
Abstract
Intravaginal (ivag) infection of mice with herpes simplex virus type 2 (HSV-2) causes genital tissue damage, quickly followed by development of fatal encephalopathy. To delineate initial host responses generated by HSV-2 infection, here oligonucleotide microarrays compared gene expression in vaginal tissue from uninfected mice and mice 1, 2, 3, 4, 5, 6, or 7 days after ivag infection with 104 pfu HSV-2. While comparison of mRNA expression in uninfected and HSV-infected vaginal tissue detected few changes during the first 2 days post infection (dpi), there were 156 genes whose expression was first significantly altered 3 dpi that remained significantly modified at all later time points examined. These 156 genes were significantly enriched in canonical pathways associated with interferon (IFN) signaling, activation of IFN elements by intracellular pattern recognition receptors, and antiviral immunity induced by cytosolic RIG-like receptors. Evaluation of this gene set with the National Center for Biotechnology Information Gene and INTERFEROME databases corroborated pathway analysis, as function of most (53%) were linked to IFN-mediated host immunity. In the final set of experiments, ivag administration of the Toll-like receptor 3 agonist polyinosinic: polycytidylic acid (poly I:C) 24 h before ivag HSV-2 infection reduced the incidence of genital pathology and encephalopathy, while these poly I:C-treated mice were subsequently protected from ocular HSV-2 challenge lethal to uninfected controls. The latter results imply that the exuberant antiviral immunity produced in our experimental model is simply formed too late to prevent viral replication and dissemination, and that poly I:C-induced formation of an antiviral state protecting against primary ivag infection also permits development of HSV-specific protective immunity.
Introduction
Providing further evidence for the significance of IFN-mediated antiviral immunity, mice lacking the IFNα/βR demonstrate enhanced susceptibility to ivag HSV-2 infection (13). After ivag HSV-2 infection, these IFNα/βR-deficient mice also display impaired NK cell responses, higher viral load, and an accelerated progression of disease (4,10). Conversely, ivag administration of the Toll-like receptor 3 (TLR3) agonist, polyinosinic: polycytidylic acid (poly I:C), to WT mice prior to or concomitant with ivag HSV-2 infection averts tissue damage and encephalopathic changes; improved outcomes attributed to poly I:C-mediated increases in IFN-β production (1,5). In the current study, we sought to enhance understanding of IFN-mediated antiviral immunity and other early host responses to viral invasion of mucosal tissue using oligonucleotide microarrays that compared gene expression in vaginal tissue from uninfected mice and mice 1, 2, 3, 4, 5, 6, or 7 days after primary ivag HSV-2 infection.
Material and Methods
Mice and ivag infection
Conducted experiments were described in a protocol approved by the University of Pittsburgh's Institutional Animal Care and Use Committee. Six-week-old WT C57BL/6 female mice from The Jackson Laboratory (Bar Harbor, ME) were subcutaneously administered 0.7 mg depot-medroxyprogesterone acetate (DMPA) (Upjohn, Don Mills, Ontario, Canada). 5 days later, mice were sedated via intraperitoneal injection of 1.8 mg ketamine hydrochloride (Fort Dodge Animal Health, Fort Dodge, IA) and 0.18 mg xylazine (Lloyd Laboratories, Shenandoah, IA), and intravaginally infected with 104 pfu WT HSV-2 333.
Genital pathology and ocular infection
Genital pathology produced by ivag HSV-2 infection was scored daily using a 5-point scale: 0, no pathology; 1, mild vulvar erythema; 2, moderate vulvar erythema and swelling; 3, sever vulvar erythema and swelling and perineal fur loss; 4, perineal ulceration; 5, extension of perineal ulceration and fur loss to surrounding tissue (8). Mice were euthanized when stage 4 pathology or greater was seen. Where indicated, 24 h before ivag HSV-2 infection mice were intravaginally administered 100 μg poly I:C (InvivoGen, San Diego, CA) suspended in 10 μL of endotoxin-free physiological water. Twenty-one days later, age-matched, uninfected controls and the mice previously rescued from ivag HSV-2 infection by poly I:C treatment were sedated, and 2.5 x 103 pfu WT HSV-2 333 topically applied bilaterally to abraded corneas. Kaplan-Meier survival curves and logrank tests compared cumulative survival incidence after primary ivag HSV-2 infection of untreated, uninfected controls versus poly I:C-treated mice and after ocular HSV-2 challenge of uninfected controls versus mice previously rescued from HSV-2 ivag infection by antecedent poly I:C- treatment.
