
Abstract
Porcine reproductive and respiratory syndrome (PRRS) has become one of the most economically important diseases to the global pork industry. Currently, the efficacies of available commercial vaccines remain questionable: the modified live-PRRSV vaccines (MLVs) were generally effective but variable in sufficient protection, and the outcomes of inactivated-PRRSV vaccines (IVs) in the field were not very promising. In the present study, we investigated the effect of swine interleukin 4 (IL-4) on the development of virus-specific immune responses elicited by an MLV. The antibody titer against PRRSV membrane proteins in pigs elicited by MLV plus recombinant plasmid encoding IL-4 (group 3) was significantly higher than those elicited by MLV alone (group 1) and MLV plus empty plasmid (group 2) from 35 days post-inoculation (dpi). Similarly, the neutralizing efficacy of sera from group 3 was markedly enhanced compared with group 1 and group 2. In cellular immunity, the ratio of CD3+CD4+/CD3+CD8+ T lymphocyte subpopulations from group 3 monitored by flow cytometry (FCM) was significantly higher than those from group1 and group 2 from 42 dpi to 21 days post-challenge (dpc). After viral challenge, pigs in group 3 showed significantly lower virus loads in peripheral blood measured by a real-time quantitative PCR (RT-qPCR), as compared with those in group 1 and group 2. Pigs in group 1 and group 2 had a low fever and displayed mild inappetence, lethargy, rough hair coats, and no lung lesions, while those in group 3 showed almost no clinical signs, no lung lesions. The scores of clinical signs of pigs in group 3 were significantly lower than those in both group 1 and group 2. Interestingly, the scores of lung lesions showed no significant differences among the three groups. Our results indicate that swine IL-4 markedly enhanced the protective immune response of pigs and improved the efficacy of the MLV in preventing PRRS disease.
Introduction
P
In the early stage of infection, a vigorous anti-PRRSV antibody response can be detected starting at 7–9 days post-infection. Non-neutralizing antibodies (Non-NAs) are generated abundantly in hosts, which do not prevent the virus infection. On the contrary, the Non-NAs might be responsible for antibody-dependent enhancement (ADE) of viral replication (20,39). Neutralizing antibody (NA) plays a critical role in preventing PRRSV infection in pigs (29), similarly as the typical equine arteritis virus (EAV) (5). However, NAs specific to PRRSV have a very late onset that can be detected about 28 days post-infection or inoculation and maintain at a low level (11,24,29).
Inoculation with vaccine is still the best method to prevent and control PRRS. To date, there are mainly two types of PRRSV vaccine commercially available, MLV and IV. MLV is effective to a certain extent but cannot provide sufficient protection to pigs. The antibody responses to MLV appear within 14 dpi, while neutralizing antibodies arise at 4 weeks post-inoculation and sustain at a low level. Vaccine-induced cell-mediated immunity (CMI) appears at a low level at 14 dpi and develops slowly (7,13,26,28). The efficacy of IV is not ideal, which induces poor CMI responses and does not induce an antibody response (measured by an IDEXX ELISA) (31,40). Thus, it is important to develop measures for elevating the neutralizing antibody titer produced by pigs inoculated with the commercial MLV or IV, extending the time period of high antibody titer and enhancing CMI. An important approach for enhancing immune responses of hosts by vaccines is the use of adjuvant. Recently, the commonly utilized adjuvants include cytokines (IL-1, IL-2, IL-6, IL-12, IFN-γ, IFN-α, CD40), bacterial products (CT, CpG ODN, HPS70), and chemical agents (LMS, Poly IC). They were used as nucleotides or proteins directed to different antigens of PRRSV including ORF5 DNA, ORF7 DNA, Ingevac MLV, In-house IV, Chinese commercial IV, recombinant N protein, and synthetic GP5 peptides (2,4,10,16,22,23,32,36,38). However, using swine IL-4 as an adjuvant to elevate immune responses against PRRSV has not been reported before.
To understand the character of immune responses of pigs infected with virus or inoculated with vaccine, we focused on swine IL-4 (Th2-type cytokine) as a vaccine adjuvant in this study. IL-4 is a central regulator of T helper 2 (Th2) immune response (8), and was first described as a cofactor in the proliferation of resting B-cells (1,30). Previous studies have demonstrated that IL-4 can promote the differentiation of T helper cells into Th2 cells during an immune response and thus enhance vaccine efficacy (30). IL-4 has been used as an adjuvant in experimental human vaccines. It significantly enhanced antigen-specific humoral immune responses in the rhesus macaques inoculated with human immunodeficiency virus (HIV) Env/Rev and simian immunodeficiency virus (SIV) Gag/Pol proteins (18). Recombinant plasmid encoding swine IL-4 induced a stronger immune response to Cysticercosis cellulosae vaccine than that of vaccine alone in mice and pigs (15).
To gain a better understanding of the variation in the immune response and protection efficacy, we tested the effect of swine IL-4 in the context of MLV-induced immune responses of pigs for PRRSV.
Materials and Methods
Virus, vaccine, and cell lines
Highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) strain SD-JN (GenBank accession No. FJ422123), the challenge virus used in this study, was isolated from an aborted fetus in Shandong province of China, stored at−70°C, and described elsewhere (37), which was determined to have 89.4% homology with the published PRRSV American isolate VR2332 (GenBank accession No. PRU87392). The virus was propagated in MARC-145 cells (17), which were cultured in Dulbecco's modified Eagle's medium (DMEM, Gibco BRL, Grand Island, NY) media containing 10% fetal bovine serum (FBS) (Gibco BRL). HEK-293A cells (ATCC CRL1573) were used for eukaryotic transfection and cultured in DMEM. The vaccine used to inoculate pigs was a MLV which contained PRRSV strain TJM-F92 (attenuated from the TJM strain, GenBank accession No. HQ679913) (21) produced by the Yebio (Yebio Co., Ltd., Qingdao, China).
