
Abstract
Lymphoid organs play an important role in prion disease development and progression. While the role of lymphoid organs and changes in immune-related genes have been extensively investigated in scrapie-infected animals, innate immunity has not. Previous studies examined lymphocyte function in scrapie-infected C3H/HeJ mice, which exhibit defects in lipopolysaccharide (LPS) response now known to result from a mutation in Toll-like receptor (TLR) 4. We examined immune function in scrapie-infected CD1 mice, which are LPS responders. Lymphocyte proliferation from CD1 mice infected with either 139A or ME7 scrapie was measured in response to concanavalin (Con) A or LPS at 1 and 3 months after infection. Following LPS exposure, mice infected 3 months with ME7, but not 139A, demonstrated significantly decreased lymphocyte proliferation compared to controls. After Con A exposure, lymphocyte proliferation in scrapie-infected mice did not differ from controls. Gender-specific comparison of lymphocyte proliferation showed significant decreases in mitogenic responses in females infected 3 months with either 139A or ME7, compared to controls. Males infected for 3 months with ME7, but not 139A, showed significantly decreased proliferation after lymphocyte exposure to LPS, but not Con A. Neither gender showed changes in lymphocyte proliferation after 1 month of scrapie infection. Innate immune activation of peritoneal macrophages was determined via production of nitric oxide (NO), IL-6, and TNF-α after exposure to TLR ligands. TNF-α and IL-6 production were reduced in macrophages from females infected with either scrapie strain for 3 months, while NO production after TLR agonist plus IFN-γ exposure was decreased in both females and males infected for 3 months with 139A, compared to ME7. These data demonstrated altered innate immunity, suggesting hormonal and/or other gender-specific regulation may contribute to gender differences in some immune functions. Our data demonstrate lymphocyte proliferation and innate immune functioning in scrapie-infected mice deteriorate with disease progression.
Introduction
Using a mixed lymphocyte reaction assay, Kingsbury et al. (33) demonstrated normal cell-mediated immunity in scrapie- infected mice compared to uninfected mice. Mitogenic proliferation using Con A was also unchanged in mice 14 to 70 days after inoculation of scrapie-infected brain homogenate. Cellular proliferation was performed in C3H/HeJ mice, LPS non-responders (Tlr4-/-) (23,44,48). In contrast, Garfin et al. (19) reported that scrapie-infected mice suppressed polyclonal B cell activation using LPS in C3H/HeJ mice, although the response to Con A was unaffected. Cellular proliferation using LPS in C3H/HeJ mice needs re-evaluation using an LPS responder mouse strain. Innate immune function in scrapie-infected mice has not been studied. In this study, immune functions of both female and male mice were investigated 1 and 3 months post injection of scrapie-infected brain homogenates in CD1 mice, LPS responders.
We examined the innate immune function of macrophages using TLR ligands including LPS, CpG ODN and zymosan with or without IFN-γ and lymphocyte proliferation of splenocytes including T and B lymphocytes using the mitogens Con A and LPS.
Methods
Reagents
LPS W (Escherichia coli 0111:B4) was purchased from BD Biosciences (Sparks, MD). RPMI 1640, murine recombinant interferon-gamma (rIFN-γ), penicillin, and streptomycin were obtained from Invitrogen (Carlsbad, CA). Heat inactivated fetal calf serum (FCS) was purchased from Gemini Products (Woodland, CA). CpG ODN 1826 [5′-TCC ATG A
Animals
CD1 mice were obtained from Jackson Laboratories (Bar Harbor, ME). Animals were fed and given water ad libitum and maintained in artificial light with a 12-hour on, 12-hour off cycle. Mice used in this study were 5–6 weeks of age.