Quantification of HSV copy number
As indicated, 2 days post infection (dpi), cervicovaginal lavage (CVL) samples were collected to quantify HSV-2 genome copy number. For this assay, 30 μL of PBS was inserted into individual vaginal vaults, recovered, and then combined with 70 μL PBS. Total DNA was isolated using DNeasy columns (Qiagen, Valencia, CA), and 5 μL of extracted DNA added to a master mix containing: 200 nmol/L each of two HSV-2 DNA polymerase gene–specific primers (3); 200 nmol/L each of a hybridization probe-quencher oligo pair; 2% dimethyl sulfoxide; 5 mM MCl2; 0.5 units uracil-DNA glycosylase; and Fast-Start DNA Master HybProbe master mix (Roche Applied Sciences, Madison, WI). After incubating at 40°C for 10 min and 95°C for 10 min, samples were amplified 50 cycles at 95°C (0 sec), 60°C (5 sec), and 72°C (10 sec) with a Light Cycler 1.2 (Roche Applied Sciences). To quantify amplification, fluorescence was measured at the end of each cycle's annealing step (60°C). Control samples (Advanced Biotechnologies, Columbia, MD) containing 5000, 500, 50, and 5 copies of HSV-2 genomic DNA were used to create standard curves, and 5-copy standards showed positive amplification with each assay run. HSV-2 DNA copy number in CVL samples from poly I:C-treated or untreated mice were compared using the unpaired Mann-Whitney U test.
Vaginal processing for microarrays
DMPA-treated, uninfected mice and DMPA-treated, HSV-infected mice (1, 2, 3, 4, 5, 6, or 7 dpi) were anesthetized and euthanized by cardiac perfusion of sterile PBS. Whole vaginas were excised and placed into vials containing 0.8 mL RNeasy solution (Qiagen). Vaginas were mechanically disrupted using a ball mill (Retsch Inc., Irvine, CA), and total RNA isolated in accordance with manufacturer's instructions (Qiagen). Thereafter, cDNA synthesis, labeling, hybridization, and scanning processes were done using protocols provided by the manufacturer (Affymetrix, Santa Clara, CA).
Microarray analysis
Vaginas from 5 DMPA-treated, uninfected mice were used to establish baseline gene expression, while three independent experimental series provided 14 HSV-infected vaginal samples. Vaginas from infected mice were collected each day 1–7 dpi (series A), 2–5 dpi (series B), and 1–3 dpi (series C), inclusive. Thus, duplicate or triplicate specimens were available for dpi 1–5, while the individual dpi 6 and dpi 7 samples were combined to provide a single gene expression value (identified in manuscript as 6 dpi). Raw microarray data was processed using Affymetrix GCOS 1.4 software and viewed using Excel (Microsoft, Richmond WA).
For gene expression analysis, the earliest day expression was significantly altered by infection was assessed using Dunn's test (α=0.05). Determination that individual gene expression, at any particular dpi, was significantly altered by ivag HSV-2 infection required satisfaction of the following criteria: 1) Pairwise comparison between an infected sample and the five uninfected samples identified at least four significant changes; 2) The higher expressing group contained detectable transcript in each sample from an individual infected group (1–6 dpi) or at least 4 of 5 uninfected group samples; 3) No overlap of expression values between an infected (1–6 dpi) and uninfected group; 4) Mean infected expression values for an infected group (1–6 dpi) differed at least 2-fold from mean uninfected values.
