
Abstract
Dengue virus (DENV) infection induces a strong B-cell immune response against the viral nonstructural protein 1 (NS1). Anti-NS1 antibodies (Abs) may affect virus production because they coexist with the virus in the patients' blood. The present study examined whether ten mouse monoclonal antibodies (MAbs) raised against NS1 affected production of the DENV-2. Three MAbs, 4C2, 4G11, and 4E5, showed weak neutralizing activity in a focus reduction assay. In addition, two serotype-specific MAbs, 4C2 and 4G11, protected suckling mice from lethal infection with DENV-2. An immunoprecipitation assay with DENV-2 showed that these MAbs, which were specific for the NS1 of DENV-4 and DENV-1, cross-reacted with the DENV-2 pre-membrane (prM) protein, but not with DENV-2 NS1. Interestingly, high concentrations of MAb 4G11 showed antibody-dependent enhancement of DENV-2 infection in human monocyte THP-1 cells. Taken together, these observations suggest that serotype-specific anti-NS1 MAbs are potentially involved in virus production.
Introduction
The DENV genome contains three structural proteins (capsid, premembrane [prM], and envelope [E]) and seven nonstructural proteins (NS1, NS2a, NS2b, NS3, NS4a, NS4b, and NS5). During the course of DENV infection, the virus induces production of Abs that mainly target the prM, E, and NS1 (39). Abs raised against DENV structural proteins play a crucial role in controlling virus production via neutralization or enhancement (9,40). Indeed, several studies report that most protective Abs bind to the E protein expressed by the virions; by contrast, anti-prM Abs are likely to increase the risk of subsequent infection through ADE (7,8).
Forty years ago, NS1 was identified as a DENV-soluble complement-fixing antigen (3). Today, the NS1 and anti-NS1 Abs are thought to be involved in pathogenesis. A unique feature of DENV is that NS1 is secreted by infected cells during the acute phase and circulates in the blood at high concentrations (1,18,26,43). Thus, a strong Ab response is generated against NS1, and this is more often observed during secondary infections (19,39). However, the roles of NS1 and anti-NS1 Abs in regulating DENV production are unclear. Passive administration of Abs against flavivirus NS1 protects mice from lethal infection by flaviviruses (17,35). The protective effect of anti-NS1 Abs may be due, in part, to cross-reactivity with the E protein, as treatment with anti-NS1 Abs showed neutralizing ability to some extent (11). Anti-NS1 Abs also play a role in DENV disease progression by cross-reacting with host molecules (4,10,27). These observations suggest that Ab responses to NS1 play an important role in disease progression.
We previously generated ten mouse MAbs specific for DENV-NS1 (28). The present study examined the neutralizing activity of these MAbs against four DENV serotypes and their ability to protect suckling mice from lethal DENV infection. We also determined the cross-reactivity of the MAbs with DENV structural proteins, identified their target epitopes using recombinant NS1 proteins, and examined their ADE activity. The results of this study provide novel insights into the involvement of anti-NS1 Abs in DENV virus production.
Materials and Methods
Cells and viruses
Hybridomas and human monocyte THP-1 cells were cultured in RPMI 1640 (NACALAI TESQUE, Kyoto, Japan) supplemented with 10% fetal calf serum (FCS). The 293T human renal epithelial cell line and the BHK-21 Syrian hamster fibroblast cell line were cultured in Dulbecco's modified Eagle medium (DMEM) (Gibco, Grand Island, NY) supplemented with 10% FCS. African green monkey kidney cells (Vero) were grown in Eagle's minimum essential medium (NACALAI TESQUE) supplemented with 10% FCS. The C6/36 cell line was cultured at 28°C in L-15 medium (Gibco) supplemented with 10% FCS and 2.9% tryptose phosphate broth (TPB) (Sigma, St Louis, MO). All cell lines (except C6/36) were cultured at 37°C in an atmosphere containing 5% CO2. DENV serotypes 1 (Mochizuki strain), 2 (16681 strain), 3 (H87 strain), and 4 (H241 strain) were propagated in C6/36 cells for 7–9 days, or in BHK cells for 3 days, in maintenance medium (L-15 medium containing 2% FCS and 2.9% TPB for C6/36 cells, and DMEM containing 2% FCS for BHK-21 cells). Viruses were stored at −80°C before use.
Production and purification of MAbs from mouse ascitic fluid
Desirable clones were expanded in pristine-primed mice and the MAbs were purified from the ascitic fluid using protein G Sepharose™ 4 Fast Flow (GE Healthcare, Uppsala, Sweden). MAb concentrations were determined using the Bradford Protein Assay (Bio-Rad Laboratories, Hercules, CA).
Plasmid construction and protein expression
To construct the plasmids expressing DENV NS1 proteins, viral RNA was extracted using a QIAmp Viral RNA Mini kit (Qiagen, Valencia, CA, USA) according to the manufacturer's instructions, and reverse transcription was performed using Superscript III (Invitrogen) and random primers. The resulting cDNAs were amplified by PCR using a set of primers (Table 1). The PCR products were inserted into the pFLAG-CMV™-3 expression vector (Sigma) via the EcoRV/BamH1 restriction sites and then transformed into E. coli XL1-blue. Extracted plasmids were subjected to sequencing analysis. To express the NS1 proteins, the plasmids were transfected into 293T cells using Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. The cells were harvested 2 days later by treatment with PBS containing 0.5 mM EDTA. Immunofluorescence assays and Western blot analysis were performed with an anti-Flag M2 Ab (Sigma).
