
Abstract
The implications of sequential prime and challenge with mismatched influenza A viruses is a concern in mammals, including humans. We evaluated the ability of pigs affected with vaccine-associated enhanced respiratory disease (VAERD) to generate a humoral immune response against the heterologous challenge virus inciting the VAERD. Vaccinated and challenged (V/C) pigs were administered an inactivated swine δ-cluster H1N2 (MN08) vaccine with an HA similar to pre-2009 seasonal human viruses and challenged with heterologous A(H1N1) pandemic 2009 (H1N1pdm09). Vaccination induced MN08-specific hemagglutination inhibition (HI) antibody but not cross-reacting H1N1pdm09 HI antibody. However, vaccinated pigs demonstrated significantly higher post-challenge anti-H1N1pdm09 serum neutralizing (SN) antibodies at 14 and 21 days post inoculation (dpi) compared to nonvaccinated, challenged pigs (NV/C), indicating a priming effect of the vaccine. Serum and lung whole virus anti-H1N1pdm09 IgG ELISA antibodies in the vaccinated group were significantly higher than the challenge only pigs at all-time points evaluated. Lung IgA ELISA antibodies to both antigens were detected at 2, 5, and 21 dpi in vaccine-primed pigs, contrasted against mucosal ELISA antibody responses detected only at 21 dpi in the naïve-challenged group. Collectively, vaccine-primed pigs demonstrated a robust humoral immune response and elevated local adaptive cytokine levels, indicating VAERD does not adversely affect the induction of an immune response to challenge with heterologous virus despite the severe clinical disease and underlying lung pathology. Thus, original antigenic sin does not appear to be a component of VAERD.
Introduction
As IAVs have become more genetically diverse in the swine population, serological cross-reactivity in the hemagglutination inhibition (HI) assay between H1 phylogenetic clusters have become more variable (20,41). Inactivated vaccines have demonstrated limited cross-protection against heterologous homosubtypic, or heterosubtypic influenza viruses (18,38 –40,42,43). In addition, inactivated vaccines have been associated with enhanced respiratory disease in swine (7) or potentially increasing the risk of illness in H1N1pdm09-infected humans who previously received the 2008–09 trivalent inactivated influenza vaccine (12,30). We recently reported a disease phenomenon in swine vaccinated with monovalent inactivated δ-cluster H1N2 vaccine and exposed to H1N1pdm09, termed vaccine-associated enhanced respiratory disease (VAERD) (7). The mechanism responsible for VAERD is unknown at this time, although the antibody response elicited by inactivated vaccines has been implicated (7,10,42). A previous report also suggested severe respiratory disease may have been associated with the immune response to prior seasonal influenza virus infection or vaccination in humans infected with H1N1pdm09 (32).
Pigs with VAERD demonstrate severe, prolonged respiratory disease and increased macroscopic and microscopic lung lesions; however, VAERD-affected pigs in experimental settings recover from disease in spite of the negative health impact (7). Due to the widespread use of inactivated vaccines and the level of reassortment and antigenic drift among IAV's in swine, it is increasingly important to understand how prior vaccine immunity may affect the immune response to the heterologous challenge virus that incited VAERD and if original antigenic sin is associated with VAERD.
Materials and Methods
Experimental design, vaccine and virus inoculum
Ninety-one 3-week-old cross-bred pigs of mixed sex were obtained from a herd free of IAV and porcine reproductive and respiratory syndrome virus (PRRSV). Upon arrival, pigs were treated prophylactically with ceftiofur (Pfizer Animal Health, New York, NY) according to label directions to reduce potential respiratory bacterial pathogens and were housed in biosafety level 2 (BSL2) containment during the vaccine phase of the study, and at ABSL3 containment for the challenge phase as previously described (6). Pigs were cared for in compliance with the Institutional Animal Care and Use Committee of the National Animal Disease Center. The experimental design is described in Table 1.
BALF, bronchoalveolar lavage fluid; dpi, days post inoculation; H1N1pdm09, pandemic A/CA/04/2009 H1N1; MN08, swine δ-cluster A/Sw/MN/02011/2008 inactivated vaccine; NV/C, nonvaccinated/challenged; NV/NC, nonvaccinated/nonchallenged; V/C, vaccinated/challenged.