To evaluate gene expression at each individual dpi, controls were constructed with pooled data from uninfected mice and infected mice at earlier dpi. For example, to evaluate expression at dpi 3, control data was comprised of uninfected mice and mice dpi 1 and 2. Sequential changes in expression (e.g., dpi 2 vs. dpi 3) were then determined using GCOS software. Data from the genes first significantly altered 3 dpi, that remained altered 4, 5, and 6 dpi, was imported into GeneSpring GX Version (Agilent Technologies Inc., Santa Clara, CA) software, and Ingenuity Pathway Analysis used to identify the canonical pathways significantly enriched in this set of early responding genes. The National Center for Biotechnology Information (NCBI) Gene and INTERFEROME (
Results
To characterize early immune responses elicited upon HSV-2 invasion of lower genital tract mucosa, oligonucleotide microarrays were performed with vaginal tissue from uninfected mice and from mice 1–7 days after ivag infection. Initial analysis was used to organize results based on the earliest dpi that individual gene expression was significantly altered by infection and the total number of consecutive days that expression of this gene remained altered (expression in infected samples was classified as significantly altered relative to uninfected samples when all four selection criteria described in Materials and Methods were satisfied) (Fig. 1A). As demonstrated in Figure 1A, transcriptional profiles in the first 2 days of infection were similar to those found in uninfected vaginas. As also displayed in this figure, we identified 156 genes whose expression became significantly altered between 2–3 dpi and that remained significantly altered 4, 5, and 6 dpi. Figure 1B provides graphic representation of expression patterns for this 156-gene group, revealing that many of these genes remained dramatically upregulated for the duration of dpi examined. Assessing biological function of this same group of 156 genes by pathway analysis, we next determined that the two most significantly enriched canonical pathways were populated with genes associated with IFN signaling and the activation of IFN response factors (IRFs) by intracellular pattern recognition receptors (Table 1). Among additional canonical pathways more enriched in this 156-gene group compared to other gene groups shown in Figure 1A were ones associated with OX40 signaling, protein ubiquitination, and the activation of RIG-like receptors (intracellular pattern recognition receptors known to stimulate type I IFN production upon identification of viral nucleic acid) (5) (Table 1).
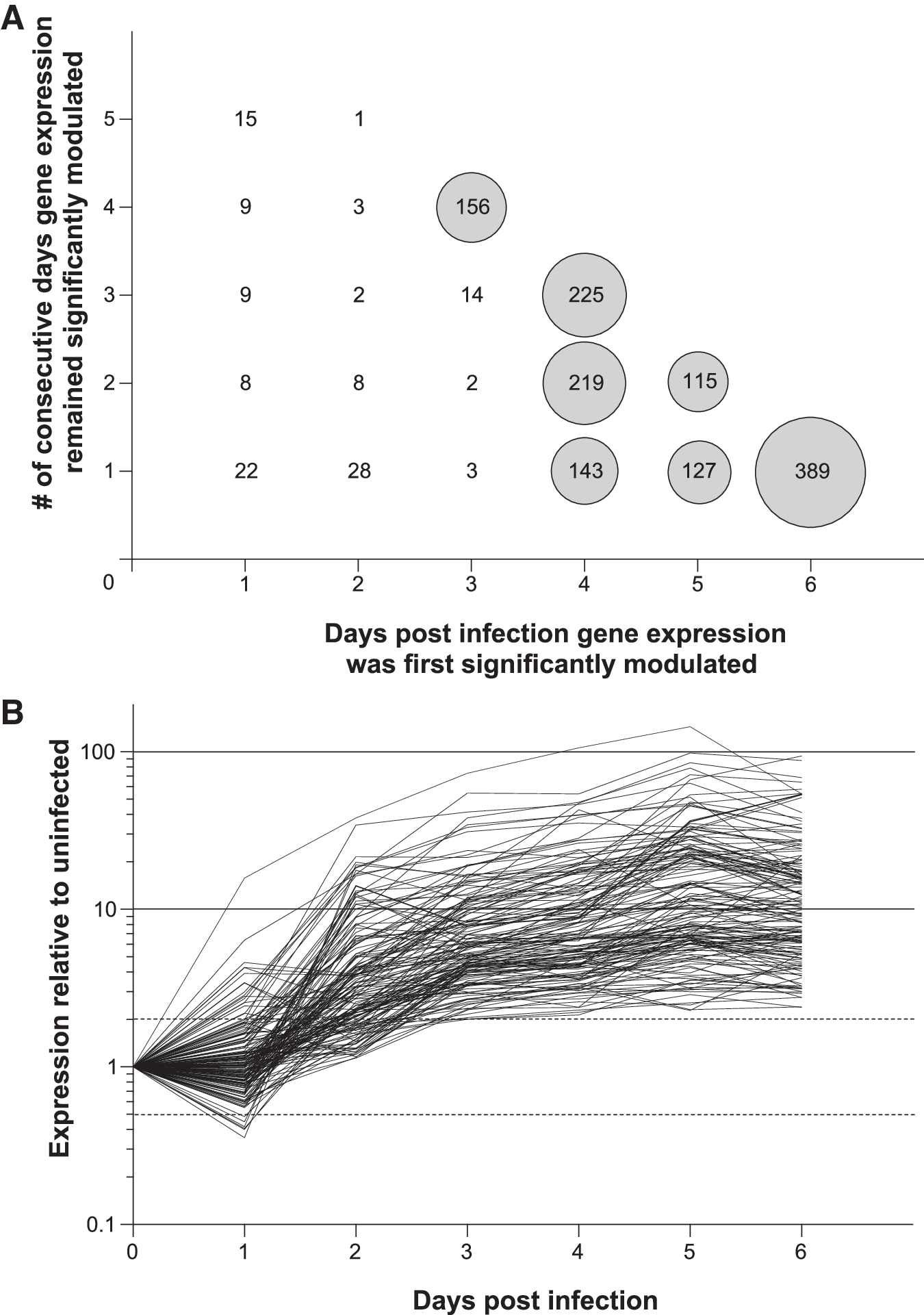
First substantial modulation of gene expression in HSV-infected vaginal tissue occurred between 2–3 days post infection (dpi).