The sites recognized by the restriction enzymes are underlined.
Immunofluorescence assay
Cells were fixed with PBS containing 4% paraformaldehyde (PFA), permeabilized with PBS containing 1% Triton X-100 for 5 min at room temperature, and then incubated with culture fluid harvested from the hybridomas (4C2 or 4G11) for 1 h. The cells were washed three times with PBS and then incubated with Alexa 488-goat anti-mouse IgG Ab (Invitrogen, Paisley, UK) for 30 min at room temperature. After washing three times with PBS, the cells were observed under a fluorescence microscope. Human plasma derived from patients infected with DENV (HPD), namely dengue immune individuals, was used as a positive control (28).
SDS-PAGE and Western blotting
DENV-infected BHK-21 cells were lysed in SDS-PAGE sample buffer and heated at 100°C for 5 min. The resulting proteins were separated by 12% sodium-dodecyl-sulfate polyacrylamide electrophoresis (SDS-PAGE) and transferred to PVDF membranes (Millipore Corporation, Bedford, MA). The membranes were incubated overnight with MAbs generated by the hybridoma clones, and then incubated with horseradish peroxidase (HRP)-conjugated anti-mouse IgG (Jackson ImmunoResearch Laboratories, West Grove, PA) or anti-human IgG (Jackson ImmunoResearch Laboratories) for 1 h. Reactive viral proteins were detected using the ECL Western blotting detection reagent (GE Healthcare, Buckinghamshire, UK).
In vitro neutralization test
Each MAb or mouse normal IgG (2 mg/mL), was serially diluted (4-fold) and reacted with the different DENVs serotypes (100 ffu/well). The final concentration of MAb in each reaction was 500, 125, 31.25, 7.81, 1.95, 0.50, 0.12, and 0.03 μg/mL. MAbs were inoculated with DENVs (100 ffu) at 37°C for 30 min. The DENV-MAb complexes were then inoculated onto a monolayer culture of Vero cells and incubated for 2 h. Overlay medium was then added, followed by a further incubation at 37°C for 24 h. The cells were fixed with PBS containing 4% PFA and viral foci were visualized by staining with an anti-DENV Ab (2H2; ATCC) for 1 h, followed by incubation with Alexa 488-goat anti-mouse IgG Ab (Invitrogen). The neutralizing activity of the MAbs was expressed as a 50% reduction in DENV infection in the focus reduction neutralizing test (FRNT50).
Mouse protection assay
Pregnant BALB/c mice were purchased from Japan SLC, Inc (Shizuoka, Japan). All animals were cared for and manipulated according to the animal experiment guidelines of Osaka University. Each of the MAbs (20 μg) was mixed with DENV-2 (400 ffu) (MAb-virus mixture) and incubated for 15 min. Two-day-old suckling mice were inoculated intracerebrally with 400 ffu/mouse of DENV in the presence/absence of 20 μg MAbs (n=16–21 per group). All infected mice were observed for more than 2 weeks, and morbidity or mortality was recorded daily.
Immunoprecipitation
At 72 h post-infection, BHK-21 cells were harvested in 1 mL of PBS and centrifuged at 900 g for 5 min. The precipitated cells were lysed in 1 mL of lysis buffer (1% [vol/vol] NP-40, 10 mM Tris-HCl [pH 7.4], 150 mM NaCl, 2 mM EDTA, and 0.1% protease inhibitors) for 1 h at 4°C and centrifuged at 900 g for 10 min to remove debris. The supernatants were centrifuged at 10,000 g for 20 min and the resulting supernatants were incubated with 1 μg or 5 μg of MAbs 4C2, 4G11, 4G2 (anti-E ), 6D6 (anti-prM ), 1A5 (anti-NS1) (28) for 1 h at 4°C, followed by immunoprecipitation with Protein G Sepharose (GE Healthcare, Piscataway, NJ) for 1 h at 4°C. The beads were then washed with lysis buffer and the precipitated proteins were analyzed by SDS-PAGE, followed by immunoblotting with HPD.