Whole influenza A virus vaccine was prepared with A/Sw/MN/02011/2008 (MN08) δ1 cluster H1N2 at approximately 32 HA units or 105.7 50% tissue culture infectious dose (TCID50) per mL and mixed with adjuvant as previously described (7). Pigs were vaccinated with 2 mL by the intramuscular route at approximately 6 and 9 weeks of age, and challenged by the intratracheal route at 13 weeks of age with 2 mL of 1×105 TCID50 of pandemic A/CA/04/2009 H1N1 (H1N1pdm09) as described previously (6). Pigs were humanely euthanized with a lethal dose of pentobarbital (Sleepaway, Fort Dodge Animal Health, Fort Dodge, IA) on 0, 1, 2, 5, or 21 days post infection (dpi) to evaluate lung lesions, viral replication. and innate cytokine concentrations as previously described (6), and the post-challenge humoral immune and adaptive cytokine responses reported here. Serum was collected from four pigs in each group at 0, 7, 14, and 21 dpi. Postmortem samples included serum (12 pigs) and bronchoalveolar lavage fluid (BALF) (91 pigs) for this study.
Diagnostic microbiology
All pigs were screened for antibody against influenza A NP by ELISA (MultiS ELISA, IDEXX, Westbrook, Maine) upon arrival to ensure absence of preexisting immunity from prior exposure or maternally derived antibody. BALF samples from 1, 2, 5, and 21 dpi were cultured for aerobic bacteria on blood agar and Casmin (NAD enriched) plates. Polymerase chain reaction (PCR) assays for PCV2 (26), Mycoplasma hyopneumoniae (31) and RT-PCR for PRRSV were conducted on nucleic acid extracts from BALF collected at each necropsy.
Antibody detection assays
Hemagglutination inhibition assays were conducted on serum from twelve pigs (4 per group) at 0, 7, 14, and 21 dpi with MN08 or H1N1pdm09 virus as antigen and turkey RBCs as indicators using standard techniques as previously described (42). Reciprocal titers were divided by 10 and log2 transformed, analyzed, and reported as the geometric mean.
Enzyme-linked immunosorbent assays (ELISA) to detect total IgG and IgA antibodies against whole virus preparations of MN08 and H1N1pdm09 present in serum from 12 pigs (4 per group) at 0, 7, 14, and 21 dpi, and BALF from 91 pigs (Table 1) at 0, 1, 2, 5, and 21 dpi were performed as previously described with modifications (7). Sera were heat inactivated at 56°C for 30 min and diluted in 5% Fraction V bovine serum albumin (BSA) (Life Technologies/Gibco, Grand Island, NY) in phosphate-buffered saline (PBS) at 1:2000 and 1:4 for IgG and IgA assays, respectively, to adsorb nonspecific binding. The BALF samples were incubated at 37°C for 1 h with an equal volume of 10 mM dithiothreitol (DTT) to disrupt mucus prior to dilution with an equal volume of 10% BSA-PBS resulting in a final BALF dilution of 1:4. The BSA-treated sera and BALF were incubated at 37°C for 1 h immediately prior to testing.
Concentrated MN08 and H1N1pdm09 were resuspended in Tris-EDTA basic buffer, pH 7.8, and diluted to an HA concentration of 100 HA units/50 μL. Immulon-2HB 96-well plates (Dynex, Chantilly, VA) were coated with 50 μL of either antigen and incubated at room temperature overnight. Plates were blocked for 1 h at room temperature with 150 μL of Starting Block (Thermo Fisher Scientific, Pittsburgh, PA) and washed 3 times with 0.05% Tween 20 in PBS (PBS-T). The assays were performed with 50 μL of diluted sera or BALF in duplicate. Plates were incubated at room temperature for 1 h, washed 3 times with PBS-T, then incubated with peroxidase-labeled goat anti-swine IgG (Kirkegaard & Perry Laboratories, Gaithersburg, MD) or IgA (Bethyl, Montgomery, TX) diluted 1:1,500 in Starting Block for 1 h at room temperature. Plates were washed 3 times with PBS-T; then 50 μL of 2, 2′-azino-di(3-ethylbenzthiazoline-6-sulfonate) (ABTS)-peroxide was added as the substrate (Kirkegaard & Perry Laboratories) for 8–10 min prior to adding 50 μL KPL Stop Solution (Kirkegaard & Perry Laboratories). The optical density (OD) was measured at 405 nm wavelength with an automated ELISA reader. Antibody levels were reported as the mean OD for each duplicate sample, and the mean OD of each treatment group was compared.