Significance expressed as −log (p) values).
We next scrutinized biological activity of the genes responding earliest to HSV-2 ivag infection using the NCBI Gene and INTERFEROME databases. The INTERFEROME database consists of all genes with evidence of INF-mediated regulation, and has been previously used to assess IFN-mediated anti-viral host responses (18). In this database, there are currently 281 genes appearing in more than one report and 1564 genes appearing in an individual report. For our analysis, we determined there was definitive link to IFN-mediated immunity when a gene was listed in the NCBI Gene database as IFN-induced or IFN-inducible or when in a gene appeared in multiple INTERFEROME reports, while a less certain IFN connection was determined if a gene appeared in only one INTERFEROME report. Among the 156 genes whose expression was first significantly altered 3 dpi, and that remained significantly altered 4–6 dpi (Fig. 1), 33.3% (52/156) were seen in multiple INTERFEROME reports or identified as either IFN-induced or IFN-inducible in the NCBI Gene database, and thus strongly linked to IFN-mediated immunity (Fig. 2). Table 2 provides additional information on these 52 genes by cataloging their individual names and fold expression increases (vs. uninfected tissue). In addition to these, another 31 (19.9%) of the 156 early-responding genes were located in a single INTERFEROME report, providing further indication of the robust IFN-mediated immunity elicited by HSV-2 ivag infection.
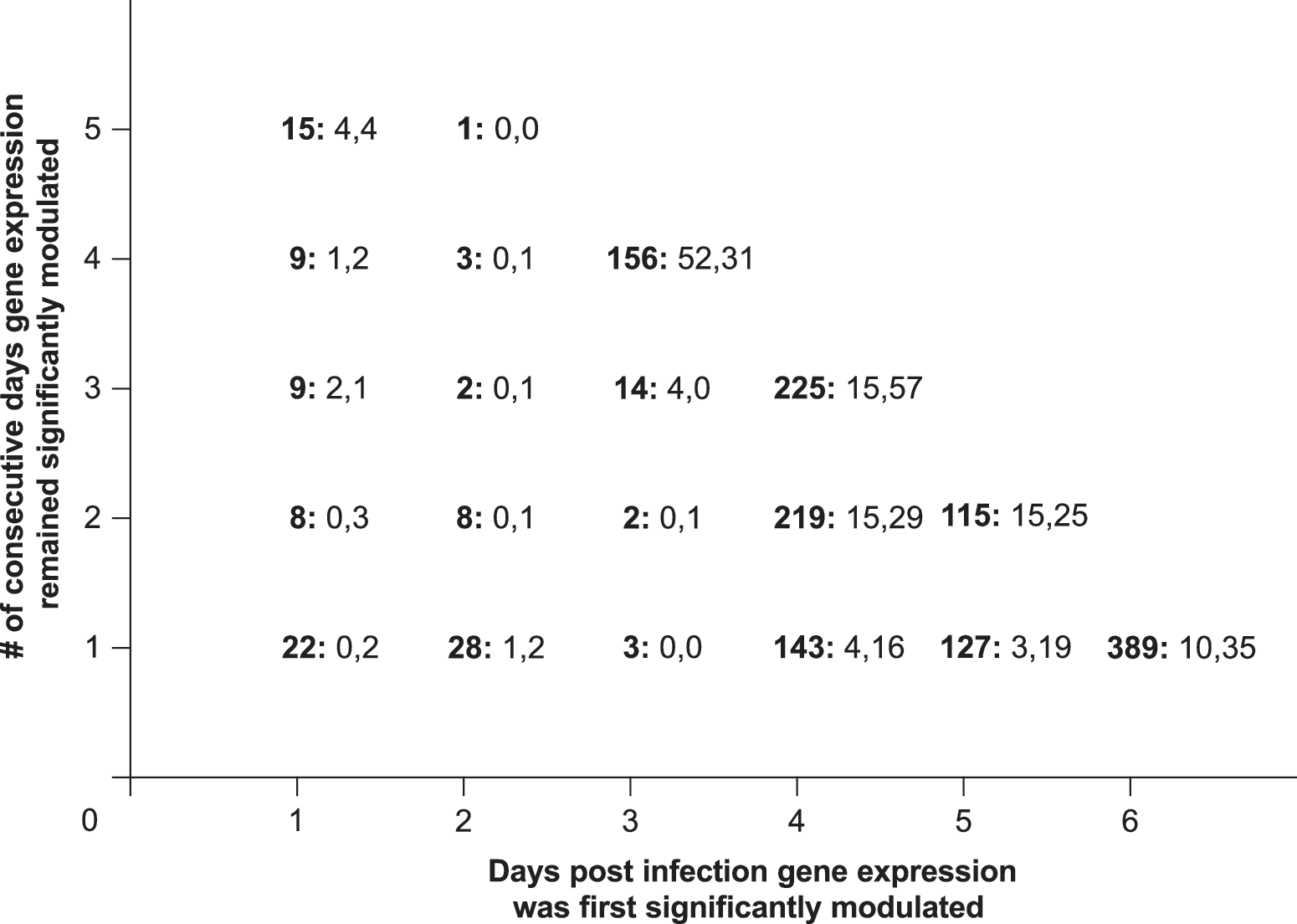
IFN-mediated immunity dominated the early response to murine HSV-2 ivag infection. Depiction of gene sets displayed in Figure 1A by IFN-relatedness (data for each gene set is presented using the following convention: total gene number: number of genes with definitive link to IFN-mediated immunity, number of genes with possible link to IFN-mediated immunity). Definitive link to IFN-mediated immunity was decided when a significantly modulated gene was either listed in the National Center for Biotechnology Information Gene database as IFN-induced or IFN-inducible or found in more than one