Antibody-dependent enhancement (ADE) assay
Differentiation of human monocytes (THP-1 cells) was induced by culture in RPMI supplemented with 10% FCS plus 34.2 mM phorbol-12-myristate-13-acetate. Serially diluted MAb (diluted 10-fold from 5 mg/mL to 5×10−4 μg/mL) was inoculated with DENV (3.75×103 ffu) at 37°C for 30 min. The differentiated cells were inoculated with a MAb-virus mixture (at a multiplicity of infection (MOI) of 0.5) and cultured for 3 days. Viral RNA was isolated from the culture supernatant using the QIAmp Viral RNA Mini kit (Qiagen) according to manufacturer's protocol and quantified using a QuantiTect SYBR Green PCR Kit (Qiagen) with the following set of dengue group-specific primers: DENV-F, 5′-CAATATGCTGAAACGCGAGAGAAA-3′ and DENV-R, 5′-CCCCATCTATTCAGAATCCCTGCT-3′ (36). Primers specific for glyceraldehyde 3-phosphate dehydrogenase (GAPDH) RNA (GAPDH-F, 5′- ACCACAGTCCATGCCATCAC-3′ and GAPDH-R, 5′- TCCACCACCCTGTTGCTGTA-3′) were used as the control. The reaction conditions were: 50°C for 30 min, followed by 95°C for 15 min, 40 cycles of 95°C for 20 sec, 55°C for 30 sec, and 72°C for 30 sec, and a melting curve analysis step using the ABI PRISM 7900HT Real Time PCR System (Applied Biosystems, Foster City, CA). Normalization was performed against GAPDH using the delta-delta Ct method.
Statistical analysis
Data are presented as mean±SEM. Statistical analyses were performed using Prism version 5.0 (GraphPad Software). Statistical significance was determined by log-rank test to compare survival curves. P values less than 0.05 was considered statistically significant.
Results
Anti-NS1 MAbs show weak neutralizing activity against DENV
To examine whether the MAbs had neutralizing activity, we performed an in vitro neutralization test against DENV-2. Of the 10 MAbs tested, 4C2, 4G11, and 4E5 showed neutralizing activity, although it was relatively weak compared with that of the anti-E 4G2 MAb (Table 2). We then examined the neutralizing activity of 4C2, 4G11, and 4E5 against the four DENV serotypes. MAb 4C2 showed moderate neutralizing activity against DENV-1 and -2, but only weak activity against DENV-3 and -4 (FRNT50=87.4, 64.4, 391.9, and 165.5 μg/mL against DENV-1, -2, -3, and -4, respectively; Table 3). MAb 4E5 showed moderate neutralizing activity against DENV-1, -2, and -4 (FRNT50=38.5, 218.6, and 206.6 μg/mL, respectively), but did not neutralize DENV-3. MAb 4G11 neutralized DENV-1 and -2 (FRNT50=60.8, and 82.5 μg/mL, respectively), but not DENV-3 or -4 (Table 3). These results indicate that the neutralizing activity of the anti-NS1 MAbs was serotype dependent.
The number of plus indicates the degree of neutralization activity. Three plus (+++), two plus (++), one plus (+), and one minus (−) indicate FRNT50≤10 μg/mL, 10≤FRNT50≤100 μg/mL, 100≤FRNT50≤100 μg/mL, and FRNT50≥500 μg/mL, respectively.
The mean value is presented on the left of each cell, with the standard deviation on the right of the cell.
Anti-NS1 MAbs protect suckling mice from lethal DENV infection
We next assessed whether MAbs 4C2, 4E5, and 4G11 protected suckling mice from lethal infection with DENV-2. The survival rate of control mice treated with anti-E MAb 4G2 (20 μg) after infection with DENV-2 was 87.5%, whereas that of mice treated with MAbs 4C2 and 4G11 was 19.05% and 14.29%, respectively (Fig. 1). However, MAb 4E5 did not protect suckling mice against DENV-2 infection at any concentration (Fig. 1), possibly due to its weak neutralizing activity (Table 3). It is likely that MAbs 4C2 and 4G11 neutralize the DENV-2 virus in vivo, thereby protecting the mice from lethal infection.
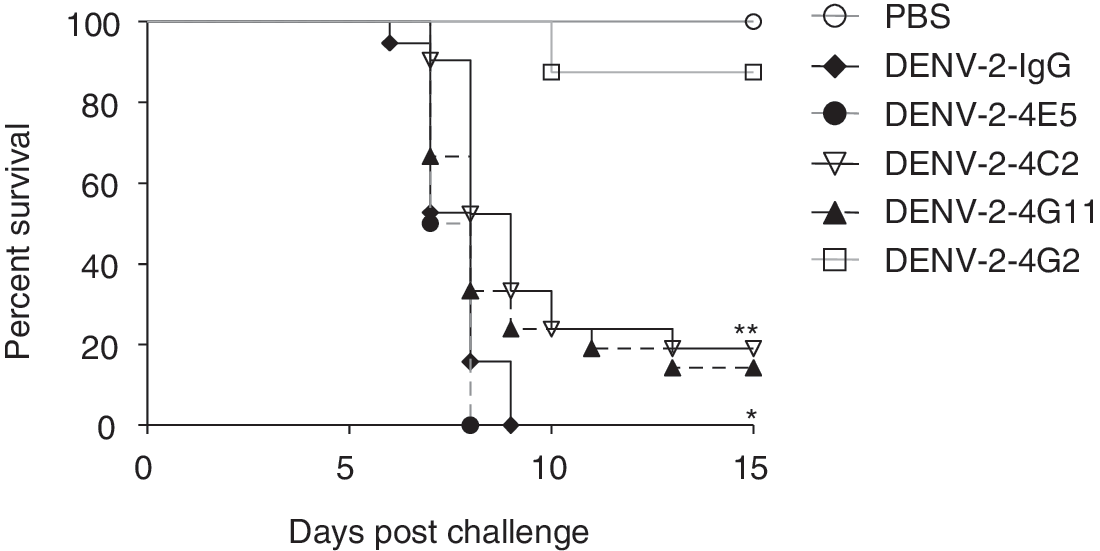
Survival of BALB/c suckling mice challenged with DENV-2. A mixture containing virus and MAbs 4C2 or 4G11 was injected into the cerebra of suckling mice. Mouse IgG and a neutralizing anti-E MAb (4G2) were used as negative and positive controls, respectively. Kaplan-Meier survival curves show the percentage of mice surviving after treatment or nontreatment with the antibodies. **p=0.0006, *p=0.0482 (versus IgG).