Serum neutralization (SN) assays were conducted on serum from 12 pigs (4 per group) at 0, 7, 14, and 21 dpi as previously described (7,43). Titers were recorded as the highest dilution negative for virus staining or CPE after the presence of virus was verified by immunocytochemistry for all plates. Reciprocal titers were divided by 10, log2 transformed, analyzed, and reported as the geometric mean.
Cytokine assays
A 5 mL aliquot of BALF collected from pigs at 0, 1, 2, 5, and 21 dpi was centrifuged at 400× g for 15 min at 4°C to pellet cellular debris. Cytokine concentrations in BALF, which included IL-2, IL-4, IL-12p70, IL-10, and IFN-γ, were determined by multiplex ELISA performed according to the manufacturer's recommendations (SearchLight, Aushon Biosystems, Billerica, MA). Cytokine concentrations were reported as the mean of duplicate samples for statistical analysis.
Statistical analysis
Log2 transformed HI and SN reciprocal titers, mean OD ELISA IgG and IgA antibody levels and cytokine concentrations were analyzed using analysis of variance (ANOVA) with a p value ≤0.05 considered significant (JMP, SAS Institute, Cary, NC; GraphPad Prism Version 5.00, San Diego, CA). Response variables shown to have a significant effect by treatment group were subjected to pair-wise comparisons using the Tukey–Kramer test.
Results
Viral and microbiological assays
Prior to vaccination, all pigs were free of influenza A NP specific antibodies and at 5 dpi, all pigs were negative for PRRSV, PCV2, or M. hyopneumoniae nucleic acids in BALF. Bacterial pathogens were not detected in the lung with the exception of Bordetella bronchiseptica in the BALF of one NV/C pig. Macroscopic and microscopic lung lesions indicative of VAERD were present in all V/C pigs (VAERD group) reported previously with nasal shedding and lung virus titers (6).
VAERD did not prevent induction of HI or SN antibody responses to heterologous challenge
To evaluate the effect of VAERD on the induction of H1N1pdm09 specific HI or SN antibody responses, V/C pre- and post-challenge titers were compared to the naïve-challenged group (NV/C) (Table 2). Anti-MN08 or -H1N1pdm09 HI antibodies were not detected in pigs that did not receive vaccine or challenge virus, respectively, during the pre- and/or post-challenge phase of the study. The geometric mean anti-MN08 HI antibody titer in the VAERD group was significantly higher (p≤0.05) than the NV/C and nonvaccinated/nonchallenged (NV/NC) group titers at 7, 14, and 21 dpi. Challenge with H1N1pdm09 boosted the mean anti-MN08 HI antibody titer in the V/C pigs, peaking at 14 dpi. However, post-challenge anti-H1N1pdm09 HI antibodies did not cross-react with MN08 antigen in nonvaccinated pigs at any time point. This is consistent with the lack of HI cross-reactivity previously demonstrated between the vaccine and challenge viruses and consistent with the VAERD model (41).
Geometric mean reciprocal HI titers with different letters within a column are significantly different (p≤0.05).
dpi, days post inoculation; H1N1pdm09, pandemic A/CA/04/2009 H1N1; MN08, swine δ-cluster A/Sw/MN/02011/2008; NV/C, nonvaccinated/challenged; NV/NC, nonvaccinated/nonchallenged; V/C, vaccinated/challenged.