Cytokine gene clone and expression vector construction
Swine IL-4 was cloned from the total RNA isolated from swine lymphocytes in peripheral blood mononuclear cells (PBMCs) using TRIzol LS (Invitrogen Life Technologies, Carlsbad, CA) following the manufacturer's recommendations. Cells were stimulated in vitro with 10 mg/mL Concanavalin A. The extracted RNA was used as a template for RT-PCR reaction by reverse transcriptase (Primescript® one step RT-PCR kit, Takara, Dalian, China) with specific primers (IL-4 F: 5′-AACTGCAGACCATGGAAGGTCTCACCTCCCA-3′, IL-4 R:5′-CGGCCTCGAGTCAACACTTTGAGTATTTC-3′) in which a Kozak sequence (19) was plugged in ahead of the latter.
The coupled IL-4 cDNA was cloned into the eukaryotic expression vector pVAX1 vector plasmid (Invitrogen) through the restriction enzymes PstI and XhoI (NEB, Ipswich, MA). The gene insert of the construct, named rpVAX1-SwIL-4, was sequenced and analyzed using the universal primer M13R-48 by Roche 454 GS FLX+system in BGI (BGI Co., Ltd., Beijing, China).
In vitro transient expression
Immunofluorescence assay (IFA)
In each well of a 96-well culture plate, approximately 5×104 HEK-293A cells were seeded in 100 μL of DMEM media containing 10% FBS. About 24 h later (the confluency for adherent cells is 70%–90%), the rpVAX1-SwIL-4 plasmids were transfected into the 293A cells using the TurboFect reagent (Thermo Scientific, Barrington, IL). The protocol was as follows: 1 μg of plasmid DNA was diluted in 100 μL of serum-free DMEM, then 2 μL of briefly vortexed TurboFect reagent was added to the diluted DNA. The cocktail was mixed immediately by pipetting or vortexing and incubated for 15–20 min at room temperature. Then 100 μL of the TurboFect/DNA mixture was added dropwise to each well, and incubated at 37°C in a 5% CO2 incubator.
The recombinant protein was analyzed 24 h later. The transfected HEK-293A cells were rinsed with PBS and fixed with cold ethanol for 30 min at 4°C, then washed and air-dried in a biosafety cabinet. The cells were incubated with 0.5 μg/mL monoclonal mouse anti-porcine IL-4 IgG1 (R&D Systems®, Inc., Minneapolis, MN) for 1 h at 37°C. After washing with PBS, the cells were incubated with goat anti-mouse IgG (H+L) conjugated with fluorescein (1:200 diluted in PBS) (Alexa Fluor, Invitrogen) for 1 h at 37°C in the dark. After rinsing five times, the cells were observed under a ZEISS fluorescence microscope.
Western blot analysis (WB)
Western blots were used to evaluate protein's expression by rpVAX1-SwIL-4. Briefly, the culture medium from HEK-293A cells transfected with rpVAX1-SwIL-4 plasmid DNA in a 24-well plate was decanted carefully. The cells were washed once in wash buffer PBS. Then, 200 μL mammalian protein extraction reagent (M-PER®, Thermo Scientific, Rockford, IL) was mixed with protease inhibitor cocktail (Pierce Halt®, Rockford, IL) and added into each well, shaking gently for 5 min. The lysates of HEK-293A cells were collected and centrifuged at 14,000 g for 10 min at 4°C. The supernatant was boiled in SDS-PAGE loading buffer (60 mM Tris-HCl (PH 6.8), 2% SDS, 0.1% bromphenol blue, 25% glycerol, 5% β-mercaptoethanol) for 5 min. The target proteins were separated by 10% SDS-PAGE and transferred to polyvinylidene difluoride (PVDF) membranes (Millipore, Billerica, MA) through the semiphor transphor unit (BioRad, Hercules, CA). The membrane was blocked with PBS Tween-20 containing 2.5% nonfat dry milk at 4°C over night, and then incubated with 2 μg/mL monoclonal mouse anti-porcine IL-4 IgG1 (R&D Systems®, Inc., Minneapolis, MN) for 2 h at room temperature, followed by incubation for 2 h at room temperature with horseradish peroxidase (HRP) conjugated goat anti-mouse IgG (1:5000 diluted in PBS) (Jackson ImmunoResearch, West Grove, PA). Detection was performed using the Super signal West Pico Chemiluminescent Substrate according to the manufacturer's suggestions (Pierce, Rockford, IL).
Vaccination and viral challenge protocol
Twenty-four PRRSV-free (determined by RT-PCR, using a pair of primers: 5′-TTTGGGGAGTGTACTCGGCCATAGA-3′, 5′-GGCATATTTGACAAGGTTTACCACT-3′), antibody-negative (tested by PRRSV ELISA Kit (IDEXX Laboratories, Inc., Westbrook, MA)), 3-week-old, healthy crossbred Yorkshire pigs were randomly assigned to four experimental groups of six animals each.
Vaccination was administered intramuscularly in the cervical region as follows. At day 1, group 1 received the MLV containing 105.0 TCID50 PRRSV TJM-F92 strain in 2 mL of volume, group 2 received a same dose of the MLV plus 500 μg empty pVAX1 plasmid in a total 3 mL of volume, group 3 received a same dose of the MLV plus 500 μg rpVAX1-SwIL-4 plasmid in a total 3 mL of volume, and group 4 received 3 mL PBS that served as an unvaccinated negative control. Plasmids and PBS used to inoculate pigs were filtrated with 0.22 μm filter (Millipore, Bedford, MA). Blood samples were drawn at 0, 14, 28, 35, 42, 56, and 63 dpi. At 63 dpi, all experimental animals were challenged intranasally with 2×105.0 TCID50 of the HP-PRRSV strain SD-JN and monitored daily for 21 days. After viral challenge, blood samples were taken at 7-day intervals, and all pigs were euthanized at 21 dpc.
In vitro detection of IL-4
The concentration of swine IL-4 in serum samples from pigs of each group was determined by a commercial ELISA kit (Invitrogen™, Camarillo, CA) following the manufacturer's instructions. The concentration of IL-4 in each sample was calculated from the corresponding OD450 value based on an established standard curve.