MAbs 4C2 and 4G11 react with DENV-4 NS1 and DENV-1 NS1 but not with DENV-2 NS1
It was strange to observe the neutralizing and protective activities of MAbs 4C2 and 4G11 to DENV-2 (Table 2 and Fig. 1) because Western blot analyses against DENV-infected cell lysates showed that MAbs 4C2 and 4G11 are serotype-specific (for DENV-1 and DENV-4 NS1, respectively) (28). To examine the reactivity of these MAbs against NS1 from different DENVs serotypes, we expressed recombinant NS1 proteins in 293T cells and examined their reaction with the MAbs by immunofluorescence analysis. MAbs 4C2, 4G11, and 4E5 reacted with the recombinant NS1 proteins, and they were either serotype-specific or partially cross-reactive (Fig. 2). Importantly, neither MAb 4C2 nor MAb 4G11 reacted with DENV-2 NS1; rather, MAb 4C2 reacted with DENV-4 NS1 and MAb 4G11 reacted with DENV-1 NS1. This was unexpected, because both 4C2 and 4G11 neutralized DENV-2 and protected mice from lethal infection with DENV-2 (Table 3 and Fig. 1). Therefore, we examined the protective effects of MAbs 4C2 and 4G11 against infection by DENV-1 and DENV-4 respectively; however, neither MAb protected the mice (Supplementary Data 1; Supplementary material is available online at
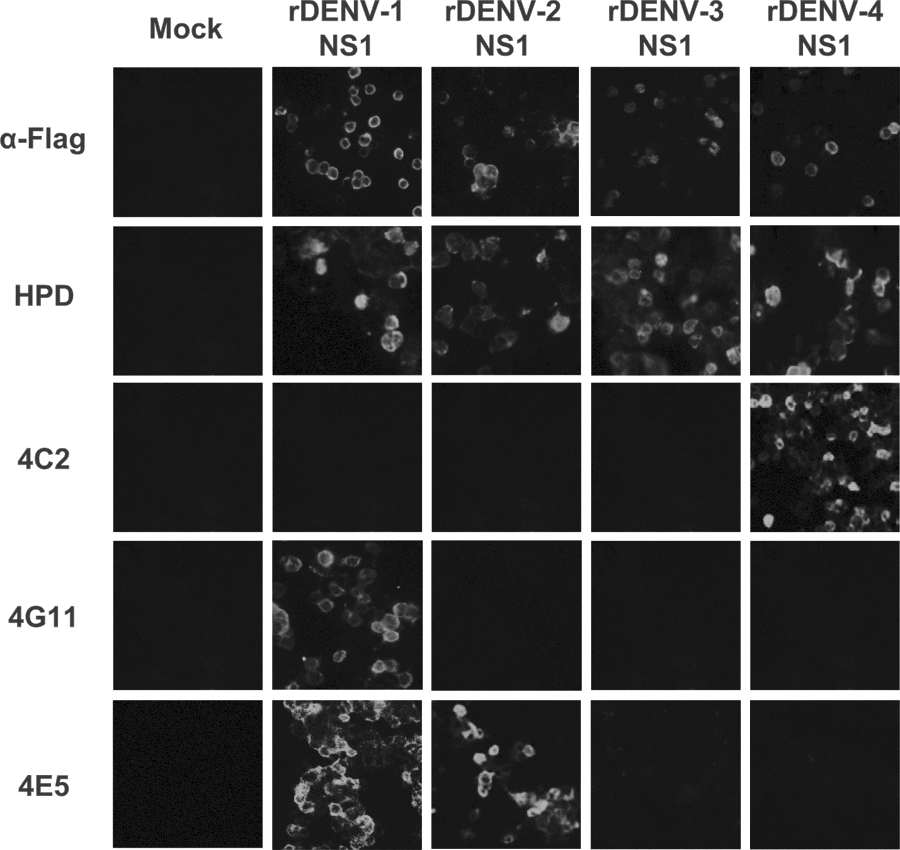
Reactivity of MAbs 4C2 and 4H11 with recombinant NS1 proteins. Recombinant NS1 proteins corresponding to individual DENV serotypes were transfected into 293T cells. The reactivity of each MAb with cells expressing each of the recombinant NS1 proteins was then examined. An α-FLAG antibody and HPD were used as positive controls.
