
Abstract
Hepatitis C virus (HCV) infection is prevalent in patients with maintenance hemodialysis (HD). Although HCV affects the survival rate, antiviral treatment for HD patients is limited. Since impaired innate immunity has been proposed both in hepatitis C and in HD, we compared the immunologic features in periphery between these patients and controls. Thirty subjects were divided into four groups on the basis of HD and HCV infection. In peripheral blood mononuclear cells, NK subsets and their activation status were analyzed by flow cytometry. Cytokine productions were measured both in the culture supernatant and at the single cell level. In HCV-infected HD patients, CD56dim NK subset was decreased (13.1±3.7%, p=0.015) but had upregulated CD69 expression (10.4±4.2%, p=0.032) compared to the other groups. LPS effectively induced neither interferon (IFN)-γ nor tumor necrosis factor (TNF)-α in NK cells of both HCV-infected and -uninfected HD patients, while TNF-α-producing monocytes were increased in HCV-infected HD patients as compared to the uninfected. These findings indicate that chronic HCV infection affects innate immunity independently of HD.
Introduction
It has been reported that ESRD patients have an increased risk for malignancies and infections possibly due to impaired immune responses(21,31), together with the attenuated natural killer (NK) cytotoxicity (9,19,43). NK cells play an important role in viral infections (6). Khakoo et al. (27) reported the correlation between spontaneous HCV clearance and haplotypes of the killer cell immunoglobulin-like receptors. In chronic hepatitis C patients, the impaired killing activity of NK cells (11) and the reduced frequency of CD56dim NK (22,33) have been reported. In vitro studies have shown that HCV envelope protein inhibited NK cell function via the cross-linking of CD81 (12,40).
Even with the piling up of reports, little is clarified about the immunologic effect of HCV in ESRD. Since HD patients were considered as a representative subpopulation of ESRD patients, in this study, we examined the peripheral NK status in HCV-infected HD and in corresponding control groups.
Materials and Methods
Study subjects
A total of 30 subjects were enrolled and categorized into four groups: HCV-infected HD patients (Group HC, n=7), HCV-uninfected HD patients (Group HN, n=7), treatment naïve uncomplicated hepatitis C patients (Group C, n=8), and healthy volunteers (Group N, n=8). There was no significant difference in age, sex, or HCV RNA levels between the groups except for the higher serum ALT in group C. Causative diseases for ESRD were chronic glomerulonephritis (five patients), diabetic nephropathy (three patients), polycystic kidney (one patient), IgA nephropathy (one patient), and unknown origin (three patients). HCV infection was confirmed by the coexistence of anti-HCV antibody (Third generation EIA test) and HCV RNA (Amplicor PCR, Roche Diagnostics, Tokyo, Japan) in the serum (Table 1). Patients with hepatitis B virus infection, human immunodeficiency virus infection, malignancies, or the use of immuno-suppressive agents were excluded. We obtained written informed consent from all subjects prior to the study. This work was performed according to the Declaration of Helsinki, and was approved by the ethical committee of Tohoku University (registration number, 2006-194).
Median and range are shown in age, serum ALT and HCV-RNA.
DM, diabetic nephropathy; GN, glomerulonephritis; N/A, not applicable.
One patient exceeded the upper detection limit; N/A, not applicable.
Cell preparation and culture
Peripheral blood was taken just before periodic HD and isolated to obtain peripheral blood mononuclear cells (PBMC) by Ficoll-Paque PLUS (GE Healthcare, Uppsala, Sweden) density gradient centrifugation. Trypan Blue staining confirmed the cell viability greater than 95% prior to experiments. PBMC were suspended at 2×106/mL in RPMI 1640 medium supplemented with 5% fetal bovine serum, 50 IU/mL penicillin and 50 μg/mL streptomycin, (all from Invitrogen, Carlsbad, CA) and cultured in the presence of 1 μg/mL purified mouse anti-human CD16 antibodies (clone 3G8; BD Biosciences, San Diego, CA) or 1 μg/mL lipopolysaccharide (LPS; Sigma, St. Louis, MO) for 16 hours using 24-well culture plate in a 5% CO2 incubator at 37°C.