Importantly, the vaccine-primed, VAERD-affected group demonstrated a humoral immune response to the H1N1pdm09 challenge virus in spite of the severe underlying pathology described previously (6). Moreover, the geometric mean V/C anti-H1N1pdm09 HI antibody titers were significantly higher (p≤0.05) than the NV/C and NV/NC group at 7 dpi and demonstrated elevated anti-H1N1pdm09 HI titers at 14 and 21 dpi compared to the naïve challenged pigs.
Anti-MN08 or -H1N1pdm09 SN antibodies were not detected in pigs that did not receive vaccine or challenge virus, respectively, during the pre- and/or post-challenge phase of the study (Table 3). The VAERD-affected group geometric mean anti-MN08 SN antibody titers were boosted post-challenge with H1N1pdm09, peaked earlier at 7 dpi compared to the anti-MN08 HI titer and declined thereafter. In addition, naïve-challenged pigs did not demonstrate cross-reactive neutralizing antibodies to MN08 in the absence of vaccination.
Geometric mean reciprocal SN titers with different letters within a column are significantly different (p≤0.05).
dpi, days post inoculation; H1N1pdm09, pandemic A/CA/04/2009 H1N1; MN08, swine δ-cluster A/Sw/MN/02011/2008; NV/C, nonvaccinated/challenged; NV/NC, nonvaccinated/nonchallenged; V/C, vaccinated/challenged.
The geometric mean anti-H1N1pdm09 SN antibody titers in the VAERD-affected group was similar to the primary immune response demonstrated in the NV/C group at 7 dpi but were significantly higher (p≤0.001) than the NV/C and NV/NC groups at 14 and 21 dpi. These data suggest VAERD does not preclude the HI or SN immune response to the heterologous virus inciting the enhanced disease.
Serum and lung IgG whole virus anti-MN08 and cross-reactive anti-H1N1pdm09 antibody responses in VAERD-affected pigs
Pre- and post-challenge serum was evaluated for the presence of cross-reactive, whole virus IgG antibodies induced by the MN08 inactivated vaccine (Table 4). Anti-MN08 and cross-reactive anti-H1N1pdm09 systemic IgG antibodies were detected in the V/C group prior to challenge at 0 dpi, suggesting the presence of conserved, although nonprotective, epitopes between vaccine and challenge viruses. Group mean pre- and post-challenge anti-MN08 and -H1N1pdm09 OD levels were significantly higher (p≤0.001) in vaccine-primed pigs compared to NV/C and NV/NC pigs at all dpi measured and demonstrated an increasing trend in post-challenge OD levels. The NV/C group demonstrated anti-H1N1pdm09 and cross-reactive anti-MN08 post-challenge IgG OD levels that were significantly higher (p≤0.05) than the NV/NC pigs at 14 and 21 dpi, indicating a primary immune response to the challenge virus that was also cross-reactive against the vaccine antigen.
Mean OD levels for each antigen with different letters within a column are significantly different (p≤0.05).
BALF, bronchoalveolar lavage fluid; dpi, days post inoculation; H1N1pdm09, pandemic A/CA/04/2009 H1N1; MN08, swine δ-cluster A/Sw/MN/02011/2008; ND, not done; NV/C, nonvaccinated/challenged; NV/NC, nonvaccinated/nonchallenged; V/C, vaccinated/challenged.
Lung lavage fluid was evaluated to assess the level of vaccine-induced, pre- and post-challenge whole virus mucosal IgG antibodies in VAERD-affected pigs (Table 4). IgG antibodies against MN08 and H1N1pdm09 were detected at significantly higher (p≤0.05) levels in the lungs from vaccine-primed pigs prior to challenge and at 1, 2, 5, and 21 dpi compared to NV/C and NV/NC groups. Nonprimed, challenged pigs demonstrated anti-H1N1pdm09 and cross-reactive anti-MN08 BALF IgG antibody levels that were significantly higher (p≤0.001) than the NV/NC group only at 21 dpi, a delay compared to serum IgG responses. These data suggest vaccine-induced, cross-reactive whole virus antibodies against H1N1pdm09 were present at the time of challenge in VAERD-affected pigs. Challenge with H1N1pdm09 boosted IgG levels immediately post-challenge (1 dpi), suggesting transudation of antibodies into the lung at the site of infection.