MAbs 4C2 and 4G11 cross-react with the prM protein
To examine whether MAbs 4C2 and 4G11 interacted with other DENV-proteins, we performed immunoprecipitation assays using DENV-2-infected cells. HPD reacted with the E, NS1 and prM proteins (Fig. 3; Lane 1). MAbs 4C2 and 4G11 both precipitated the prM protein (Fig. 3; lanes 4, 5 and 6 [compare with lanes 5 and 8]). As expected, the E, NS1, and prM proteins of DENV-2 were precipitated by anti-E, -NS1 and -prM MAbs (Fig. 3; lanes 11, 13, and 15). We also confirmed that DENV-2 NS1 was not precipitated by MAbs 4C2 or 4G11 (Fig. 3; lanes 3, 4, 6 and 7). MAb 4E5 did not precipitated DENV-2 prM, but NS1 precipitated DENV-2 NS1 (Supplementary Data 2A). Presumably, 4C2 and 4G11 neutralized DENV-2 and protected mice from lethal infection with DENV-2 by interaction with prM (some DENV progeny viruses express prM) (20, 21, 25).
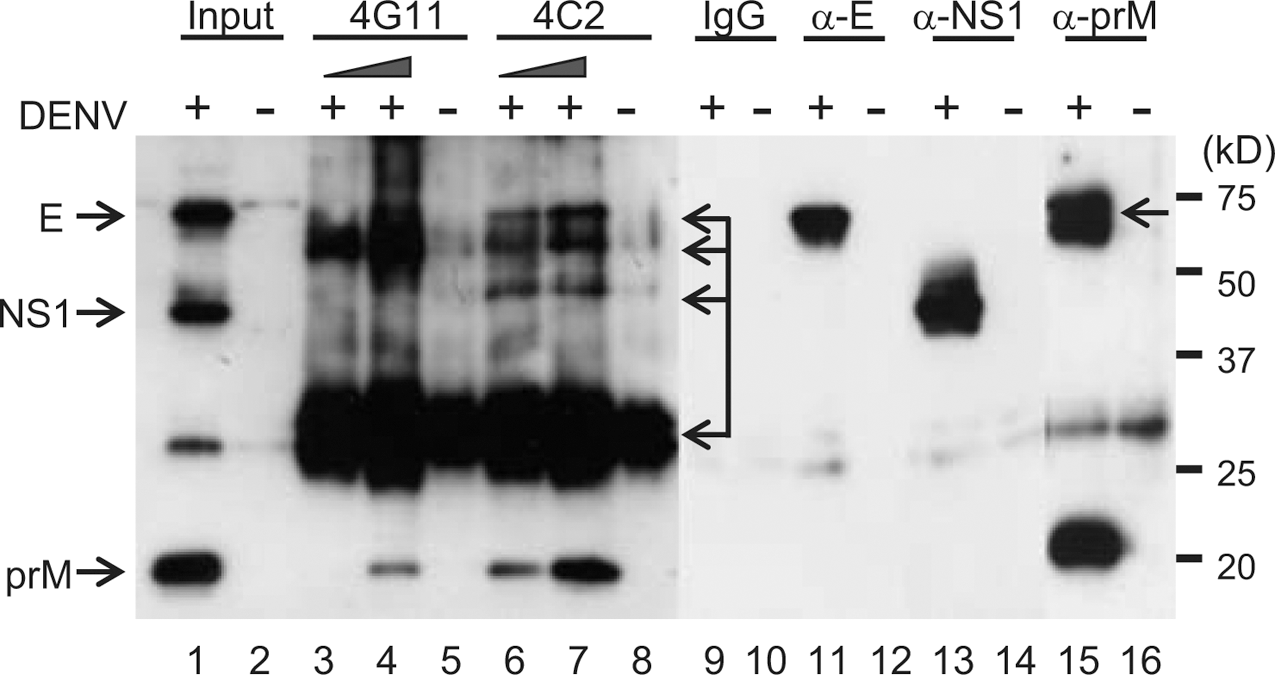
Cross-reactivity of MAbs 4G11 and 4C2 with the DENV-2 prM protein. Cell lysates infected with or without DENV-2 were stained with HPD (lanes 1 and 2). Lysates from DENV-2-infected BHK cells were co-precipitated with the following MAbs: 4G11 (1 μg, lane 3; 5 μg, lane 4; mock, lane 5); 4C2 (1 μg, lane 6; 5 μg, lane 7; mock, lane 8); mouse IgG (lanes 9 and 10); α-E 4G2 (lanes 11 and 12), α-NS1 1A5 (lanes 13 and 14), and α-prM 6D6 (lanes 15 and 16). The arrows indicate nonspecific bands.
MAbs 4C2 and 4G11 react with the N-terminus of DENV-4 NS1 and the C-terminus of DENV-1 NS1, respectively
Although MAbs 4C2 and 4G11 reacted with DENV-4 NS1 and DENV-1 NS1, respectively, but not with DENV-2 (Fig. 2), they both reacted with DENV-2 prM in the immunoprecipitation assays (Fig. 3). These results suggest that DENV-1 NS1 or DENV-4 NS1 share cross-reactive regions with DENV-2 prM. Therefore, we used recombinant mutant NS1 expressed in 293T cells to map the antibody target epitopes. MAb 4C2 and 4G11 recognized different regions of NS1. MAb 4C2 reacted with the N-terminal region of DENV-4 NS1 (aa 1–168), whereas 4G11 reacted with the C-terminal region of DENV-1 NS1 (aa 169–312) (Fig. 4).