Inocula
Scrapie strains used in this study were selected for study because these have among the shortest incubation periods of all mouse scrapie strains (14,27,36). The ME7 and 139A scrapie strains were originally provided by Drs. Alan Dickinson and Richard Kimberlin (Neuropathogenesis Unit, Edinburgh, UK), respectively. These strains were passaged and maintained in our laboratory as previously described (13). Whole brain homogenates were prepared as 1% w/v suspensions in PBS, and 0.025 mL was injected into mice intracerebrally. Age-matched control mice were injected with brain homogenates prepared from normal mice. The intracerebral infection route was chosen for this study because this route has the shortest and most consistent presymptomatic incubation period compared to peripheral routes of administration.
Preparation of splenocytes and peritoneal macrophages
After the spleens were removed, spleen and body weight were measured. A spleen index was calculated by (spleen weight/body weight) x 1000. Splenocytes were prepared after lysis of red blood cells using lysis buffer. Cells suspended in complete media containing RPMI 1640 plus 10% FCS and penicillin/streptomycin were counted by two independent researchers. After splenocytes (2×105/well) were added to a 96-well plate, LPS (1 μg/mL) or Con A (2.5 μg/mL) were added for mitogenic stimulation for B and T lymphocytes, respectively. For preparing peritoneal macrophages, mice were injected intraperitoneally with 1 mL of 4% thioglycollate (BD Biosciences, Sparks, MD). Four days after thioglycollate injection, peritoneal macrophages were obtained by peritoneal lavage with phenol red-free HBSS (without Mg++ and Ca++) (26). Macrophages were suspended in complete media and cells were counted by two independent researchers. Differential staining showed greater than 98% purity of macrophages. After 2 h adherence in a 96-well plate (2×105 cells/well) in 5% CO2 at 37°C, nonadherent cells were removed using complete media. Three ligands for TLRs including LPS (TLR4, 1 μg/mL), CpG ODN (TLR9, 1 μM), and zymosan (TLR2, 50 μg/mL) with or without IFN-γ (50 U/mL) were added for macrophage activation. After 24 h incubation, supernatants were collected and frozen at −70°C.
Cellular proliferation assay
Splenocytes (2×105 cells /well) were activated with Con A (2.5 μg/mL) or LPS (1.0 μg/mL) for 72 and 48 h at 37°C in a 96-well plate, respectively (42). One μCi of H3-thymidine was added to each well 4 h before collection. H3-thymidine incorporation was measured using a liquid scintillation counter (Packard Instrument Co., Meriden, CT).
Nitrite determination
Nitrite was measured by the method described previously (43). Briefly, 25 μL of supernatant from a 96-well plate culture was mixed with an equal volume of Griess reagent (1% sulfanilamide, 0.1% naphthylethylene diamine dihydrochloride, and 2.5% phosphoric acid) and incubated at room temperature for 10 min. The optical density of each sample was read at 550 nm using a microplate reader (Bio-Tek Instrument, Winooki, VT). The calibration curve was prepared using sodium nitrite as a standard.
Measurement of TNF-α and IL-6 using ELISA
Secretion of TNF-α was measured using Duoset ELISA kits for TNF-α (R & D Systems, Minneapolis, MN) by a modified method described previously (43). Briefly, 50 μL of capture antibody, goat anti-mouse TNF-α antibody, diluted 1:180 in phosphate-buffered saline (PBS) with 1% BSA was incubated in a 96-well ELISA plate (Thermolab Inc., Chantilly, VA) overnight at 4°C. After washing the plate with PBS-tween 20 (T-PBS, 0.01 M PBS with 0.05% Tween 20), 1% BSA and 5% sucrose in PBS with 0.05% NaN3 was added, and the plate was incubated at 37°C for 2 h. The plate was washed with T-PBS and 50 μL of either various concentrations of standard murine rTNF-α and diluted samples were added to the plates. After incubation at 4°C overnight and washing with T-PBS, 50 μL of biotinylated detecting antibody, goat anti-TNF-α antibody, diluted 1:180 in PBS with 1% BSA was added, and the plate was incubated at 37°C for 2 h. After washing, the plate was incubated with 50 μL of streptoavidin-horseradish peroxidase conjugate diluted 1:200 in PBS with 1% BSA for 1 h. The color was developed with tetramethylbenzdine and H2O2 in citrate buffer (pH 5.0) for 10 min. The plate was read at 450 nm using a microplate reader. The concentration of TNF-α and IL-6 released into the medium was calculated by using murine rTNF-α or IL-6 as a standard, respectively.