Antibody detection
Antibodies specific to the membrane proteins in PRRSV (including GP2, GP3, GP4, and GP5) in pig sera in various time-points (0, 14, 28, 35, 42, 56, 63 dpi, and 0, 7, 14, 21 dpc) were tested using the LSIVET SUIS PRRS A/S kit (Laboratoire service international, Lissieu, France) as described previously (9), a commercially available indirect enzyme-linked immunosorbent assay (iELISA) with a sensitivity and specificity of 90.6% and 98.3%, respectively, according to the manufacturer. All tests were performed according to the manufacturer's instructions and results based on the IRPC (relative index×100) of the sample with a cut off of >20 determining seropositivity, where: IRPC=(OD450 Sample – Mean OD450 Negative control)/(Mean OD450 Positive control – Mean OD450 Negative control)×100. The sera of pigs were diluted serially (1:200, 1:400, 1:800, to 1:25600) in sample diluents and measured by the kit. The antibody titer was expressed as the highest dilution of serum when the corresponding IRPC value was more than 20.
Serum neutralization (SN) assay
SN assay was performed as previously described (14). Briefly, serum samples from pigs were inactivated at 65°C for 30 min, and then they were applied in two-fold dilution series and mixed with an equal volume of HP-PRRSV strain SD-JN containing 200 TCID50. After incubation at 37°C for 1 h, the mixtures were transferred to a MARC-145 monolayer in quadruplicate in a 96-well tissue culture plate. Then the plate was incubated at 37°C in a 5% CO2 incubator and observed daily for up to 5 days for the appearance of cytopathic effects (CPEs). The titers of neutralizing antibody were expressed as the reciprocal of the highest serum dilution in which no CPE was observed. Meanwhile, porcine antiserum against HP-PRRSV was used as a positive control.
Flow cytometry (FCM)
At 0, 14, 42, 63 dpi and 0, 14, 21 dpc, serum samples of all pigs were collected and suspended in EDTA-K2 of BD vacutainer (BD, Franklin Lakes, NJ). 100 μL anticoagulant sera, which the cell number was determined at a concentration of 106 cells per test, were mixed with PerCP-Cy5-5- labeled mouse anti-pig CD3ɛ IgG1, FITC-labeled mouse anti-pig CD4a IgG2b, and PE-labeled mouse anti-pig CD8a IgG2a (BD Pharmingen™, Franklin Lakes, NJ) and incubated in dark for 20 min. Then 600 μL formic acid of 1.2 ml/L, 265 μL buffer (Na2CO3 6 g/L, NaCl 14.5 g/L, Na2SO4 31.3 g/L), 10 μL paraformaldehyde solution (10 g paraformaldehyde dissolved in 1 L PBS) were added into the mixture orderly and vortex for 30 sec, respectively. Flow cytometry was performed on a FACS Arial™ (BDIS, San Jose, CA).
Real-time quantitative PCR (RT-qPCR)
Real-time quantitative PCR was used to quantify virus loads in the sera of experimentally infected pigs. Two primers specific to HP-PRRSV were designed to amplify a 131-base pairs fragment in the conserved Nsp2 region, which is absent in the vaccine strain TJM-F92, using the DNA software Primer Premier 5.0 (QF: 5′-ACCAGGCGTTTCGCATCT-3′ and QR: 5′-ACTCTCTGCACTCACGGAAGG-3′). The amplified product of target gene was cloned into pMD18-T vector (TaKaRa, Dalian, China). Positive recombinant plasmid was diluted from 1×109 to 1×104 copies/mL as positive quantitative templates to establish a standard curve.
Viral RNA was extracted from 250 μL of sera from all pigs at 0, 3, 7, 14, and 21 dpc with TRIzol LS (Invitrogen Life Technologies) following the manufacturer's protocol, and the RNA pellet was suspended in 20 μL of diethylpyrocarbonate (DEPC)-treated water. A genome DNA eraser reaction and a reverse transcription reaction were performed using the Primescript® RT reagent with gDNA Eraser kit (Takara, Dalian, China). 10 μL of purified RNA with oligo dT primer were reverse transcribed to cDNA. Real-time PCR was performed using 12.5 μL SYBR green I Premix Ex Taq II, 0.4 μM of each primer (QF and QR) and 3 μL of the RT reaction product, following the manufacturer's protocol for SYBR® Premix Ex Taq™ II (Takara, Dalian, China). The samples were heated for 5 min at 95°C and a two-step cycle (15 sec at 95°C and 10 sec at 50°C) was repeated 40 times. The fluorescence signals were collected at the end of a 50°C heating period. Finally, the samples were heated for 2 min at 99°C, 20 sec at 60°C, and 10 min at 95°C, respectively. Virus RNA quantity of the samples was determined by linear extrapolation of the Ct value plotted against the standard curve.
Rectal temperature, clinical signs, and lung lesions after viral challenge
After viral challenge, the rectal temperatures of pigs were measured in the morning and the severity of the clinical signs of all animals was evaluated as reported (35). Observations included behavior, respiration, and cough. Scores for each of three individual observations ranged from 1 to 4. The overall score for clinical condition was determined by sum of daily observations of behavior, respiration, and cough. For example, a clinically normal animal would be given a total score of 3 (i.e., behavior=1, respiration=1, and cough=1), an animal with maximum clinical illness would be given a total score of 9 (i.e., behavior=3, respiration=3, and cough=3), and a dead animal would be given a total score of 12 (i.e., behavior=4, respiration=4, and cough=4). At day 21, the animals were euthanized and the gross lesions of lungs were evaluated at necropsy. Lungs were evaluated by the percentage of lesions noted per lobe, following which, using a standard scoring system, an overall level of gross lung pathology was determined (12).
Statistical analysis
Data were analyzed and the differences were determined by one-way repeated measurement ANOVA and least significant difference (LSD). A p value<0.05 was considered statistically significant. The statistical analyses were performed by the IBM SPSS Statistics 20 software (SPSS Inc., Chicago, IL).
Results
Construction of recombinant cytokine plasmid
As shown in Figure 1, the eukaryotic expression vector pVAX1 containing the swine IL-4 gene and a Kozak sequence (ANNATGG) under the control of a CMV promoter was constructed. The result of sequencing analysis showed that the nucleotide sequence of the insert gene in the recombinant plasmid was the same as the published porcine IL-4 sequence (2) and had the correct open reading frame (data not shown).

Schematic diagrams of the recombinant eukaryotic expression plasmid. The swine interleukin 4 gene and Kozak sequence were cloned into pVAX1 vector through the restrict enzymes Pst I and Xho I tandem in frame.