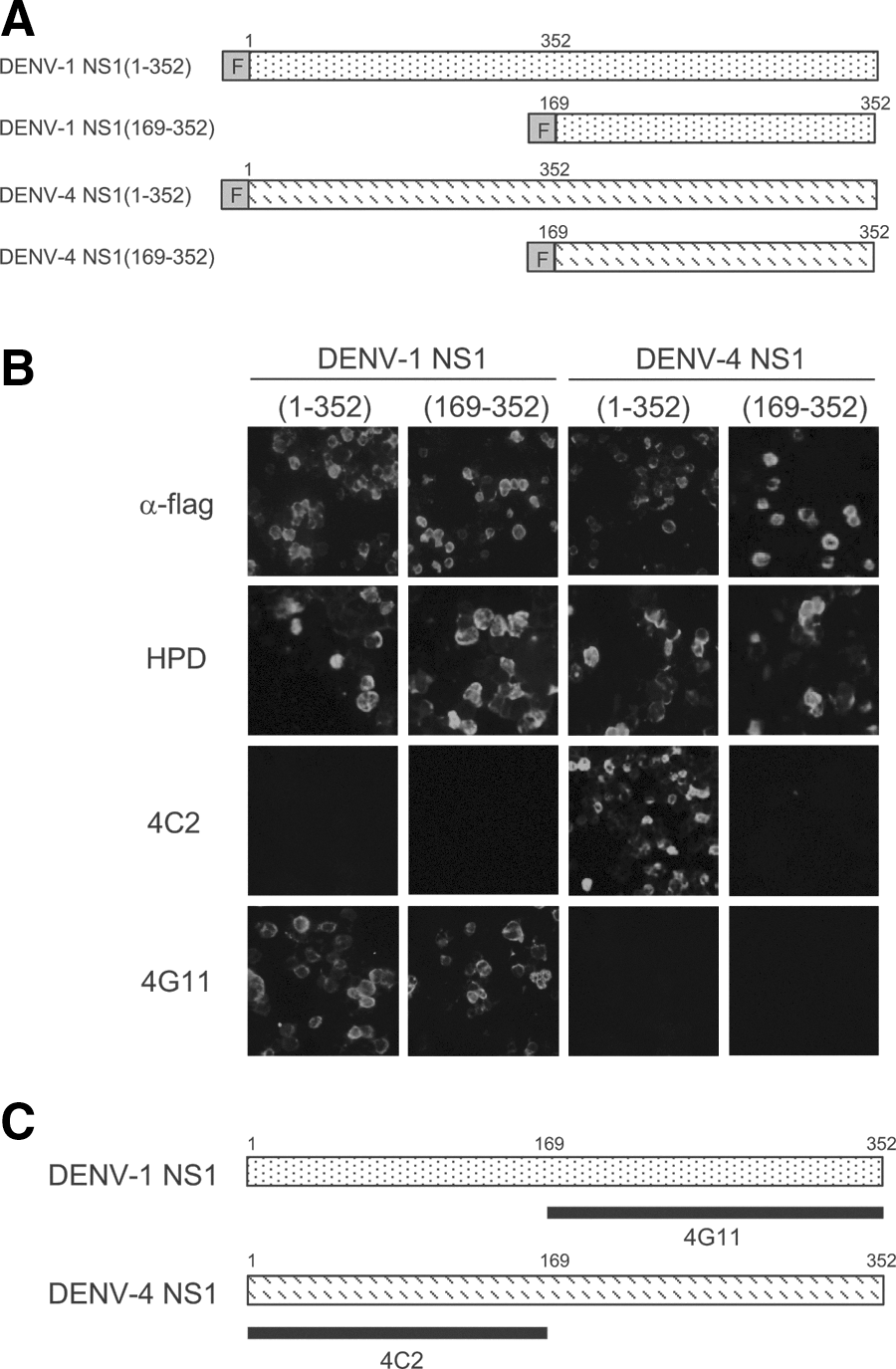
Epitopes recognized by MAbs 4C2 and 4G11.
MAb 4G11 facilitates DENV-2 infection of THP-1 cells
MAbs against prM increase viral infectivity because they capture the prM protein expressed by virus particles, thereby facilitating the infection of immune cells bearing Fc receptors (8). Because MAbs 4C2 and 4G11 cross-reacted with the DENV-2 prM protein (Fig. 3), we next examined whether they also increased DENV-2 infectivity via ADE. THP-1 cells were infected with mixtures of DENV-2 and serially-diluted MAbs. The anti-E MAb (4G2) increased infectivity by approximately 5-fold. Noteworthy, MAb 4G11 increased infection by approximately 4-fold compared with a control mouse IgG, but MAb 4C2 did not (Fig. 5). MAb 4G11 showed ADE activity at high concentrations (0.5–5 mg/mL), whereas anti-E 4G2 showed ADE activity at lower concentrations (0.5–5 μg/mL). Concentration of MAbs is thought to be a key for ADE activity.