Statistical analysis
Differences between control and experimental groups were determined by Student's independent t-test, as well as LSD and Tukey analysis under breakdown and ANOVA using Statistica (Statsoft, Tulsa, OK). Statistical significance of differences between the groups is considered at the level of p<0.05.
Results
Innate immunity and proliferation of lymphocytes were examined after mice were infected with either scrapie strains, 139A or ME at 1 and 3 months after infection. Mice developed neurological abnormalities, including abnormal gait 3 months after injection of scrapie-infected brain homogenate and death occurred within 3–4 weeks after development of neurological symptoms.
Lymphocyte proliferation of splenocytes
As the innate immune system regulates the B and T cells of the acquired immune system through the Toll-like receptors, we measured cellular proliferation by mitogenic stimulation of splenic B and T lymphocytes using LPS (1 μg/mL) and Con A (2.5 μg/mL), respectively (39). Immune function was measured 1 and 3 months after ic injection of either 139A or ME7 strains of scrapie. The spleen index showed no significant difference in scrapie-infected mice compared to age- and sex-matched controls (Table 1). Immunity as measured by cellular proliferation measured 1 month after 139A or ME7 scrapie infection was not significantly different compared to controls. In contrast, cellular proliferation using LPS or Con A 3 months after scrapie infection was reduced 58% and 32%, respectively. Cellular proliferation was significantly reduced in mice infected with ME7, not with 139A (Table 2). Data show a significant decrease of cellular proliferation using LPS in female mice 3 months after scrapie infection with 139A (58%) or ME7 (53%) (Fig. 1A). Cellular proliferation using Con A in female mice 3 months after infection was also reduced in both 139A (51%) and ME7 (52%) (Fig. 2A). Cellular proliferation using LPS in males 3 months after infection was different from females (Fig. 1A and 1B). Mitogenic stimulation using LPS in males 3 months after scrapie ME7 infection was decreased 63%. There was no difference in proliferation using both Con A and LPS in males or females infected with scrapie for 1 month compared to controls.
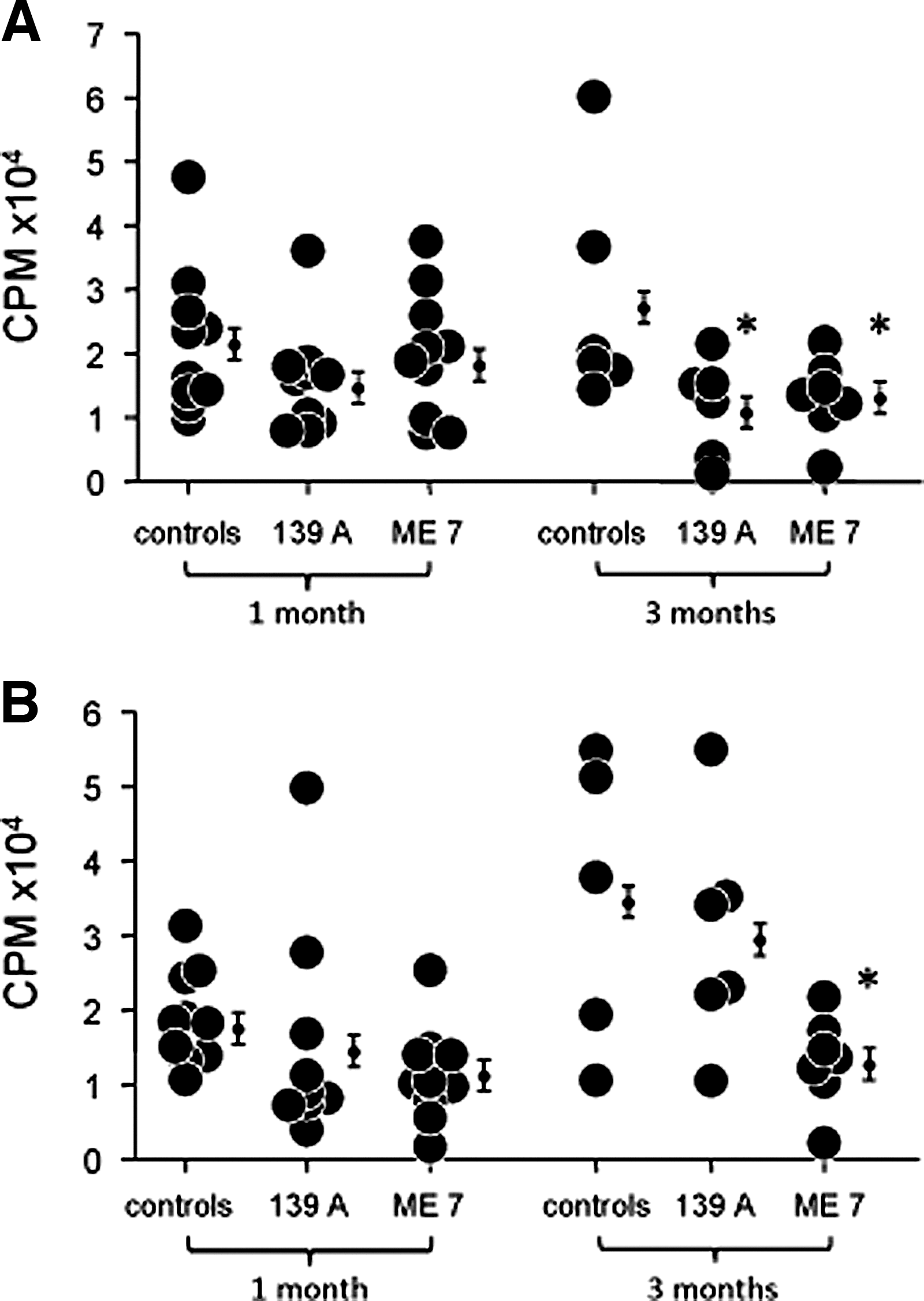
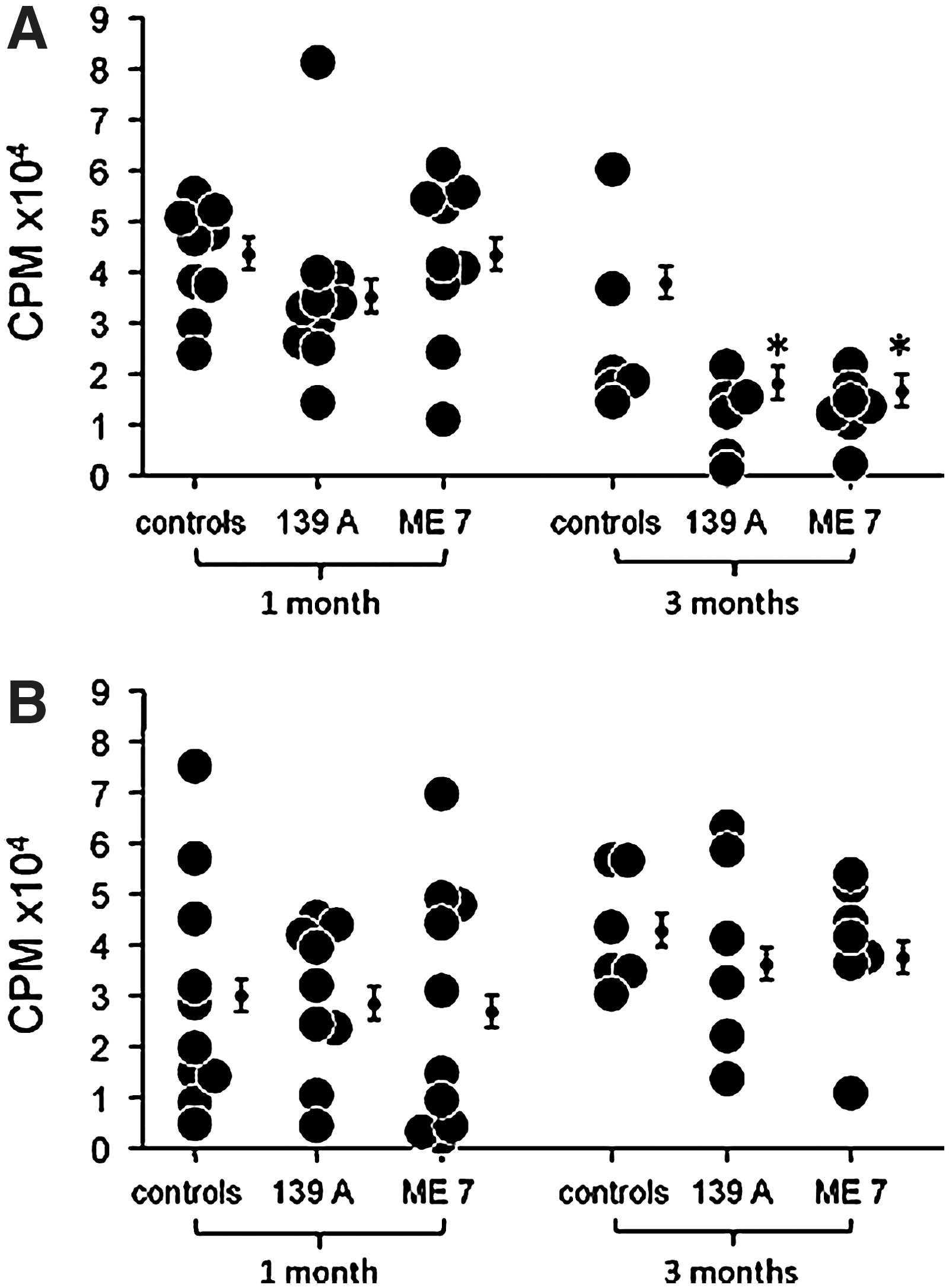
SI, (spleen weight/body weight)*1000; bData are mean±SE.
Data are mean±SE (CPM). a N=14; b p<0.05, compared to controls.
Innate immune function of macrophages