Analysis of cell surface antigens
Freshly isolated or cultured cells were suspended in the staining buffer (PBS containing 0.5% bovine serum albumin and 0.09% sodium azide) at 5×105/mL. Cells were stained with various combinations of FITC-anti-CD3, FITC-anti-CD4, FITC-anti-CD16, PE-anti-CD69 (all from BD Biosciences), and APC-anti-CD56 (Miltenyi Biotec, Auburm, CA). The threshold of antigen expression was defined by FMO (full minus one staining) control and isotype-matched antibodies. All samples were run on FACSCalibur (BD Biosciences) and data were analyzed by FlowJo ver. 8 for Mac OSX (Tree Star, Ashland, OR).
Cytokine quantification by ELISA
Interferon (IFN)-γ, tumor necrosis factor (TNF)-α, and interleukin (IL)-10 were quantified by using commercial enzyme-linked immunosorbent assay (ELISA) kits purchased from Biosource International (Camarillo, CA) according to the manufacturer's protocols.
Intracellular cytokine staining (ICS)
For cytokine accumulation in cells, Golgi-Plug (BD Biosciences) was added in the last 6 hours of the culture. Cells were washed twice with PBS and stained with PerCP-anti-CD3 (BD Biosciences) and APC-anti-CD56, followed by the treatment of cell membrane with CytoFix/CytoPerm (BD Biosciences). After the staining with FITC-anti-IFN-γ and PE-anti-TNF-α (both from BD Biosciences) for 30 min on ice, cells were washed again and fixed with 1% paraformaldehyde. For monocyte experiments, APC-anti-CD14 and PE-anti-IL-10 (both from BD Biosciences) were used instead.
Statistical analyses
Paired-t test was used to compare data in each patients/donors with different stimulation, while one-way ANOVA analysis was used to compare between patient groups. P values less than 0.05 were defined as significant. A mean±SEM was shown in the text and figures unless otherwise indicated.
Results
Identification of NK cell fractions in PBMC