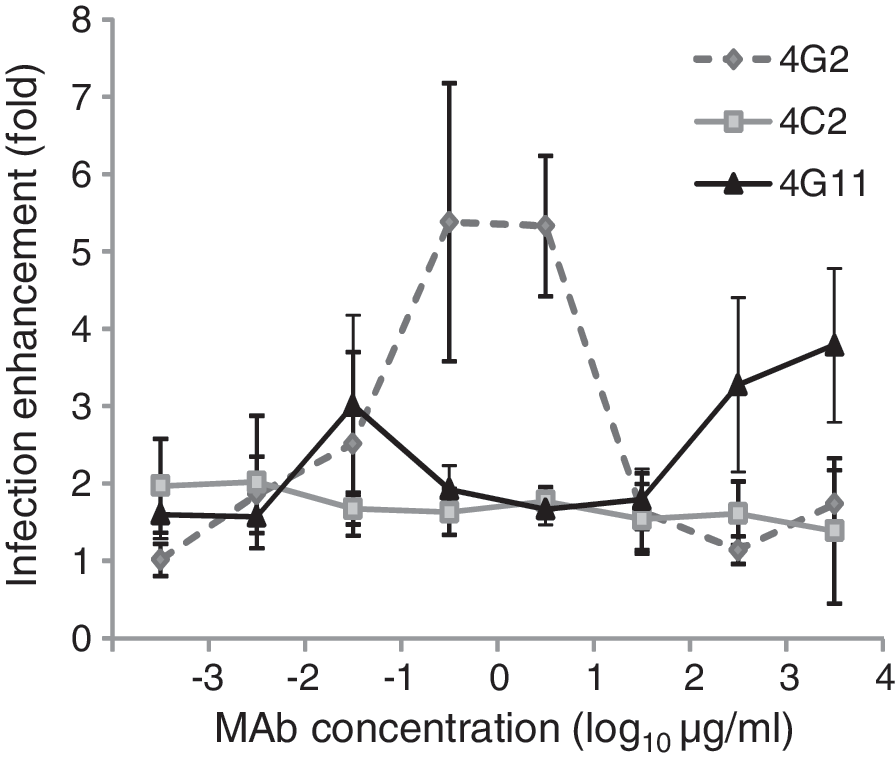
MAbs enhance DENV infection. THP-1 cells were infected with DENV-2 at a MOI of 0.5 in the presence of a set of serially-diluted MAbs (4C2, 4G11, 4G2) or mouse normal IgG. Viral RNA was examined by quantitative RT-PCR. Enhancement of infection is expressed in terms of “fold activation” with reference to the IgG control.
Discussion
In the present study, we investigate the new role of anti-DENV NS1 in the control of virus production. Two anti-NS1 specific MAbs, namely 4C2 and 4G11, showed moderate protection against DENV2 infection both in vitro and in vivo studies. However, their neutralizing activity does not seem to be caused by the direct interaction with NS1, but rather by cross-reactivity to DENV-prM (Fig. 3). Although anti-NS1 Abs repress virus replication by directly interacting with NS1 protein (12,17,35), the neutralizing activity of MAbs 4C2 and 4G11 against DENV-2 observed in this study can be attributed to cross-reactivity with prM (Fig. 3). The prM protein is expressed only on immature virions; however, recent studies show that DENV is released as a mixture of mature, partially mature, and immature particles (6,8). Immature virions are noninfectious; however, they can become infectious once they are introduced into cells via anti-prM Abs (the endosomal protease furin processes the prM protein) (25,31). Abs against prM do not have strong neutralizing activity; rather, they facilitate infection via ADE (8). In the present study, MAb 4G11 facilitated the infection of THP-1 cells by DENV-2 (Fig. 3). This is the first report showing that an anti-NS1 MAb cross-reacts with prM protein and facilitates infection by DENV. Besides their direct binding to NS1, Abs against NS1 may have additional effects during infection. Although it is unclear how anti-NS1 Abs in DENV-infected patients are involved in infection, we suggest that anti-NS1 Abs positively or negatively regulate virus production.