To evaluate innate immunity in scrapie-infected mice, TLR agonists including LPS (TLR4), zymosan (TLR2), and CpG ODN (TLR9) were used. Cardinal parameters for TLR agonist-induced inflammation include NO, TNF-α, and IL-6. These proinflammatory mediators were measured in peritoneal macrophages activated with LPS (1 μg/mL), zymosan (50 μg/mL), or CpG ODN (1 μM) in the absence or presence of IFN-γ (50 U/ml) (25,39 –41). Macrophages obtained from peritoneal lavage four days after i.p. thioglycollate injection were pooled from four mice.
NO production in macrophages activated with CpG ODN plus IFN-γ was decreased significantly in females infected for both 1 and 3 months with either scrapie strain (Table 3). NO production using zymosan, LPS or CpG ODN plus IFN-γ, was significantly decreased in females and males after scrapie infection for 3 months with 139A, not with ME 7 (Tables 3 and 4). In males infected with scrapie 139A for 3 months, NO production with TLR agonists plus IFN-γ was significantly reduced.
W/o or w/ means absence or presence of IFN-γ (50 U/mL).
The concentrations of LPS, CpG ODN and zymosan are 1 μg/mL, 1 μM, and 50 μg/mL, respectively.
ND, not detected. Data are mean±SD (μM) from samples measured in triplicate.
p<0.05, statistically significant. NO production with IFN-γ all was 7.4±1.5 μM. Peritoneal macrophages were pooled from 4 mice. Similar results were obtained from an additional experiment.
