
Abstract
Background: After invasion of hepatocytes and immune cells, hepatitis C virus has the ability to escape from the host immune system, leading to the progression of disease into chronic infection with associated liver morbidities. Adenosine 5’triphosphate (ATP) is released in most of the pathological events from the affected cells and acts as a signaling molecule by binding to P2X receptors expressed on the host's immune cells and activates the immune system for pro-inflammatory response. Therefore, the present study was designed to analyze the transcript expression of the ionotropic purinergic P2X receptors on peripheral blood mononuclear cells (PBMCs) of chronic HCV patients to have study the immune responses mediated by P2X receptors in chronic HCV infections. Methods: PBMCs were isolated from the collected blood samples. Transcript analysis of P2X receptors in PBMCs was done. The identity of amplified product was confirmed by sequencing PCR, while the quantification of the transcript expression was done by real time PCR. The relative expression of the P2X receptors was analyzed by unpaired Student's t test using GraphPad Prims 5 software. Results: We found that out of seven isoforms of P2X receptors, P2X1, P2X4, P2X5, and P2X7 receptors are expressed on the PBMCs. P2X1 and P2X7 are significantly upregulated in treatment-naïve chronic HCV patients by 2.2- and 2.5-fold, respectively. However, only P2X7 expression is found increased by 2.7-fold in patients achieving sustained virological response (SVR) after antiviral treatment compared to healthy controls. The expression of P2X receptors remained unaltered in chronic HCV patients not responding to the treatment. Conclusion: The present study confirms the significant involvement of P2X receptors in the immune responses mediated by the PBMCs in the chronic HCV infection, which should be further investigated to devise strategies to augment the immune system against this chronic viral disease.
Introduction
HCV is a hepatotropic virus and mainly infects the hepatocytes; however there are many studies that have reported extra-hepatic invasions by this virus (4). Among the extra-hepatic reservoirs, a major contribution is by the peripheral blood mononuclear cells (PBMCs) in which HCV has been found replicating or resting in quiescent phase in a number of studies (46).
The genome of HCV comprises of ? 9600 nucleotides, which contains a single open reading frame (ORF) coding a polyprotein that is processed to produce structural and nonstructural HCV proteins. These viral proteins interact with various cellular functions in a complex fashion that is still not properly identified (39). Research in the last 2 decades has shown that HCV proteins perturbs the host's cellular machinery and leads to a variety of pathological events, including production of reactive oxygen and nitrogen species in infecting cells, accumulation of fats in hepatocytes, deposition of extracellular matrix in hepatic structure, suppression of the immune response, insulin resistance, programmed cell death, and oncogenesis (8,48). Many studies have revealed that adenosine 5’-triphosphate (ATP) is released from the stressed and dying cells in extracellular milieu as a result of mechanical stress (altered cell volume) and changes in physiological functions/integrity of cell membrane induced by these pathological events (13).
Extracellular ATP (eATP) and its metabolites (adenosine 5’-diphosphate ADP and adenosine) have now a well-established role as signaling molecules, besides their role in intracellular metabolic and energy processes (23).These molecules, upon release from the damaged and stressed cells, activate the purinergic receptors that are categorized into three distinct families on the basis of molecular, pharmacological, and physiological properties. P2X receptors are eATP-activated ligand gate ion channels, while P1 (adenosine receptors) and P2Y receptors are G protein-coupled metabotropic receptors. The P2X receptor family is comprised of seven P2X receptor subunits (P2X1, P2X2, P2X3, P2X4, P2X5, P2X6, and P2X7), which are about 379 (P2X6) to 595 (P2X7) amino acids long, with 30%–50% similarity between the amino acid sequences (31). These subunits vary in affinity for ATP and assemble together as homo or hetero trimers to form cationic channel in the membrane which, upon activation by eATP, lead to downstream signaling events specific for the receptors subtype, cells expressing those receptors, and the stimulus (47).