Serum and lung IgA whole virus anti-MN08 and cross-reactive anti-H1N1pdm09 antibody responses in VAERD-affected pigs
Serum IgA levels specific for MN08 vaccine antigen and challenge H1N1pdm09 virus were evaluated pre- and post-inoculation in VAERD-affected pigs (Table 5). Challenge with H1N1pdm09 did not induce a detectable serum IgA antibody response to either antigen regardless of prior vaccine priming. In addition, the MN08 inactivated vaccine did not prime for detectable levels of local IgA in the lower respiratory tract, as anti-MN08 IgA mucosal antibodies were not significantly different in the V/C group at 0 or 1 dpi compared to the NV/NC pigs. However, at 2 and 5 dpi, the anti-MN08 and -H1N1pdm09 group mean IgA antibody levels in the lungs of vaccine-primed pigs were significantly higher (p≤0.05) than the NV/C and NV/NC groups, suggesting the inactivated vaccine administered intramuscularly primed for a more rapid local immune response to challenge. In contrast, 21 dpi anti-MN08 and -H1N1pdm09 group mean V/C and NV/C OD levels were similar and significantly higher (p≤0.0001) than the NV/NC group, suggesting a primary IgA response to the H1N1pdm09 challenge virus in NV/C pigs. This indicates mucosal challenge with live virus stimulated a cross-reactive IgA immune response, in contrast to the undetectable IgA antibodies induced by the inactivated vaccine.
Mean OD levels for each antigen with different letters within a column are significantly different (p≤0.05).
BALF, bronchoalveolar lavage fluid; dpi, days post inoculation; H1N1pdm09, pandemic A/CA/04/2009 H1N1; MN08, swine δ-cluster A/Sw/MN/02011/2008; ND, not done; NV/C, nonvaccinated/challenged; NV/NC, nonvaccinated/nonchallenged; V/C, vaccinated/challenged.
Post-challenge local adaptive cytokine responses are increased in VAERD affected pigs