W/o or w/ means absence or presence of IFN-γ (50 U/mL).
The concentrations of LPS, CpG ODN and zymosan are 1 μg/mL, 1 μM, and 50 μg/mL, respectively.
ND means “not detected”.
Data are mean±SD (μM) from samples measured in triplicates; * p<0.05, statistically significant.
NO production with IFN-γ al1 was 5.3±0.8 μM. Peritoneal macrophages were pooled from 4 mice. Similar results were obtained from an additional experiment.
TNF-α secretion from LPS activated cells was reduced in females infected for 1 month and 3 months with 139A (Table 5). TNF-α secretion from LPS plus IFN-γ activated cells was increased in males for 1 month infected with ME7 (Table 6). TNF-α secretion using CpG ODN with and without IFN-γ for macrophage activation was decreased in females infected with either 139A or ME7 for 3 months, not for 1 month. However, TNF-α secretion using LPS with and without IFN-γ for macrophage activation was reduced in both females and males infected with 139A, and TNF-α secretion induced with LPS only was reduced in both sexes infected with either strain. TNF-α secretion using TLR agonists plus IFN-γ was reduced in males infected with 139A for 3 months. (Tables 5 and 6).
W/o or w/ means absence or presence of IFN-γ (50 U/mL).
The concentrations of LPS, CpG ODN and zymosan are 1 μg/mL, 1 μM, and 50 μg/mL, respectively.
Data are mean±SD (μg/mL) from samples measured in triplicates.
p<0.05, statistically significant, compared to controls.
TNF-α with IFN-γ al1 was not detected.
Peritoneal macrophages were pooled from 4 mice. Similar results were obtained from an additional experiment.
W/o or w/ means absence or presence of IFN-γ (50 U/ml).
The concentrations of LPS, CpG ODN and zymosan are 1 μg/mL, 1 μM, and 50 μg/mL, respectively.
Data are mean±SD (μg/mL) from samples measured in triplicates.
p<0.05, statistically significant.
TNF-α with IFN-γ al1 was not detected.
Peritoneal macrophages were pooled from 4 mice. Similar results were obtained from an additional experiment.
IL-6 production using LPS with and without IFN-γ was significantly decreased in females infected with ME7 at 3 months after infection (Table 7). IL-6 production using CpG ODN with IFN-γ was significantly downregulated in females infected with 139A for 3 months, but not at 1 month after infection. IL-6 production using zymosan with and without IFN-γ was significantly decreased in ME7 scrapie-infected females at 3 months after infection. IL-6 production using LPS or zymosan without IFN-γ was reduced in males infected with 139A or ME7 3 months after infection, respectively (Table 8). Overall, IL-6 appeared to be more downregulated in macrophages from females compared to males activated with and without IFN-γ.
W/o or w/ means absence or presence of IFN-γ (50 U/mL).
The concentrations of LPS, CpG ODN and zymosan are 1 μg/mL, 1 μM, and 50 μg/mL, respectively.
Data are mean±SD (μg/mL) from samples measured in triplicates.
p<0.05, statistically significant.
IL-6 with IFN-γ al1 was not detected.
Peritoneal macrophages were pooled from 4 mice. Similar results were obtained from an additional experiment.
W/o or w/ means absence or presence of IFN-γ (50 U/mL).
The concentrations of LPS, CpG ODN and zymosan are 1 μg/mL, 1 mM, and 50 μg/mL, respectively.
Data mean±SD (μg/mL) from samples measured in triplicates.
p<0.05, statistically significant.
IL-6 with IFN-γ al1 was not detected.
Peritoneal macrophages were pooled from 4 mice. Similar results were obtained from an additional experiment.