report of the INTERFEROME database. Possible linkage to IFN-mediated immunity was determined when a gene appeared in only one INTERFEROME report. Thus among the set of 156 early-responding genes whose expression is depicted in Figure 1B, 52 (33.3%) were strongly linked to IFN-mediated immunity, and another 31 (19.9%) had possible association.
An additional expressed sequence (AW112010), identified by the NCBI Gene database as IFN-induced, showed a 3-fold increase in expression.
While our microarray analyses showed ivag infection of mice with 104 pfu WT HSV-2 elicits exuberant antiviral immunity, previous studies found this response was unable to prevent mice from developing fatal encephalopathy (11,14,16). Conversely, earlier work also revealed that ivag administration of poly I:C to C57BL/6 mice before or concomitant with HSV-2 infection prevents genital pathology and encephalopathy (1,5). We thus hypothesized that poly I:C treatment of mice prior to HSV-2 ivag infection lowers incidence of HSV-associated morbidity by suppressing primary replication of virus. We tested this hypothesis by intravaginally administering 100 μg of poly I:C to mice 24 h before HSV-2 ivag infection, and saw that, compared to untreated controls, treated mice had less local tissue damage and enhanced rates of survival (Fig. 3A, B). Poly I:C treatment also typically reduced, but did not eliminate, HSV-2 DNA copy number measured in CVL specimens collected 48 h after ivag infection (Fig. 3C). Moreover, poly I:C-treated mice (n=24) not succumbing to primary HSV-2 ivag infection were also more likely to survive ocular challenge with a HSV-2 dose lethal to uninfected controls (Fig. 3D). Among the 24 poly I:C-treated mice protected from ivag infection, all 16 that had detectable HSV-2 DNA levels (> 5 copies) in CVL specimens collected 2 dpi (displayed in Fig. 3C) also survived ocular challenge, compared to only 25% (2/8) that had undetectable levels of virus (Fig. 3E). This implied that poly I:C-treatment prevented development of genital pathology in the presence of a primary infection that evoked formation of protective HSV-specific adaptive immunity.