In vitro transient expression
The recombinant swine IL-4 gene was expressed in mammalian cells. HEK-293A cells were transfected with rpVAX1-SwIL-4 plasmid and detected by Western blot and IFA with monoclonal mouse anti-porcine IL-4 IgG1. Western blot analysis demonstrated that the specific protein band was coincident with the predicted size of porcine IL-4, whereas no specific protein band was found in the control pVAX1 vector-transfected HEK-293A cell lysates (Fig. 2a). Meanwhile, the result of IFA indicated that HEK-293A cells transfected with rpVAX1-SwIL-4 plasmid could react with antibodies specific to porcine IL-4 and be stained with goat anti-mouse IgG-FITC, but the control pVAX1 vector-transfected cells were not stained (Fig. 2b).

Identification of expression of swine IL-4 gene in vitro.
Concentration of IL-4 in serum
The concentrations of IL-4 in serum samples at different time points (0 dpi, 42 dpi, 7 dpc) were detected by a commercial ELISA kit. As shown in Figure 3, the concentrations of IL-4 in the serum samples were markedly decreased after inoculation pigs with MLV alone or with MLV plus pVAX1. In contrast, the concentrations of IL-4 in the sera from pigs inoculated with MLV plus pVAX1-IL-4 were significantly higher than that of the control group of PBS (p<0.05). However, the variation tendency of IL-4 levels in the pig sera from the different treatment groups underwent no obvious change after challenging the pigs with HP-PRRSV.
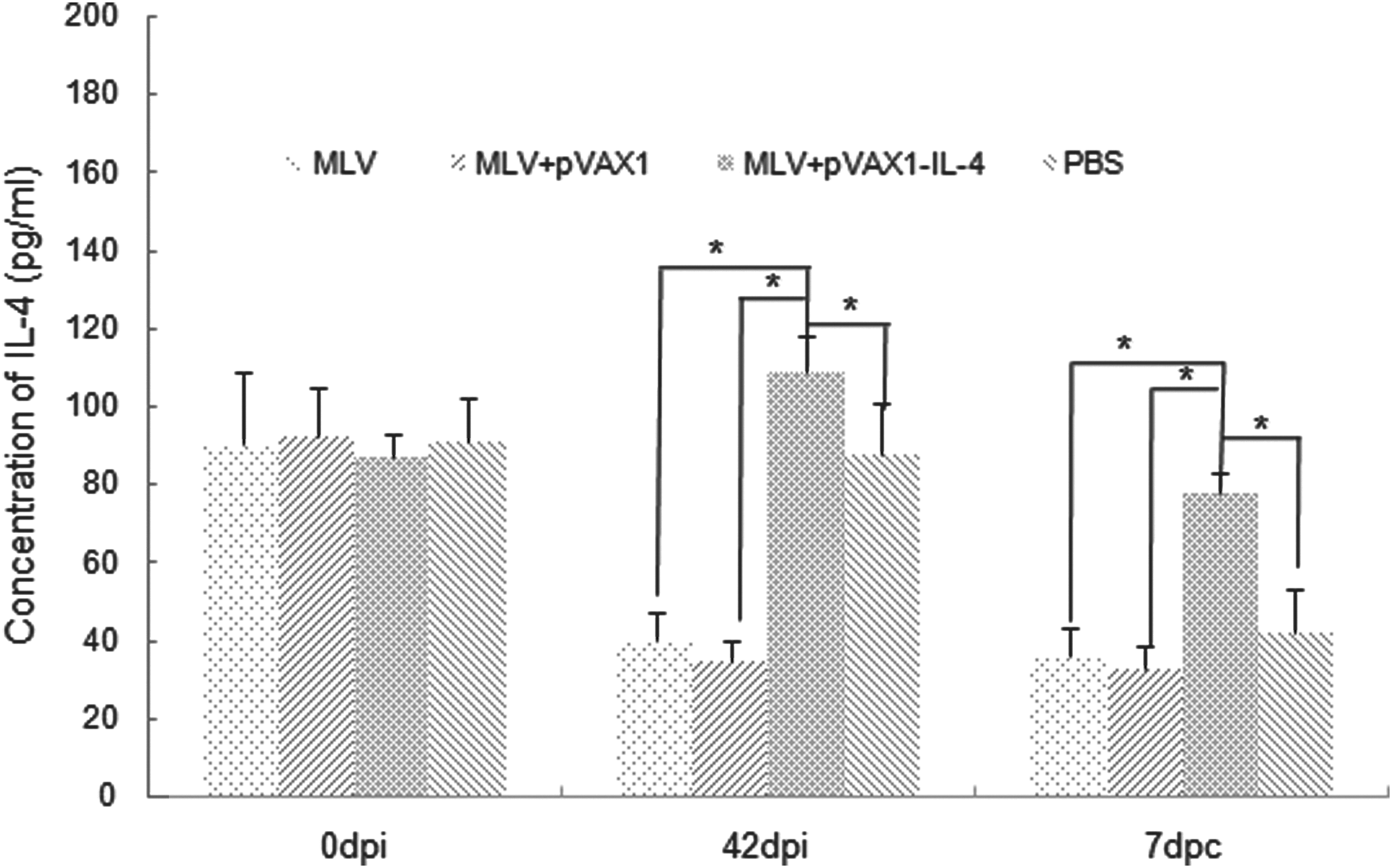
The concentration of swine IL-4 in serum samples from pigs of each groups at different time points (0 dpi, 42 dpi, and 7 dpc) was determined by a commercial ELISA kit. An asterisk indicates that the concentration of IL-4 in one group was significantly higher than that of the other group (p<0.05). Data were shown as mean±S.D.
Humoral immune responses
Swine IL-4 enhanced the humoral responses of pigs inoculated with MLV. Anti-PRRSV membrane proteins antibodies in sera of all pigs in various time-points were measured by the iELISA (Fig. 4). Antibodies could be detected at 14 dpi and kept at a low level, and antibody titers from all pigs vaccinated with MLV plus rpVAX1-SwIL-4, MLV alone and MLV plus pVAX1 markedly increased from 28 dpi onwards. Beginning at 35 dpi, the antibody titers form the group of MLV plus rpVAX1-SwIL-4 were significantly higher than those of MLV alone and MLV plus pVAX1 (p<0.05). After viral challenge, the antibody titers in pig sera from the three groups decreased distinctly at 7 dpc. But the antibodies of pigs from the group of MLV plus rpVAX1-SwIL-4 rebounded to previous levels rapidly and significantly higher than those of MLV alone and MLV plus pVAX1 (p<0.05). On the contrary, the antibodies of pigs from the group of MLV alone and the group of MLV plus pVAX1 were sustained at relative low levels. No PRRSV-specific antibody was detected in the control group of PBS, with exception of a very low level at 14 and 21 dpc.