Discussion
TLRs are known for the initial recognition of pathogen associated molecular patterns (PAMPs) and the subsequent signal transduction that determines the ultimate fate of infection (28,54). TLR4 is essential for response to LPS, a major constituent of the outer membrane of Gram-negative bacteria, a potent immunostimulatory molecule and the cause of septic shock (23,30,44,48). TLR2 recognizes a wide range of PAMPs derived from various pathogens, ranging from bacteria, fungi, parasites, and viruses. Zymosan, a β-glucan of yeast cell wall, is an agonist for TLR2 and dectin-1, as well as a stimulant for macrophages to produce NO, IL-6, and TNF-α in various in vivo animal models and/or in vitro cell culture models (9,16,21,26,29,47,49). Microbial DNA serves as the physiological ligand for TLR9, which is present in the intracellular endosomal compartment, differing from membrane-bound TLRs (5,15,20,53). Discrimination between foreign and host DNA is dependent upon recognition of unmethylated CpG dinucleotides frequently within microbial DNA. The ability of unmethylated CpG motifs, either within the context of bacterial DNA or short single-strand synthetic oligodeoxynucleotides (ODNs) to activate the innate immune response is well documented (20,22,54,55). As the innate immune system regulates the acquired immune system, we measured innate immunity and proliferation of lymphocytes after mice were infected with scrapie strains, either 139A or ME7. Immune function including cellular proliferation and innate immunity was significantly decreased in mice 3 months after i.c. injection of brain homogenates infected with either strain. However, immune function was unchanged at 1 month in mice infected with either strain. Mice developed neurological abnormalities, including abnormal gait, 3 months after injection of scrapie-infected brain homogenate and death occurred within 3–4 weeks after development of neurological symptoms. Garfin et al. (19) demonstrated suppression of B cell activation using LPS in C3H/HeJ scrapie infected mice, although T cell activation using phytohemagglutinin and Con A was unaffected. Our current study is consistent with suppression of B activation and no change of T cell activation in scrapie-infected mice. Experiments by Garfin et al. performed prior to the discovery of TLRs used mice (C3H/HeJ) that were non-responders to LPS. Poltorak et al. (44), Qureshi et al. (48), and Spinner et al. (52) demonstrated that C3H/HeJ and C57BL/10ScCr are mutant strains (Tlr4-/-) lacking Tlr4, an agonist for LPS. Peritoneal macrophages from the responder strain (C3H/OuJ, Tlr4+/+) produced IL-6 and TNF-α using a synthetic fragment of scrapie protein. The non-responder strains (C3H/HeJ, Tlr4-/-) did not produce these cytokines (52). These data, in part, support the lengthened life-span of C3H/OuJ compared to that of C3H/HeJ, suggesting TLR-4 may be involved in the pathogenesis of prion diseases. In contrast, Kingsbury et al. (33) reported a normal immune response as measured by mixed lymphocyte reaction and mitogenic activation 14–70 days after infection with scrapie. The data indicate no obvious splenomegaly was present in infected BALB/c and C57BL/6 mice throughout infection. C3H/HeJ mice demonstrated transient mild splenomegaly at 42 and 49 days. Our data demonstrated no splenomegaly in mice infected at 1 and 3 months (Table 1).
The current study examined LPS-mediated cellular proliferation using an LPS responder mouse strain, and the data show that mitogenic B cell activation was suppressed (Table 2). The data further demonstrate that mitogenic activation differed with regard to gender (Figs. 1 and 2). Comparison of gender differences in cellular proliferation is also dependent on the strain of scrapie. LPS activation was suppressed in females 3 months after infection with both scrapie strains and was suppressed only in ME7 infected male mice (Fig. 1A and B). Con A activation was reduced in females 3 months after infection with both scrapie strains, but was not reduced in male. These data indicated that hormones may be involved in cellular proliferation. Similarly few changes in innate immunity were demonstrated in both females and males infected with either strain for 1 month. The innate immune response was decreased in females infected with either strains for 3 months and in males infected with 139A for 3 months, except for IL-6 production (Tables 3 –8). Although NO production induced with TLR agonists plus IFN-γ are reduced in both females and males infected with 139A, not ME7, TNF-α with and without IFN-γ was decreased in males infected only with 139A for 3 months. The innate immune response in scrapie-infected animals is complex and may depend both on gender and scrapie strains, not on specific TLR agonists. Production of NO, TNF-α, and IL-6 induced with various TLR agonists including LPS, CpG ODN, and zymosan in the presence of IFN-γ, was inhibited greater than in the absence of IFN-γ. These results suggest that immune function with a secondary infection may deteriorate more readily in scrapie-infected animals. There is some evidence that in the spleen ME7 may preferentially target follicular dendritic cells (FDC) for infection, while 139A (also referred to as Chandler strain and RML strain) targets both FDC and lymphocytes (36). Difference in cell types infected may result in some of the functional change seen in this report.