Cytokine protein concentrations in BALF for IL-2, IL-4, IL-12p70, IFN-γ, and IL-10 were measured to evaluate T cell responses in the lungs of VAERD-affected pigs (Fig. 1). All cytokines were significantly higher (p<0.05) at 5 dpi (necropsy) in the vaccine-primed pigs who demonstrated VAERD. Additionally, V/C pigs demonstrated significantly higher (p<0.05) IFN-γ levels compared to the other two groups at 1 and 2 dpi, suggesting prior vaccination may have resulted in the induction of memory T cells as previously described in MN08-vaccinated pigs and T cell activation upon challenge (28). However, elevated IL-10 may be a response by the host to inhibit pro-inflammatory cytokine production, IFNγ, or natural killer cells that may be contributing to the immunopathology described in VAERD-affected pigs (6,24). Concentrations of adaptive cytokines in the V/C pigs returned to baseline levels by 21 dpi and no significant differences were observed between the groups.
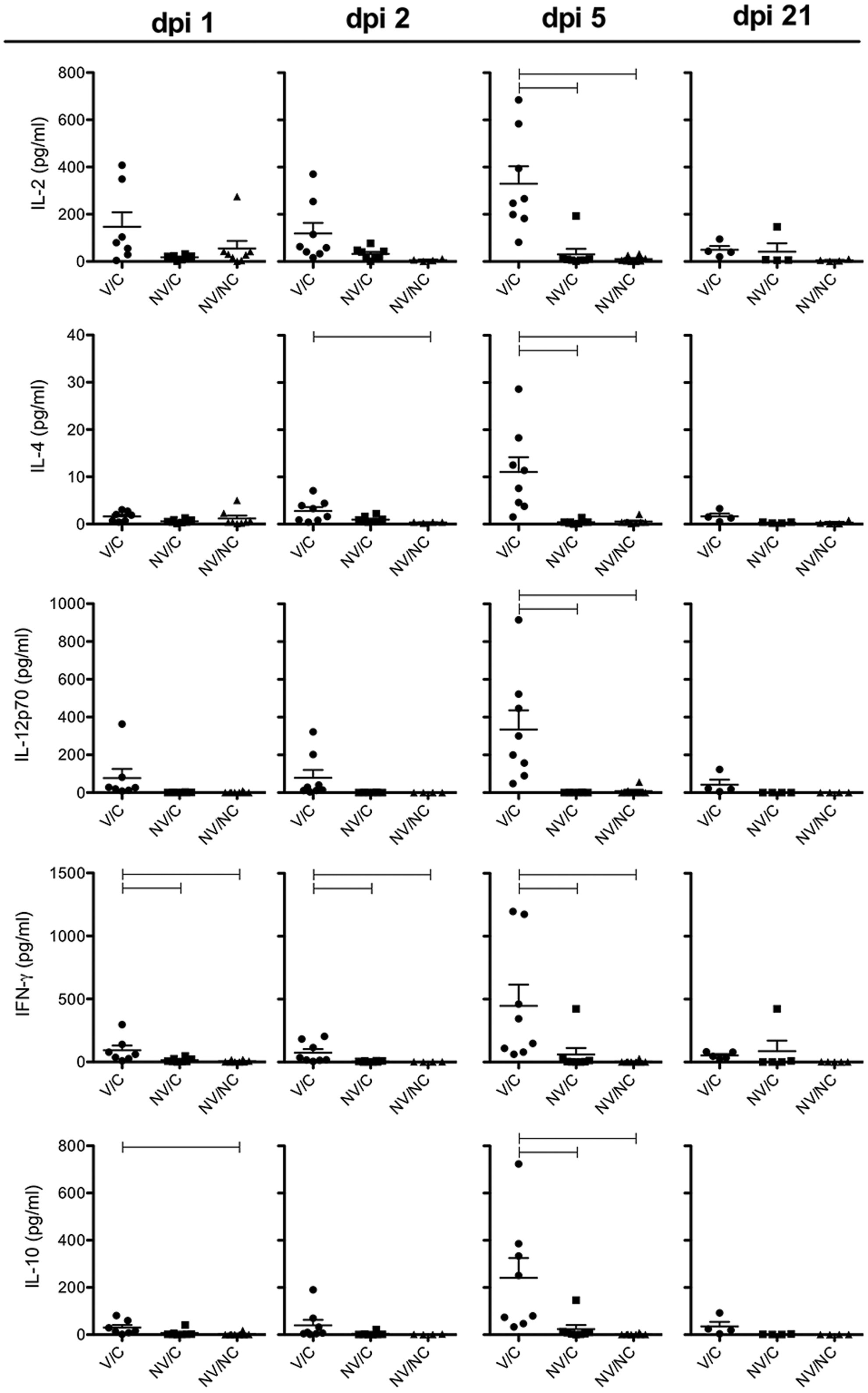
Cytokine protein levels (pg/mL) in bronchoalveolar lavage fluid of vaccinated and H1N1pdm09-challenged (V/C), nonvaccinated and H1N1pdm09-challenged (NV/C), and nonvaccinated and nonchallenged (NV/NC) pigs measured with multiplex ELISA at 1, 2, 5, and 21 days post inoculation (dpi). Each symbol represents the cytokine-specific level of individual pigs. Bars represent the mean±standard error of the mean. Connecting horizontal lines are significantly different (p<0.05).
Discussion
Genetic mutation and reassortment drives the evolution of antigenically diverse IAV's in swine (20 –22,44). Genetic diversity has been demonstrated in all currently identified H1 phylogenetic clusters (α-, β-, γ-, δ1, and δ2); additionally, the δ-cluster viruses exhibit greater genetic divergence compared to the other three clusters (20,21,44). Therefore, vaccine-challenge mismatch is more likely between influenza viruses derived from the cH1N1 HA lineage and δ-cluster viruses. The objective of the current study was to evaluate if VAERD-affected pigs establish an adaptive immune response specific to the heterologous challenge virus, H1N1pdm09, or if original antigenic sin would predominate. We have previously demonstrated VAERD using a model where pigs were vaccinated with an inactivated δ1-cluster (human-like) IAV and challenged with H1N1pdm09 (6,7).