Humoral immune responses in pigs inoculated with different immunogens. Serum samples (n=6) were collected at various time-points and antibodies specific to PRRSV membrane proteins were detected by iELISA. The antibody titer was expressed as the highest dilution of serum when the corresponding IRPC value was more than 20. An asterisk indicates that the antibody titer of pigs in one group was significantly higher than that of the other group (p<0.05). Data were shown as mean±S.D.
The serum samples of pigs were also evaluated for the ability to neutralize PRRSV in vitro by SN assay. As shown in Fig. 5, the titers of neutralizing antibodies in sera from the group of MLV plus rpVAX1-SwIL-4 were significantly higher than those of MLV alone and MLV plus pVAX1 from 35 dpi till the end of the experiment (p<0.05). While in the primary period after inoculation, the difference of three groups was not significant (p>0.05). The neutralizing titer of sera in the control group of PBS approached to zero throughout the experiment.
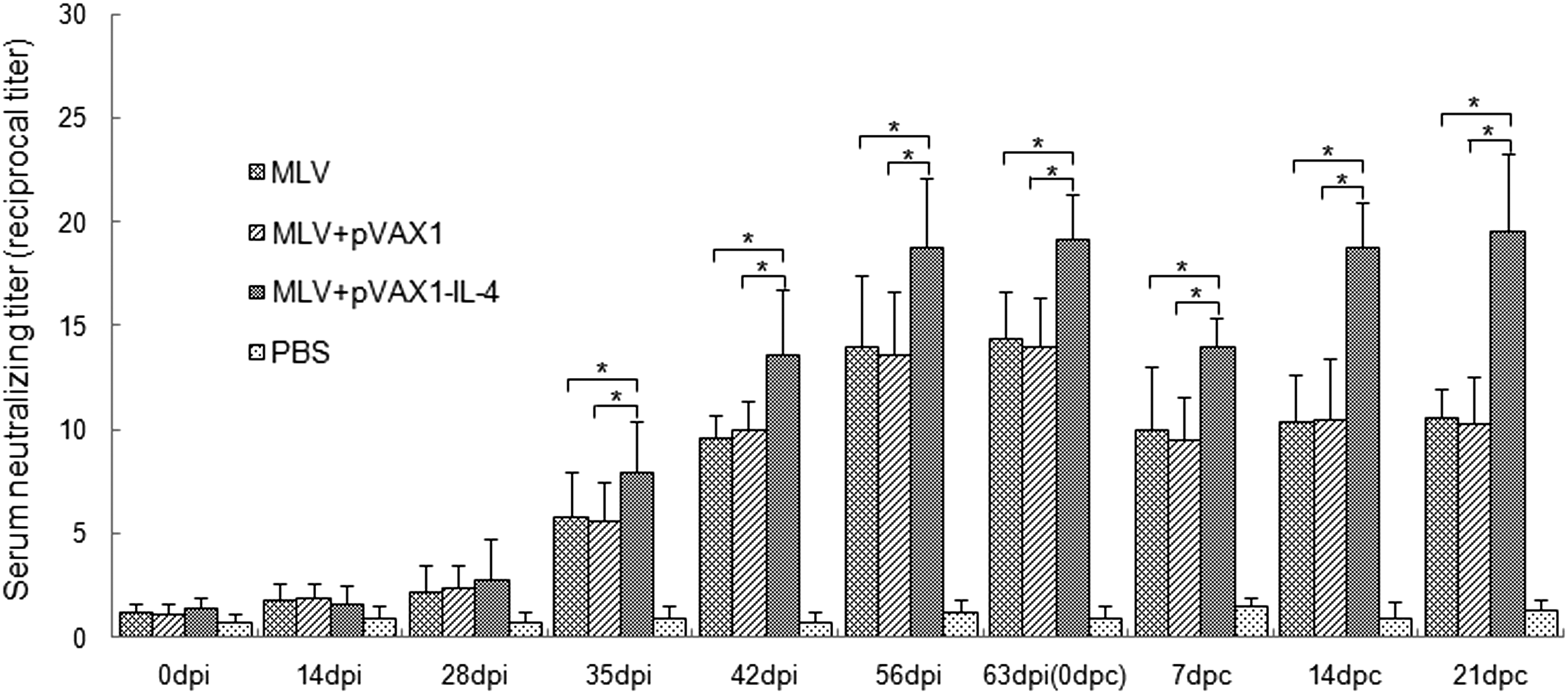
Serum neutralizing titers of pigs inoculated with different immunogens. Serum samples (n=6) were collected at various time-points and neutralizing antibodies specific to PRRSV were evaluated by SN assay in MARC-145 cells. The titer of neutralizing antibodies was expressed as the reciprocal of the highest serum dilution in which no CPE was observed. An asterisk indicates the serum neutralizing titers of both inoculated groups were significantly different (p<0.05). Data were shown as mean±S.D.
CD3+, CD3+CD4+ and CD3+CD8+ T lymphocyte subpopulations
In this experiment, the percentages of CD3+, CD3+CD4+, and CD3+CD8+ T lymphocyte subpopulations were measured by FCM, respectively. The percentage of CD3+ T subpopulations in the group of MLV plus rpVAX1-SwIL-4 markedly increased and was significantly higher than those of MLV alone and MLV plus pVAX1 (p<0.05) in the whole period of 21 days post-challenge. Meanwhile, the percentage of CD3+ T subpopulations in pigs inoculated with PBS also increased at 7 dpc, but it decreased rapidly and had no significant difference as compared with those of MLV alone and MLV plus pVAX1 (Fig. 6a). Pigs inoculated with MLV plus rpVAX1-SwIL-4 showed an enhanced ability to generate CD3+CD4+ T subpopulations, but displayed an inhibited ability to generate CD3+CD8+ T subpopulations especially in the post-challenge period (data not shown). As shown in Figure 6b, the ratio of the CD3+CD4+/CD3+CD8+ T subpopulations in peripheral blood from the group of MLV plus rpVAX1-SwIL-4 was increased from 42 dpi and was maintained at a high level after viral challenge. And the ratio from the group of MLV plus rpVAX1-SwIL-4 was significantly higher than those of the other three groups (p<0.05). On the contrary, the ratio from the groups of MLV alone and MLV plus pVAX1 decreased obviously after viral challenge, but it showed no significant difference compared with the control group of PBS as a whole.