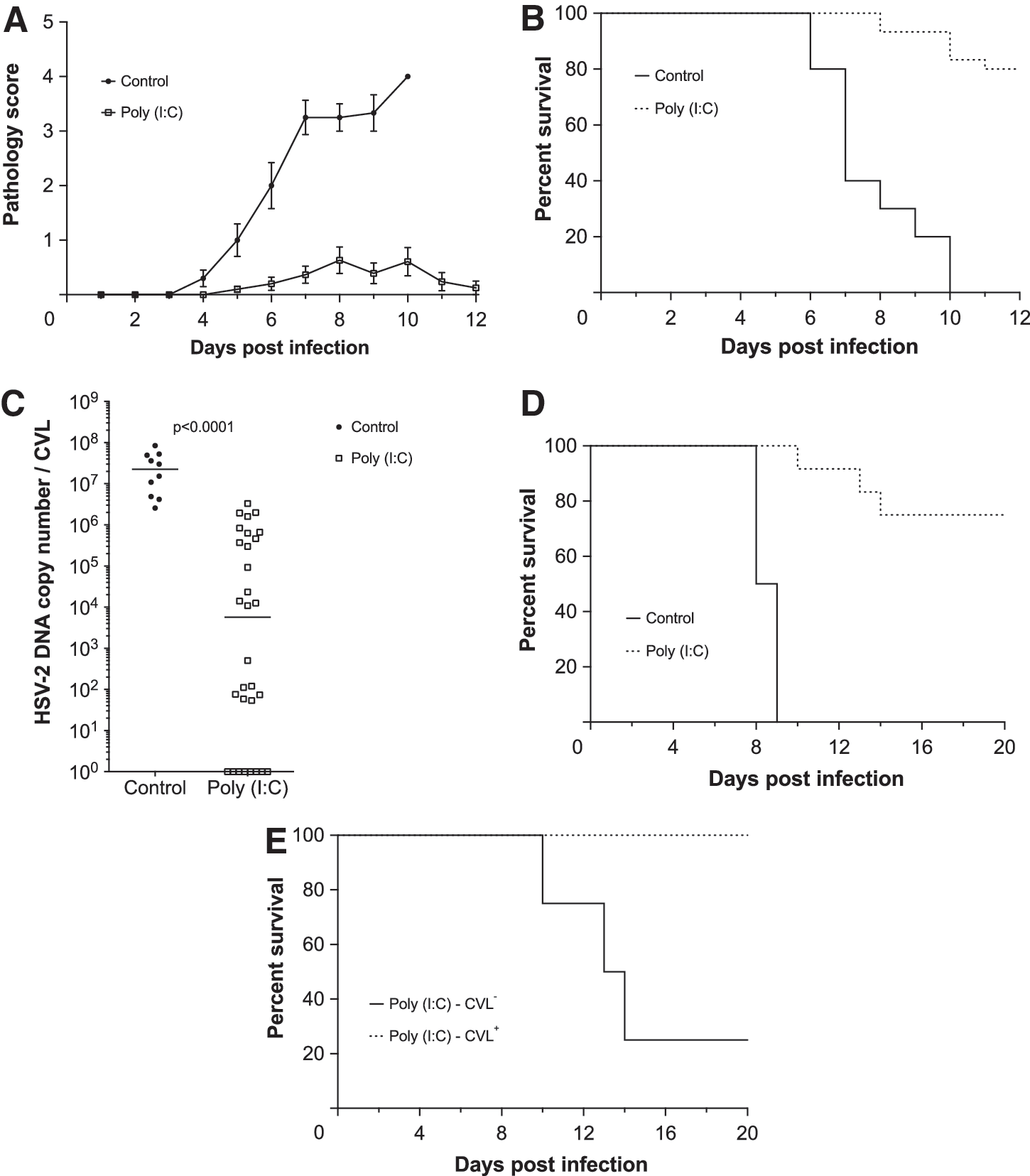
Intravaginal polyinosinic:polycytidylic acid (poly I:C) administration 1 day prior to ivag HSV-2 infection limited viral replication, reduced incidence of encephalopathy, and allowed development of HSV-specific protective immunity. Untreated, uninfected controls or uninfected mice that had been treated with 100 μg of poly I:C 24 h earlier were intravaginally infected with 104 pfu HSV-2. Mice were then monitored daily for
Discussion
Our comparative analysis of vaginal oligonucleotide microarray data from uninfected mice and mice 1–7 days after HSV-2 ivag infection elucidates several features of the immune response elicited in HSV-2-infected vaginal tissue. Specifically, ivag infection with 104 pfu WT HSV-2 produces limited number of vaginal transcriptome changes that are detectable by oligonucleotide microarray in the first 48 h after infection. Conversely, analysis of gene expression that was significantly altered in the next 24 h of infection reveals that the recognition of HSV-2 nucleic acid by cytosolic TLRs and RLRs promotes development of an exuberant host response thoroughly dominated by IFN-mediated antiviral immunity. This conclusion was further supported by interrogation of microarray data with the NCBI Gene and INTERFEROME databases, analyses which indicate approximately half of the 156 genes whose expression was first altered between 2–3 dpi (and whose expression remained altered through 6 dpi) are linked to IFN-mediated antiviral immunity. Together, such findings suggest that although mice generate exuberant antiviral immunity against HSV-2 ivag infection, this response is generated too late to adequately control primary infection or prevent dissemination of the virus to sacral ganglia and the central nervous system.
This finding may be related to strategies evolved by HSV to escape IFN-mediated innate defense systems with viral products (e.g., ICP0, ICP27, ICP34.5, and vhs) (12). ICP0 inhibits activation of IFN-stimulated genes by blocking nuclear translocation of IRF3 (9), ICP27 inhibits IFN-induced STAT1 phosphorylation and nuclear translocation (7), ICP34.5 inhibits IRF3 activation via interaction with TANK-binding kinase (21), and vhs inhibits JAK/STAT signaling, IRF7 expression, and IFN-α production (22). Consistent with these immunoevasive strategies of HSV-2, transcriptional analysis