DENV infection of humans induces a strong B-cell response against the E, prM, and NS1 (32,39). Although Ab responses to other nonstructural proteins, such as NS3 and NS5, have been detected, the production of specific Abs is low and appears not to contribute to disease progression (32). However, the production of anti-NS1 Abs is high, particularly in the case of secondary infections (24,39), and anti-NS1 Abs are detected during the acute phase of secondary infections and during the convalescent phase (23,24). This suggests a potential interaction between the virus particles and the anti-NS1 Abs in the patients' blood at the early stage of disease progression. Even though anti-NS1 Abs cross-react with virus particles, this interaction is expected to be relatively weak. Indeed, Falconar reported that monoclonal/polyclonal Abs against NS1 cross-react with the E protein and show only weak neutralizing activity (10,11). The weak neutralizing activity of cross-reactive Abs is generally regarded as immaterial because it does not contribute to the clearance of pathogens; in other words, these cross-reactive Abs are not involved in progression of disease. However, the situation of DENV infection is different from other infectious diseases because the Abs often facilitate infection (16). Non-neutralizing or weakly neutralizing Abs are thought to contribute to ADE (38). This also occurs with other viruses that target cells bearing Fc receptors (30). Recently, Meyer et al. reported that a weak neutralizing Ab response to the hepatitis C virus envelope glycoprotein facilitates virus infection (29). This is a major concern for vaccine development and for the use of MAbs as therapeutic agents. This study points to a potential interaction between anti-NS1 Abs and partially mature virions at early stage in disease progression, resulting in neutralization or ADE. The concentration of Abs also determines the balance between neutralization and ADE. MAb 4G11 showed ADE activity when present at high concentrations (Fig. 5), suggesting that anti-NS1 Abs potentially enhance infection when anti-NS1 Abs are present at particularly high concentrations during secondary infections.
Flavivirus NS1 is thought to be involved in pathogenesis of human disease and mouse model (2,13,26), although the detailed mechanism remains unknown. Immunization with naïve or recombinant NS1 protects mice from lethal dengue encephalitis (12,35,44) and non-neutralizing, serotype-specific anti-NS1 MAbs partially protect passively immunized mice from lethal infection (17). DENV NS1 also activates complement in patients infected with DENV, which leads to the production of anaphylatoxin (2). By contrast, Chung et al., reported that extracellular NS1 produced by cells infected with West Nile Virus inhibits complement activation, which allows viruses to evade the immune system (5). These reports suggest that NS1 and anti-NS1 Abs are involved in the pathogenesis of these diseases. In those studies, the protective ability of anti-NS1 Abs was likely due to their reactivity with NS1, because non-neutralizing Abs specific for NS1 protect mice from lethal yellow fever virus encephalitis (33). In this study, MAbs 4C2 and 4G11 did not react with DENV-2 NS1 (Fig. 3), however, neutralize DENV-2 (Table 3). Contrary, MAb 4E5 showed lower neutralizing activity (Table 3) and did not protect mice from infection (Fig. 1). This is probably because it did not react with DENV-2 prM (Supplementary Data 2A). Furthermore, even though MAbs 4G11 and 4C2 react with NS1 derived from DENV-1 or DENV-4, respectively, they did not protect mice from the lethal infection with DENV-1 or DENV-4 (Supplementary Data 1). We also examined whether other MAbs to NS1 (MAbs 1A5, 2H11, 3C4, 3F10, 4E5, 5E2, and 5G12), which react with DENV-2 NS1 (28), protected suckling mice from lethal infection with DENV-2; however, none did (Supplementary Data 3). Virus neutralization, protection of suckling mice, and the ADE mediated by the anti-NS1 MAbs in the present study are all likely attributable to their ability to interact with the DENV-2 prM protein (Table 3 and Figs. 1 and 5). In another aspect, MAb 4G11 enhanced infection with DENV-2 in vitro (Fig. 5) but, nevertheless, it did not enhance mortality of suckling mice (Fig. 1 and Supplementary Data 4). This is probably because major target cells for DENV replication in the mouse brain are unlikely to be monocyte/macrophage bearing the Fc receptor.
Collectively, our data suggest that DENV-2 prM contains an epitope similar to that recognized by NS1 because MAbs to NS1 cross-react with prM (Fig. 3). MAbs 4G11 and 4C2 were mapped to different regions of NS1 (the C-terminal and N-terminal NS1 regions, respectively; Fig. 4). The epitopes recognized by MAbs 4C2 and 4G11 were analyzed using BLAST to identify similarities with DENV-2 prM; however, no candidate region was found (data not shown) probably because these MAbs presumably recognize conformational epitopes of prM and/or NS1. Although MAbs 4C2 and 4G11 precipitated prM under native condition (Fig. 3), these MAbs did not stain prM on Western-blots when prM was transferred from a denaturing SDS-polyacrylamide gel (data not shown). It has previously been shown that MAbs 4C2 and 4G11 have decreased reactivity to NS1 on Western blot when a reducing agent is employed (28). These observations suggest that MAbs 4C2 and 4G11 recognize conformational epitopes in both prM and NS1. Even though both MAbs interacted with the prM, showed similar FRNT50 values against DENV-2 (Table 3), and protected mice from lethal infection (Fig. 1), only 4G11 facilitated infection by DENV-2 in the ADE assay (Fig. 5), suggesting that these two MAbs recognize different regions of the DENV-2 prM. Further detailed study is needed.
Taken together, the findings of the present study lead us to the conclusion that anti-NS1 Abs should be analyzed for their neutralizing/enhancing activity and cross-reactivity to prM proteins. We also recently examined anti-NS1 Abs derived from patients to examine whether they are similarly cross-reactive (36).
Footnotes
Acknowledgments
This work was supported in part by the SYSMEX Corporation (Osaka, Japan). The manuscript was proofread by Bioedit (Manchester, UK). We thank Dr. Akifumi Yamashita for help with computational analysis. P.M was supported by the Ministry of Education, Culture, Sports, Science and Technology of Japan
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.