Representative FACS dot plots are shown in Figure 1. After gating lymphocyte fraction by forward scatter and side scatter, expressions of CD3 and CD56 were displayed (Fig. 1A). CD3-CD56+ cells were defined as NK cells, and were further divided into two subpopulations, CD56bright and CD56dim cells, showing the different CD16 expression (Fig. 1B).
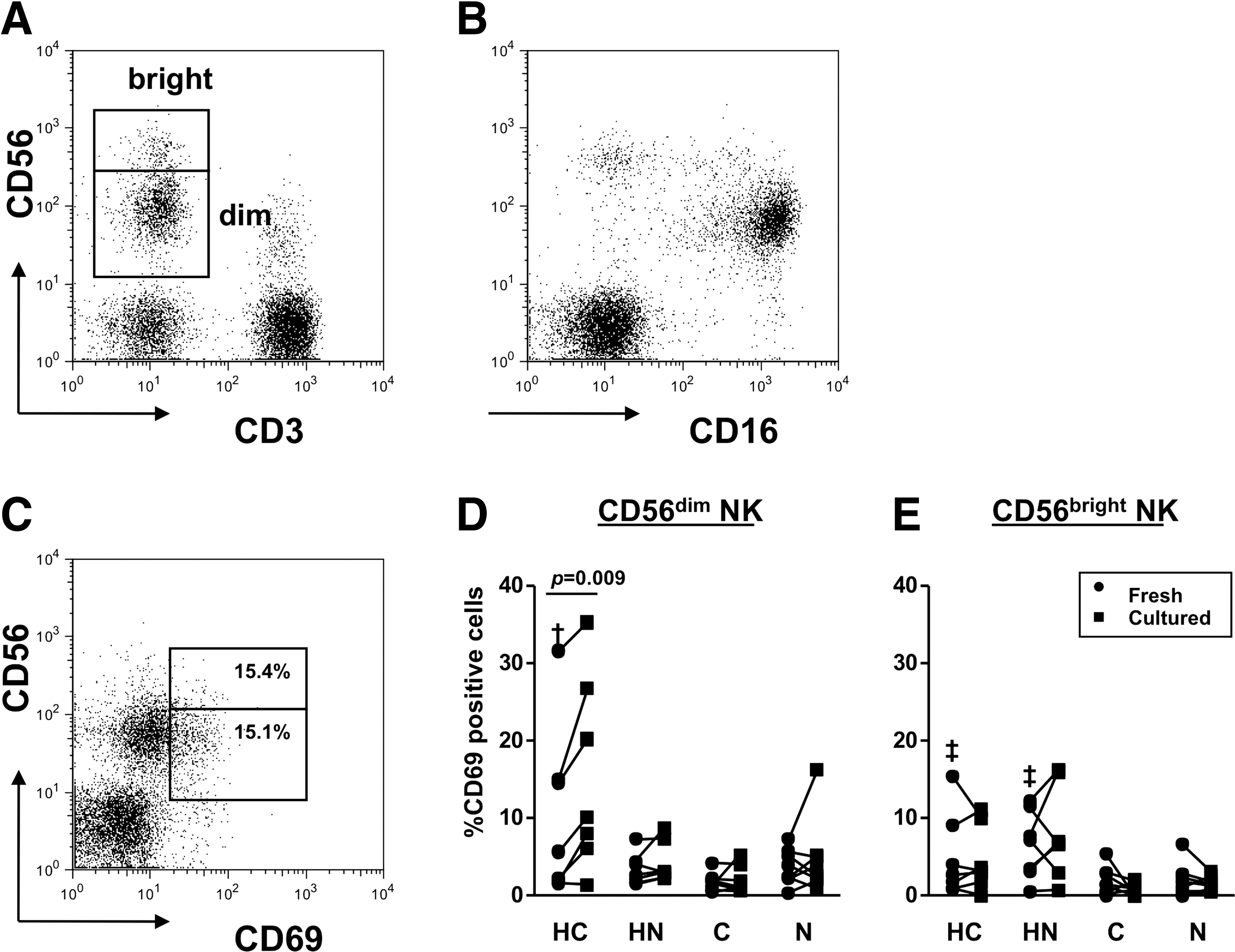
Identification of NK cell subsets and their activation status. Freshly isolated PBMC were stained and run on FACSCalibur.
Altered balance of NK subsets in HCV-infected HD patients
Table 2 shows the frequencies of NK population in PBMC. CD56dim NK composed the majority in total NK in all cases. Total NK and CD56dim NK were significantly decreased in group HC. When we looked CD56bright/CD56dim ratio, HCV-infected groups (n=15), including groups HC and C, showed greater ratio (0.102±0.034) than uninfected groups (n=15; 0.029±0.006, p=0.048).
N.S., not significant.
NK cells are activated in HCV-infected HD patients
The CD69 expression was examined for NK activation. Prominent CD56dim NK activation was found in group HC compared to the other groups (p=0.032), and the intensity increased after the culture (p=0.009, Fig. 1D). On the other hand, the higher CD56bright NK activation was seen in HD patients, including groups HC and HN patients (5.75±1.28%) than in non-HD group patients (1.89±0.48%, p=0.006, Fig. 1E).
To examine NK activation by known stimulatory pathways, we used LPS (a ligand of toll-like receptor 4) and anti-CD16 to stimulate NK cells. Interestingly, the upregulation of CD69 in CD56dim NK was still apparent in group HC both by LPS (p=0.001) and by anti-CD16 (p=0.005) compared to the other groups (Fig. 2A). On the other hand, anti-CD16 showed the limited stimulation on CD56bright NK, probably due to less CD16 expression on the cell surface (Fig. 1B).