Surviving cohort pigs in the VAERD group developed anti-H1N1pdm09 HI and SN antibodies in a similar pattern as the naïve NV/C pigs, suggesting neither antecedent vaccine immunity nor VAERD affected the induction of a post-challenge immune response to heterologous H1N1pdm09. The VAERD-affected, vaccine-primed pigs challenged with H1N1pdm09 responded with mildly elevated HI or significantly elevated SN titers against H1N1pdm09, suggesting that priming with inactivated vaccines may result in a more robust SN antibody response compared to HI antibody levels, regardless of the challenge virus. Conversely, the SN assay may be more sensitive and able to demonstrate modest increases in antibody levels undetectable by the HI assay, implying the SN test may be more sensitive for assessing functional anti-influenza antibody responses in swine. Serum neutralization titers have reportedly been higher compared to HI titers in assays using homologous swine anti-sera and antigen (35). Previous reports have also demonstrated anti-influenza SN antibodies in humans that were seronegative by the HI test further indicating a heightened sensitivity of SN tests (8).
Anti-MN08 HI and SN antibodies in the V/C group were boosted post H1N1pdm09 challenge compared to pre-challenge levels demonstrated at 0 dpi. Priming naïve pigs with inactivated IAV resulted in a more robust post-challenge humoral immune response to both the priming antigen and heterologous challenge virus. This is in spite of the lack of HI/SN cross-reactivity between vaccine and inoculum strains prior to secondary exposure and in spite of the dramatic lung pathology associated with VAERD. Similar results were reported in pigs where preexisting HI and SN immune responses to primary inoculation increased following challenge with an antigenically different influenza virus of the same subtype (35). In that same study, administration of an inactivated influenza vaccine to pigs previously inoculated with heterologous virus also boosted preexisting antibody, suggesting this phenomenon may occur regardless of how the immune system is initially primed (35). In the current study, elevated anti-H1N1pdm09 HI and SN titers in the MN08 vaccine-primed pigs after challenge suggests pigs may be protected from a subsequent re-exposure to H1N1pdm09. Detection of post-challenge anti-H1N1pdm09 HI and SN antibodies in the MN08 vaccine-primed pigs in addition to the boosted antibody levels to the priming MN08 vaccine antigen does not fit the concept of original antigenic sin where antibodies exclusive to the priming antigen are augmented at the expense of a response to unique epitopes presented by the challenge virus (4,5,9). As the two viruses evaluated in this study are both of the H1 subtype and demonstrate non-neutralizing antibody cross-reactivity, they may be too similar to induce original antigenic sin or it is possible inactivated vaccines are less prone to induce the phenomenon (16).
In contrast to the absent cross-reactive anti-H1N1pdm09 HI or SN antibodies in the vaccine-primed pigs prior to challenge, whole virus cross-reacting anti-H1N1pdm09 IgG ELISA antibodies were detected in serum and BALF in pigs euthanized at 0 dpi in the V/C pigs. This suggests cross-reactive lung IgG antibodies were likely present at the time of challenge. However, the increase between 0 and 5 dpi also suggests a rapid post-challenge transudation across pulmonary capillaries from systemic IgG in the V/C pigs. The vaccine-induced anti-H1N1pdm09 systemic or mucosal IgG antibodies were not cross-protective as confirmed by the presence of infectious virus in the lungs and nasal swabs of V/C pigs from this study described previously (6) and in an additional study (7). Although plausible, it remains unknown if these cross-reacting IgG antibodies contributed to the enhanced pneumonia described in VAERD (7).
Unlike the anti-MN08 and -H1N1pdm09 IgG antibodies detected in the lungs of V/C pigs, the inactivated vaccine did not induce a measureable IgA antibody response to either antigen by our methods prior to challenge. Consistent with previous reports, inactivated influenza vaccines administered intramuscularly to swine induce lower levels of mucosal IgA compared to live inoculation or intranasal vaccination with live attenuated influenza vaccines (LAIV), suggesting mucosal immunization is required for adequate production of IgA antibody secreting cells to protect the upper respiratory tract (11,19,27,45). In contrast, live exposure or mucosal vaccination induced cross-reactive IgA, since challenge with H1N1pdm09 induced a cross-reactive anti-MN08 IgG and IgA response in the lungs of NV/C pigs at 21 dpi. Although likely, it is unknown if the anti-MN08 IgA response detected in the lungs of NV/C pigs in this study is predictive of cross-protection against subsequent infection with the same or other heterologous IAV without further study.