McFarlin et al. (38) demonstrated that T-cell-mediated immunity neither contributes to the pathogenesis of scrapie nor plays a significant role in the host's defense against the agent. Neonatal thymectomy did not alter the incubation period or pathology in mice inoculated i.c. with scrapie. They studied the course of scrapie produced by i.p. inoculation in thymectomized, lethally irradiated mice that were reconstituted with fetal liver cells. Although the mice were markedly depleted of thymus-derived (T) lymphocytes and were incapable of rejecting allogeneic skin grafts, the response to the scrapie agent as measured by life-span was not different from that of normal mice. These data suggest that scrapie infection does not induce interferon synthesis, proinflammatory cytokines, and immunosuppression with cyclophosphamide (6). A potential explanation for this lack of immune response to the infectious scrapie particle came with the discovery that the major protein component of scrapie protein (PrPsc) is a modified form of a host cellular protein (PrPc); thus organisms may not recognize the scrapie agent as foreign (6). Mice may be tolerant to both self PrPc and infectious scrapie PrPsc, due to the clonal deletion of PrP-reactive T cells during thymus development. Although T cell recognition of PrPc and PrPsc was lacking, innate immunity was activated by peptides made from prion proteins as indicated by production of TNF-α and IL-6 (52), indicating potential prevention and treatment through active and passive immunization.
The current study demonstrated that lymphocyte proliferation using LPS was decreased in mice infected with ME7 for 3 months while lymphocyte proliferation using Con A in most groups was unchanged. Moreover, lymphocyte proliferation using both LPS and Con A in either strain was significantly suppressed in females compared to males infected with 139A. Similarly, innate immune function in females infected with either strain for 3 months was reduced greater than males infected with ME7. The decrease of immune function in male mice is scrapie strain specific. Previous studies have in some cases shown differences in scrapie pathogenesis dependent on the interaction between gender and route of infection (1,31,32,34), although dependent on the strain of scrapie being studied (32). Our study extends those findings and further shows that immune function deteriorates with increased neurological disease in scrapie-infected CD-1 mice and may be regulated by sex hormones and/or other gender-specific differences. Gender-specific differences in CD-1 mouse immunity to fungal and viral infections has also been observed (10,41).
Previous studies by others have demonstrated alterations in immune-specific gene expression and anatomy during scrapie infection in mice and sheep (8,17,18,39). Our previous study in TLR4-mutant C3H/HeJ and TLR4 wild-type mice suggested that innate immune responses serve a protective effect in scrapie (52). In the current study, worsening immune functions coinciding with increasing scrapie progression suggests acceleration of disease progression may occur as immune responses lose the ability to resist the infection. Therefore, methods that bolster immune responses may potentially serve to decelerate prion disease progression. As our current study examined immune functions in mice infected with scrapie via the intracerebral route, the extent to which our findings extend to prion infections acquired through peripheral routes (e.g., orally-acquired variant CJD) is uncertain and warrants replication in peripherally administered scrapie models.
Footnotes
Acknowledgments
This work was supported by the New York State Office for People with Developmental Disabilities, NIH Contract N01-NS-0-2327 to R.J.K., and Alzheimer's association/Stranahan Foundation grant NIR-04-1162 to D.S.S.
Author Disclosure Statement
We declare no conflict of interest.