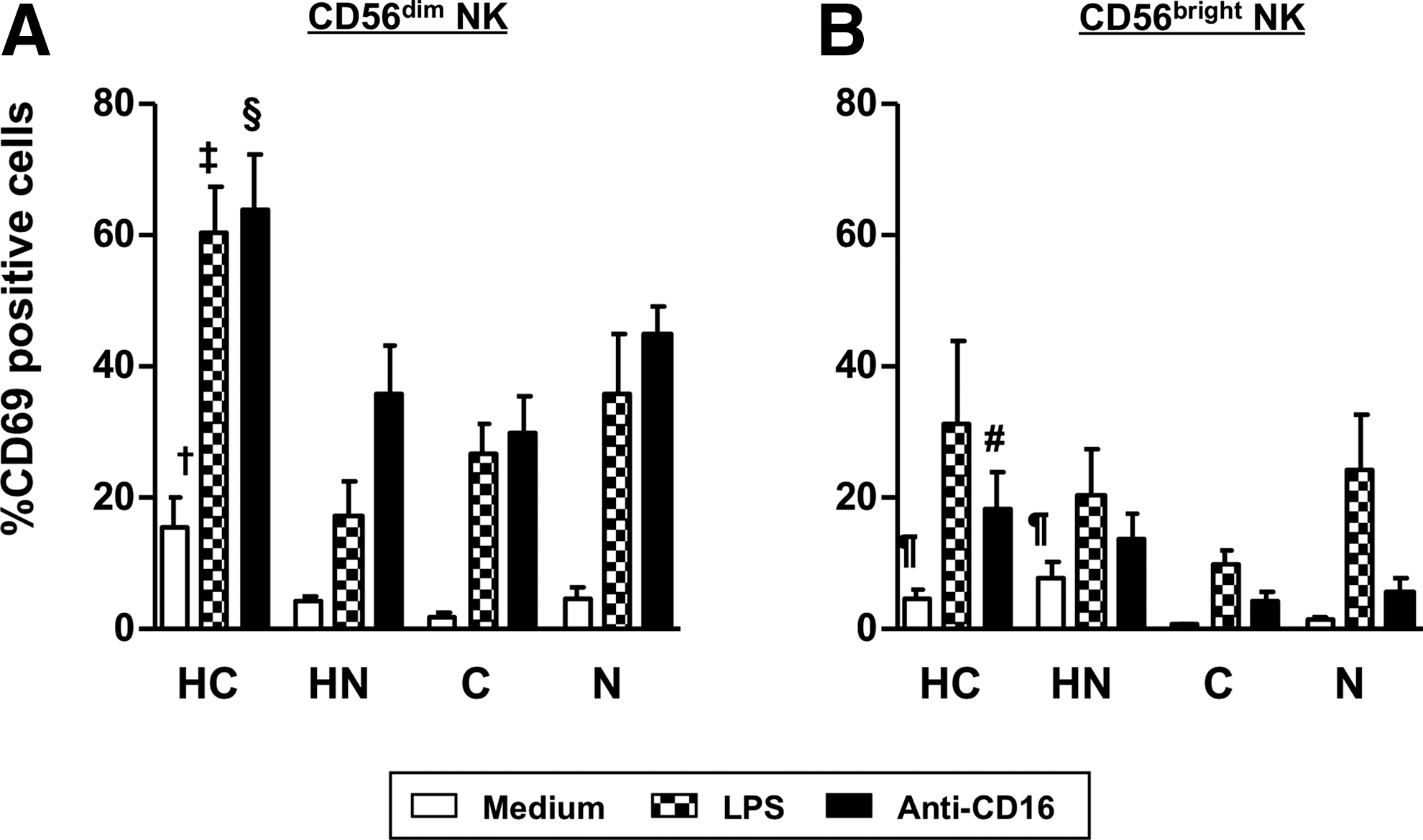
Induction of NK activation by LPS and anti-CD16. PBMC were cultured either with LPS or with anti-CD16, and CD69 expression of cell surface was analyzed.
Reduced IFN-γ production in HD patients
The culture supernatant was collected and the concentration of cytokines was measured by ELISA (Table 3). The production of IFN-γ remained at low level in HD groups, including groups HC and HN, independent of HCV infection except for that with LPS stimulation. The production of TNF-α and IL-10 was significantly upregulated by the stimuli, although there was no difference between groups.
p<0.001, ‡ p<0.05: Compared to medium control.
N.S., not significant.
LPS induces TNF-α production not in NK cells but in monocytes
Next, we perfomed ICS to visualize the source of these cytokines. Since LPS also stimulates monocyte, we analyzed the synthesis of IFN-γ and TNF-α in NK cells, together with TNF-α and IL-10 in monocytes. IFN-γ-positive NK cells were not upregulated either in group HC or in group HN (Fig. 3B), whereas TNF-α-positive NK cells were downregulated in group HN (Fig. 3C). LPS strongly upregulated TNF-α-positive monocytes in group HC than in group HN (p=0.019, Fig. 3D), while the induction of IL-10-positive monocytes was independent of HCV infection (Fig. 3E).