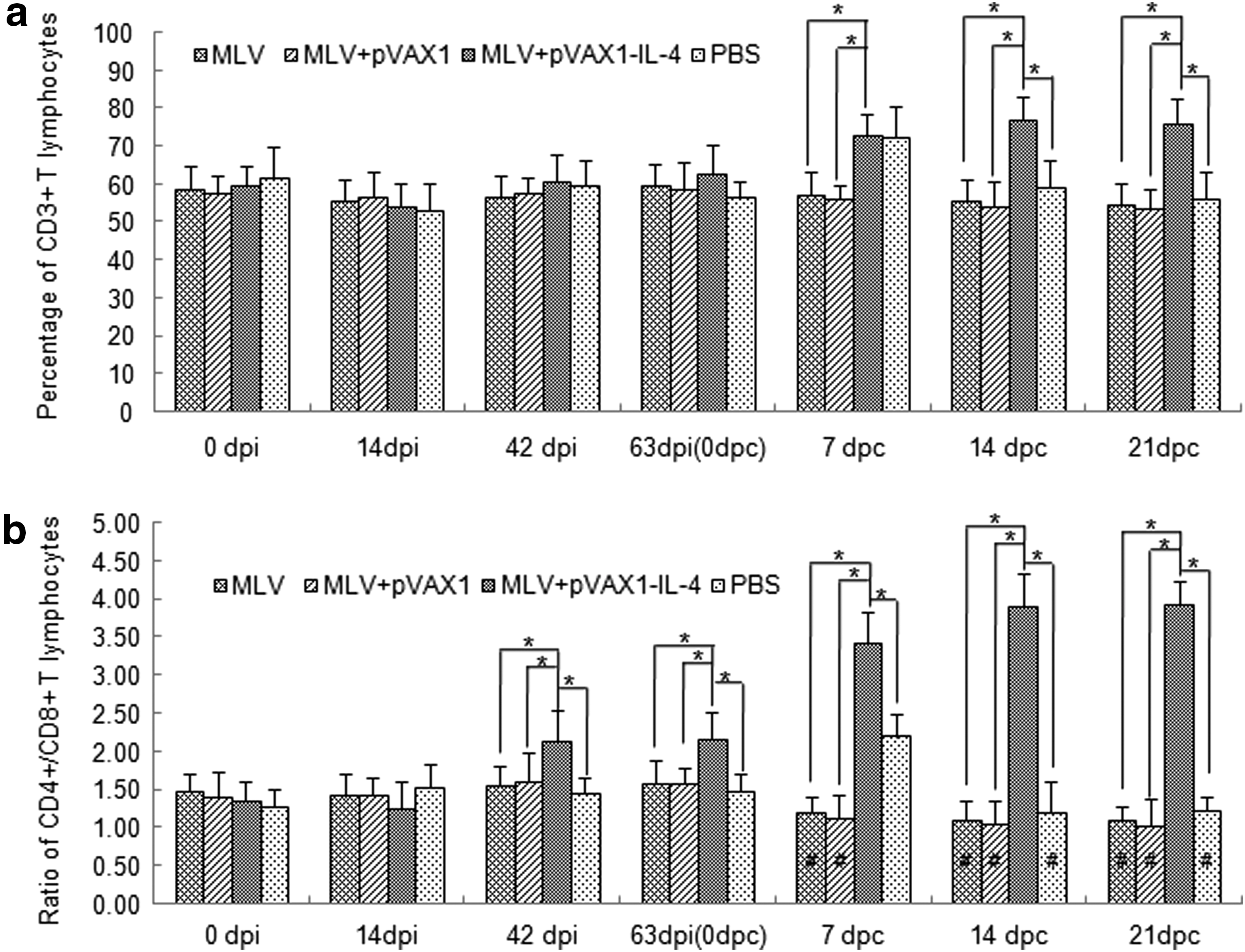
Percentage of CD3+, CD3+CD4+, CD3+CD8+ T lymphocytes, and ratio of CD3+CD4+/CD3+CD8+ T lymphocytes in peripheral blood of pigs. The percentage of CD3+, CD3+CD4+, and CD3+CD8+ T lymphocytes in peripheral blood of pigs from every group (n=6) were monitored by FCM. The ratios of the CD3+CD4+/CD3+CD8+ T subpopulations from all groups were calculated and analyzed statistically. An asterisk indicates the percentage or ratio of both inoculated groups were significantly different (p<0.05). Meanwhile, # p<0.05, as compared to the corresponding group respectively before challenge. Data were shown as mean±S.D.
Virus loads analysis
At 0, 3, 7, 14, and 21 dpc, the blood samples of all pigs were collected and PRRSV RNA in sera was measured by RT-qPCR. At 7 dpc, the virus loads in peripheral blood from the group of MLV plus rpVAX1-SwIL-4 were significantly lower than those of MLV alone, MLV plus pVAX1, and the control group of PBS (p<0.05). Meanwhile, there was also a significant difference in the groups of both MLV alone and MLV plus pVAX1 as compared with the PBS control group (p<0.05). At 14 dpc, the virus loads in the three inoculated groups declined markedly and significantly lower than that of the control group (p<0.05). The pigs inoculated with PBS had the most highest viremia at 7, 14, and 21 dpc (Fig. 7).