Influenza A virus-induced pro-inflammatory cytokine responses in swine have been extensively studied and shown to be correlated with viral replication, disease, and lung pathology (1,7,13,14,33,34,36,37). In addition, elevated pro-inflammatory cytokines associated with the innate immune system have been associated with the VAERD phenomenon (6,7). In contrast, adaptive cytokine responses to influenza infection in swine have been less frequently reported. The adaptive cytokines are important in the activation of lymphocytes and cells of the innate immune system, proliferation of T and B cells, cell differentiation, and immune regulation (29). The V/C pigs in the current study demonstrated significantly elevated levels of IL-2, IL-4, IL-12p70, IFN-γ, and IL-10 protein in the lung at 5 dpi compared to the NV/C and NV/NC groups. Most notable was the increased IL-2 and significantly elevated IFN-γ concentrations at 24 and 48 hours post infection in the MN08 vaccine-primed pigs. This cytokine profile suggests T cell priming in the immunized pigs, and is consistent with the presence of IL-2 and IFN-γ secreting CD4+CD8+ memory T cells in swine (3) and with the rapid increase in total antibody against the heterologous challenge virus in the vaccine primed pigs. An influenza-specific, cross-reactive CD4+CD8+ memory T cell population was detected in MN08 vaccine-primed pigs prior to challenge and reported previously by our group (28). In the same study, a significant increase in cross-reactive CD8+ T-cells was detected by flow cytometry in the V/C pigs post ex vivo stimulation with H1N1pdm09, suggesting CD8+ T-cells were primed by the inactivated vaccine (28). These cytotoxic T-cells may have contributed to the elevated IFN-γ concentrations detected in the V/C post-challenge BALF (3), but also may be associated with the immunopathology in VAERD. Cellular immune mediators in the lung may be responsible for damage to infected tissue as a consequence of viral clearance, as previously reported (2). However, the previous study evaluated pre-challenge T-cell responses in the peripheral blood that may demonstrate a different phenotype than post-challenge T-lymphocytes or central memory CD4+CD8+ T cells that preferentially traffic in lymphoid tissues or pulmonary lymph nodes. The magnitude, quality, timing, and the immune compartment of the CMI response should be considered in future studies in assessing the correlates of protection as opposed to the determinants of VAERD.
Although VAERD-affected pigs were able to respond immunologically to the heterologous challenge without evidence of original antigenic sin, this priming advantage is not likely to outweigh the clinical impact of the severe lung pathology associated with decreased growth and potential losses from secondary infections. Prevention of IAV infection in swine will continue to rely on the use of vaccines; however, vaccine efficacy depends on the ability to cross-protect against antigenically diverse influenza viruses that currently co-circulate in the U.S. and Canada. Therefore, it is critical to understand the effectiveness of vaccine immune responses to heterologous infection and how this response may be improved. There is a real risk of VAERD in swine due to the use of inactivated vaccines and the potential for mismatch of the HA and NA in the circulating strains. Increasing evidence has been reported that a similar clinical outcome may have occurred with some human inactivated vaccine preparations following the emergence of H1N1pdm09 (23,30,32). The model we describe here would be useful for exploring the risk of inactivated human vaccines in inducing VAERD to confirm the epidemiologic links reported in people. Collectively, many aspects of the immune and/or immunopathologic response in VAERD require further investigation to understand the mechanisms associated with vaccine-induced exacerbation of disease or the effects VAERD may have on future infections, either viral or bacterial.
Footnotes
Acknowledgments
The authors thank Michelle Harland and Gwen Nordholm for technical assistance and Brian Pottebaum, Jason Huegel, and Jason Crabtree for assistance with animal studies. Assistance with microbiologic screening from Dr. Susan Brockmeier is appreciated. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. Funding was provided by USDA-ARS. USDA is an equal opportunity provider and employer.
Author Disclosure Statement
No competing financial interests exist.