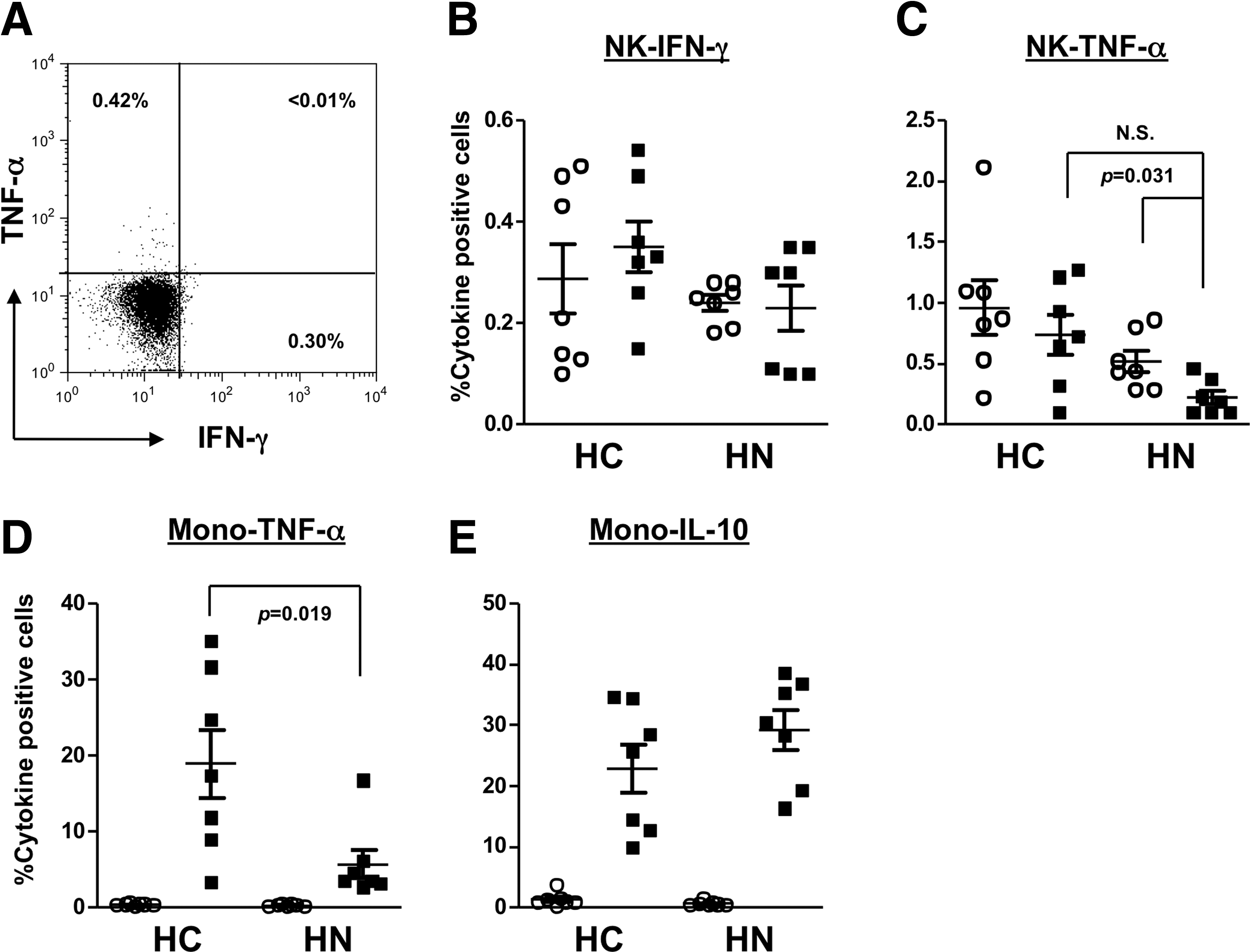
Intracellular cytokine staining of NK and monocytes in HD patients. PBMC were cultured for 16 hours with cytokine accumulation in the last 6 hours. After the staining of surface markers, the cell membrane was permeabilized and cytoplasmic cytokines were stained.
Discussion
NK cells are enriched in the liver and are suggested to play an important role in the pathogenesis of liver diseases (14,36). In humans, two NK subsets, CD56dim NK and CD56bright NK, have been described (10,29). Most of CD56dim NK co-express CD16, also known as Fcγ receptor III, and show strong cytotoxicity either directly or by antibody-dependent cellular cytotoxicity (ADCC), while CD56bright NK produce high levels of cytokines upon stimuli (10).