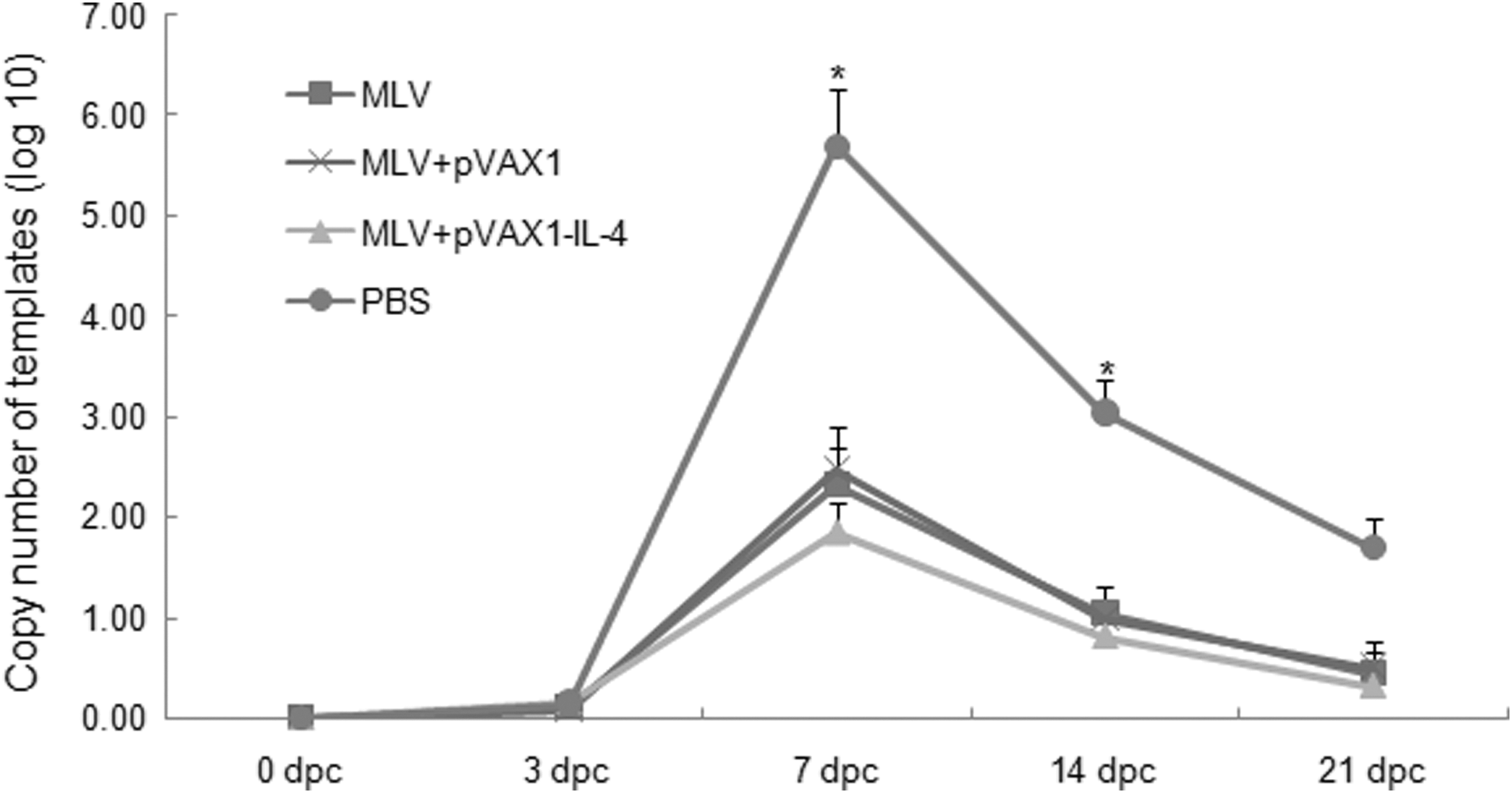
Analysis of virus loads in sera of pigs. Viral loads (viral RNA copies) in peripheral blood of pigs from every group (n=6) collected at 0, 3, 7, 14, and 21 dpc were measured by real time PCR. Asterisks indicate a significant difference in the viral titer between the group of MLV+pVAX1-IL-4 and the groups of MLV, MLV+pVAX1, or between the group of PBS and the groups of MLV+pVAX1-IL-4, MLV, MLV+pVAX1 (P<0.05) at 7 dpc. There was significant difference between the group of PBS and the others, while no significant difference among the groups of MLV+pVAX1-IL-4, MLV and MLV+pVAX1 at 14 dpc. Data were shown as mean±S.D. The standard curve here is: CT=− 3.468 log10TCID50+37.21.
Rectal temperature, clinical signs, and lung lesions after viral challenge
After viral challenge with HP-PRRSV, all pigs in the PBS control group had high fever (≥40.5°C) and displayed a range of clinical signs including inappetence, lethargy, skin cyanopathy, dyspnea, and coughing. However, pigs inoculated with MLV alone and MLV plus pVAX1 only had low fever and some mild clinical signs including inappetence, lethargy, and rough hair coats. Meanwhile, pigs in the group of MLV plus rpVAX1-SwIL-4 had no fever and just displayed a little fluctuation of rectal temperatures during 21 days post-challenge (Fig. 8). The scores of clinical signs of pigs in the group of MLV plus rpVAX1-SwIL-4 were significantly lower than those of MLV alone and MLV plus pVAX1 (p<0.05), while there was no significant difference between the latter two groups (p>0.05). More importantly, the scores of clinical signs in the three immunized groups were significantly lower than that of PBS control group (p<0.05) (Table 1).

Mean rectal temperature of six pigs in each group. Some pigs inoculated with MLV plus rpVAX1-SwIL-4, MLV alone, or MLV plus pVAX1, respectively, and the others were the control of PBS (n=6). All pigs were challenged with highly pathogenic PRRS viruses named SD-JN strain. Asterisks indicate a significant difference between the group of MLV+pVAX1-IL-4 and the groups of MLV, MLV+pVAX1, or PBS (p<0.05) at different time points. Data were shown as mean±S.D.
Within each column, values followed by different letters (A, B and C) are significantly different (p<0.05); 2Scores for clinical signs were determined by sum of daily observations of behavior, respiration, and cough according to the severity or the illness; 3Evaluation of the percentage of the entire lung affected by pneumonia.
