-cell responses targeting Core protein (0/8), compared to 4/10 HCV-RNA+ injecting DU and 4/9 HCV-RNA+ never-injecting DU. (Fig. 1A and B) The median Core-specific T-cell response in those with a detectable response was 28.7% (range 11.9–71.2%) of the total HCV-specific response (Fig. 1B). The only resolving individual (patient 19777) had a similar HCV-specific CD4+ T-cells response compared to the other HCV-RNA+ individuals, but did not have detectable core-specific T-cell responses, which is in line with previous data showing low core-specific responses in resolvers.(8)
Our data suggest that the T-cell response to HCV is differently affected by repeated exposure and persistence of HCV-RNA. Whereas continuous re-exposure seems to boost the immune response, the presence of HCV-RNA is associated with T-cell responses directed at Core protein. Frequent injecting and sharing of needles in injecting DU can be seen as a proxy for frequent exposure to HCV. Although HCV re-infection may not necessarily have occurred after each shared needle, it is plausible to assume that the chance of exposure was very high, since HCV-prevalence ranged between 63% and 93% in Amsterdam in the 20 years since the start of the ACS (14). This repetitive exposure is likely to have boosted the immune system, leading to higher HCV-specific T-cell responses in injecting DU, similar to HIV-specific T-cell boosting in HIV-negative African prostitutes (7). Although the frequency of injecting drug use and the number of shared needles was very high in the selected injecting DU, 44.4% of them were HCV-RNA¯ at the first ACS visit, suggesting they were (at least temporarily) able to control HCV. It is possible that this high HCV-RNA¯ proportion in these DU with a long history of high-risk injection drug use has another reason (e.g., survival bias in which HCV-RNA+ DU with similar risk behavior may have deceased before study entry from HIV or other causes related to hard drug use).
In addition, we show that both injecting and never-injecting HCV-RNA+ DU have a higher HCV Core-specific CD4+ T-cell response in comparison to injecting HCV-RNA¯ DU. As the presence of HCV-RNA in serum is indicative of active viral replication in the liver, this also denotes continuous antigen production. The amount of Core protein present in serum is correlated with the height of HCV viral load (5,6). When more Core antigen is produced, this will probably result in skewing of the T-cell response. Since HCV is translated as a polyprotein, it is unlikely that more Core protein than NS proteins is produced (11). Nevertheless, T cells might be more exposed to Core than to NS proteins since it has been shown that nonenveloped HCV nucleocapsids occur naturally in the serum of HCV-infected individuals and that Core protein is abundantly present in serum of HCV-infected patients (and chimpanzees) (5,6).
In conclusion, our results suggest that repetitive exposure to HCV by frequent injection drug use and a high number of shared needles affect the height of the HCV-specific immune response, suggesting boosting of the immune system that may be favorable. On the other hand, the presence of HCV-RNA affects the specificity of the HCV-specific CD4+ T-cell-response. Both findings may impact the interpretation of immunological parameters in IDU and may have implications for future vaccine development.
Footnotes
Acknowledgments
The Amsterdam Cohort Studies on HIV infection and AIDS, a collaboration between the Health Service of Amsterdam, the Academic Medical Center of the University of Amsterdam, Sanquin Blood Supply Foundation and the University Medical Center Utrecht, are part of the Netherlands HIV Monitoring Foundation and financially supported by the Netherlands National Institute for Public Health and the Environment. Website:
Part of the data shown in this manuscript was presented as a poster at 13th International Symposium on Viral Hepatitis and Related Liver Disease (March 20–24, 2009, Washington DC).
Author Disclosure Statement
No competing financial interests exist.