of lesional tissue from individuals that were experiencing episodes of HSV-2 reactivation were remarkable for the absence of type I IFN induction (17). As low HSV-2 DNA copy numbers were also detected in those lesions (17), the exuberant (albeit delayed) type I IFN immune responses induced in our studies may have been stimulated by the relatively larger inoculums of HSV-2 that were utilized in our experimental model of infection.
Though the mice in our studies did not generate sufficiently robust IFN-mediated host defenses quickly enough to permit establishment of the nonlethal, latent HSV-2 infections characteristic of infection in the genital tract of immunocompetent adults, our results do support further use of this model to delineate early host immunity to invasive viral pathogens and to explore specific components of this response that optimize virus-specific adaptive immunity. In findings corroborating this conclusion, we found that mice intravaginally administered poly I:C prior to HSV-2 ivag infection were spared development of genital pathology and encephalopathy. This implies that poly I:C, a potent inducer of Type I IFN production, suppresses HSV-2 replication by accelerating activation of an antiviral state. While earlier studies demonstrated that poly I:C protected mice against HSV-2 ivag infection (11,16), our study is the first to indicate that the antiviral state stimulated by this TLR-3 agonist confer protections despite detection of 106 HSV-2 DNA copies in CVL of poly I:C-treated mice (Fig. 3C). Further evidence that poly I:C treatment suppresses, but does not prevent, a productive primary infection was provided by experiments in which poly I:C-rescued mice were spared encephalopathic changes when ocularly challenged with HSV-2 doses lethal for uninfected controls (Fig. 3D, E). This indicates that further exploration of this experimental model may elucidate immune responses that prevent HSV-2 acquisition or inhibit spread of the virus from epithelial to nervous system tissue.
Footnotes
Acknowledgments
This work was supported by grants from the National Institutes of Health (R56AI085110 and P30EY008098) and the Pennsylvania Department of Health (the latter claims no responsibility for analyses, interpretations, and conclusions contained in this article).
Disclosure Statement
Authors declare no competing financial interests.