At 21 dpc, all pigs were euthanized, and the scores of lung lesions were evaluated. The results showed that all pigs in the PBS control group had diffuse tan consolidation of the lungs. Pigs in the other three groups had no obvious lung lesions, and the scores of lung lesions were significantly lower than that of the PBS control group (P<0.05) (Table 1).
Discussion
PRRS has caused a global problem for the swine industry. In particular, HP-PRRSV has been reported as the main causative agent of large-scale outbreaks of PRRS with a high mortality in 2006 in China (33,34). Although MLV and IV against PRRSV have been available for more than a decade, the disease remains difficult to control (13). The insufficient efficacy of the vaccines is the major barrier to control this disease.
In this study, we paid much attention to the effect of swine IL-4 to induce virus-specific immune responses and the protective efficacy of pigs elicited by MLV. In a previous study, IL-4 was used as an adjuvant to a DNA vaccine targeting the ORF7 gene of PRRSV; however, it was shown to have a negative effect on the immune protection (32). The N protein encoded by the ORF7 gene could not induce neutralizing antibody against PRRSV. In our study, swine IL-4 was used as an adjuvant that had been expected to enhance the immune response of pigs inoculated with a MLV. iELISA and SN assays were performed to evaluate the antibody titer specific to the virus membrane proteins and the neutralizing antibodies, respectively. The neutralizing epitopes were considered to be located in the membrane proteins of PRRSV, but not in the N protein. Therefore, the quantity of antibodies against the membrane proteins can indirectly indicate the level of neutralizing antibody in pig sera. The level of neutralizing antibodies were measured by using an iELISA kit, which was coated with the membrane proteins including GP2, GP3, GP4, and GP5 (9). The results from the two assays showed that the titers of both antibody against membrane proteins and neutralizing antibodies were significantly elevated following the addition of IL-4, especially in the post-challenge period. Both the membrane protein antibodies and the neutralizing antibodies of pigs in all groups transiently declined due to the challenge of a large quantity of HP-PRRSV; however, the titers from the groups of MLV alone and MLV plus pVAX1 did not rise again in the experiment. Obviously, swine IL-4 play an important role in enhancing the humoral immune response of pigs inoculated with MLV.
Furthermore, we monitored the percentage of several T lymphocyte subpopulations including CD3+, CD3+CD4+, and CD3+CD8+ surface markers. Our data showed that the frequency of total CD3+ T subpopulations of pigs from all groups was relatively stable during the inoculation period. The frequency of total CD3+ T subpopulations of pigs from the group of MLV plus pVAX1-SwIL-4 climbed after viral challenge. On the contrary, the frequency of the CD3+ T subpopulations from pigs inoculated with MLV alone, MLV plus pVAX1 and PBS showed almost no change except for the time point of 7 dpc in PBS control group. The ratio of CD3+CD4+/CD3+CD8+ T subpopulations of pigs from the group of MLV plus pVAX1-SwIL-4 began rising from 42 dpi and climbed to a constant high point after viral challenge. However, the ratio from the other groups had no significant variation and even decreased in the post-challenge period as a whole. These results demonstrated that the participation of IL-4 improved the frequency of CD3+CD4+ T subpopulations and reduced that of CD3+CD8+ T subpopulations in peripheral blood of pigs, meanwhile the ratio of CD3+CD4+/CD3+CD8+ T subpopulations was improved markedly, especially after viral challenge. It is well known that the CD4+ and CD8+ T lymphocytes play a critical role in immune responses to antigens. The increased ratio of CD3+CD4+/CD3+CD8+ T subpopulations indicated that IL-4 could initiate some process to enhance the T helper activity in the pigs. Therefore, we speculate that the addition of IL-4 is closely associated with the enhancement of antibodies against PRRSV. To our knowledge, this is the first time to demonstrate that the variation tendency of CD3+, CD4+, and CD8+ T subpopulations in the peripheral blood of pigs inoculated with MLV plus recombinant plasmid encoding IL-4.
To distinguish the HP-PRRSV strain (SD-JN) with the vaccine strain in the peripheral blood, we designed a pair of specific primers that were located in the HP-PRRSV (SD-JN) Nsp2 region, and the targeting gene fragment was absent in the MLV strain TJM-F92. The TJM strain virus contains 360 nucleotides (120 amino acids) deletion. Therefore, the virus loads detected in sera are only directed to the HP-PRRSV strain (SD-JN), but not to the vaccine strain TJM-F92. The viremia of pigs inoculated with MLV plus pVAX1-SwIL-4 was significantly lower than those of MLV alone, MLV plus pVAX1 and PBS, and all of the former three groups had not almost been detected at 14 dpc.
Based on the enhanced neutralizing antibody titer and CMI response, we selected a more higher challenge dose of 2.0×105.0 TCID50 of HP-PRRSV strain SD-JN (19), which was significantly higher than that of the MLV recommendation. HP-PRRSV strain SD-JN shares a high nucleotide identity (98.84% nucleotide identity in the ORF5 gene) with the TJM strain, both of which are homologous. It should be emphasized that, although challenging the pigs with a relatively high dose of HP-PRRSV, all pigs inoculated with vaccines did not show obvious lung lesions and serious disease. Moreover, the pigs inoculated with MLV plus pVAX1-SwIL-4 displayed more mild clinical signs and lower virus loads as compared with those of MLV alone and MLV plus pVAX1. Thus, we think that although the MLV is efficacious for pigs to prevent PRRS, the pigs inoculated MLV plus pVAX1-SwIL-4 obtained more sufficient protection than those of MLV alone and MLV plus pVAX1.
In conclusion, swine IL-4 can markedly enhance the immune responses of pigs and elevate the protective efficacy to pigs elicited by the MLV. We think that it might be an attractive candidate adjuvant to MLV for the prevention and control of highly pathogenic PRRSV infections in pigs.
Footnotes
Acknowledgments
The authors of this article wish to acknowledge Dr. Yongning Zhang at Institute of Animal Quarantine, Chinese Academy of Inspection and Quarantine, for his assistance in the development and data analysis of tests. Also we would like to acknowledge Ms. Lingzhi Yang and Mr. Ping Yang for their help with the animal care and husbandry.
This work was supported by National Natural Science Foundations of China (Grant No. 31170146 and 31100119). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author Disclosure Statement
The authors declare that they have no competing financial interests.