In this study, first we observed the decreased frequencies of total NK and CD56dim NK in HCV-infected HD patients and the increased CD56bright/CD56dim ratio in HCV-infected patients in accordance with previous reports (22,33). In HCV-infected HD patients, interestingly, the frequency of CD69-positive, “activated”, CD56dim NK cells was much higher and further upregulated after the culture with or even without stimuli. Since CD69 has been reported as a stimulatory receptor (7), which is related to apoptosis(32), CD56dim NK in the periphery might be terminally differentiated phenotype (10) and might undergo irreversible activation in these patients.
NK cells produce IFN-γ and TNF-α to control viral infection, while their proliferation can be inhibited by IL-10 signal. Our ELISA data showed that the baseline IFN-γ production was lower in HD patients and was still at a low level after anti-CD16 stimulation, suggesting some discrepancy between the activation and the cytokine production of NK cells in these patients. On the other hand, the production of TNF-α and IL-10 did not differ between the groups. Although LPS induced TNF-α production, ICS data showed that it did not upregulate TNF-α-producing NK cells but upregulated TNF-α-producing monocytes in HCV-infected HD patients compared to the uninfected. It has been reported that hepatitis C patients showed higher serum TNF-α, which is related to the augmented liver inflammation (35,44) and monocytes of HCV infected patients produced higher levels of both TNF-α and IL-10 in response to the stimulation by either LPS or HCV proteins than that of healthy controls (15).
Recently, Ahlenstiel et al. (1) reported that NK cells of chronic hepatitis C patients expressed higher levels of TNF-related apoptosis inducing ligand and NKp46 predominantly in the liver correlating with CD107a expression, a marker of degranulation, while IFN-γ production was not induced. Upregulation of CD107a NK cells with decreased IFN-γ and TNF-α production was also reported by Oliviero et al. (37), suggesting the role of NK cells in liver inflammation and disease progression in chronic hepatitis C and the importance of IFN-γ for viral clearance. Although we did not perform a degranulation assay, the NK cell status in HCV-infected HD patients might be similar because reduced IFN-γ level and no upregulation of IFN-α-producing NK were described contrary to their activation.
NK cells are also known to work in adaptive responses via the crosstalk between dendritic cells (20), monocytes/machropages (34), and regulatory T cells (Treg) (39), which has also been highlighted in HCV infection (2). Since the impaired T cell activity has been reported in HD (13), we examined T cell function using enzyme-linked immunospot assay to quantify antigen-specific T cells. The frequencies of HCV-specific T cells, against core, NS3, NS4, and NS5, were independent on undergoing hemodialysis (data not shown). Treg modify the pathogenesis of viral infection by inhibiting activated T cells either via the direct cell to cell contact or via suppressive cytokines (3). We could not find any difference of frequencies of Treg, identified by the feature of CD4+CD25+CD127low/-, between the groups (data not shown).
There are still some basic questions, that are whether HD and/or dialyzer itself modifies peripheral immune status and whether primary disease for ESRD is related. It might be answered if results are campared to the data from samples taken just after HD and the control group for ESRD without HD are set properly. However, it seems difficult due to the wide distribution of renal function (serum creatinine). Because HCV infection associates cryoglobulinemia and glomerulonephritis, in some patients (16,26), the effect of HCV infection and that of ESRD/HD cannot be separated strictly. In this regard, our study is inadequate to conclude an overview of NK status, but is still the first report to show the decreased frequency and the activation of CD56dim NK cells in HD patients. These immune disorders may give some disadvantage for the pathophysiology and the treatment of HCV, although whether HCV has direct effect on NK cells is still controversial, even using cell culture generated HCV or HCV pseudo-particles (18,41,42). To clarify the real impact of HCV on immune responses in such subpopulation, large-sized studies that match HCV genotype and etiology of ESRD are necessary.
Footnotes
Acknowledgments
The authors thank Drs. Tetsuya Ootaka, Yoichi Ohashi (Katta General Hospital), Takaomi Sato (Kojinkai Kimachi Hospital), and Tomoo Kobayashi (Tohoku Rosai Hospital) for sample arrangement, Dr. Toshinobu Sato (Tohoku University) for special advice, and Mr. Satonori Kon for technical assistance. This work was supported by the grant of Ministry of Health, Labour and Welfare of Japan (H18-hepatitis-general-002 to KK).
Author Disclosure Statement
All authors declare that there are no competing financial interests.