P2X receptors are widely expressed on the liver cells and the peripheral blood mononuclear cells (PBMCs) where they are involved in multiple cellular functions (11,49). It has been reported in many studies that ATP discharged from the stressed or decaying cell is sensed as a ‘danger signal’ by the immune cells through the P2X receptors and it activates the immune cells for pro-inflammatory response. High eATP levels are believed to be important for the activation of monocytes to form inflammasomes which triggers the release of pro-inflammatory cytokines such as IL-1β and IL-18 (17). Similarly, P2X receptor-mediated purinergic signaling is known to be involved in maturation and activation of dendritic cells, resulting in release of IL-1β, IL-6, IL-18, and TNFα from the dendritic cells, along with the induction of Th 1 or Th 2 mediated immune response (10). It is also well known that activation of P2X receptors on T lymphocytes through autocrine and paracrine release of ATP serves to amplify the antigenic co-stimulatory signals and boosts effector T cell response by increased production of IL-2 (12).
HCV, on the other hand, is known to escape the innate and adaptive immune response in favor of viral persistence and chronic HCV infection (38). HCV proteins suppress the innate immune response by hindering the activation of NK and dendritic cells, thus attenuating the release of IFNs and IL-12 while increasing the release of anti-inflammatory cytokine IL-10 (27,43). Similarly, HCV proteins hamper the adaptive immune response by impairing antiviral activity of CD8+ and CD4+ T lymphocytes and skewing differentiation into regulatory T cells Tregs (18,34).
In chronic HCV infection, very little is known regarding the involvement of purinergic signaling in immune response. In our previous study, we showed that HCV envelop proteins (E1 and E2), when stably expressed in the human hepatoma (Huh 7) cell line, significantly increased the transcript expression of P2X4 receptors by 6.2 fold (28). However, no other study is available till present to investigate the involvement of purinergic signaling complex in HCV pathogenesis.
Therefore, the current study was designed to investigate the transcript expression of the P2X receptors in the PBMCs of chronic HCV patients as compared to healthy individuals. Chronic HCV patients were categorized into three groups: i) treatment naïve chronic HCV patients; ii) treatment nonresponders; iii) patients achieving SVR after antiviral treatment. PBMCs are responsible for the immune functions in the host's body and the transcript analysis of P2X receptors in these cells will help to apprehend the role of P2X-mediated signaling in host immune response against chronic HCV infection.
Materials and methods
Study design
This study recruited 45 chronically infected HCV patients, including 15 treatment naïve chronic HCV patients, 15 treatment nonresponders, and15 patients who achieved sustained virological response after the treatment, along with 24 healthy individuals who served as control. The treatment offered to patients was according to the standards of care set for the HCV genotype 3 infections (16). Pegylated interferon with ribavirin was administered to these patients for 24 weeks. The study design was approved by the ethical committees of the Nuclear Medicine Oncology and Radiotherapy Institute (NORI) Islamabad, and the Atta urRahman School of Applied Biosciences (ASAB) NUST Islamabad, Pakistan.
Blood sample collection
Adult chronic HCV patients (both male and female) infected with genotype 3a, with age ≥25 years, were enrolled for the study from the NORI Islamabad. Patients were HCV positive for more than 6 months, with mostly elevated ALT levels.
The HCV RNA presence in serum was confirmed by qualitative PCR assay and was quantified by Robogene HCV Quantification Kit (Catalogue No. 0207200102) in NORI and ASAB diagnostic laboratories. The quantification of HCV RNA was carried out according to the manufacturer's protocol using Applied Biosciences 7300 Real time PCR. The samples were run parallel along with the quantification standards (synthetic internal controls). HCV RNA level in each sample was quantified by the help of CT values for the sample and standard curve resulting from the analysis of quantification standard and the assay specific calibration coefficient. The HCV RNA concentration is expressed in IU/mL, where one IU/mL is equal to two copies/mL. The genotype of the virus was determined by using genotype specific primers reported by Ohno et al (32). Patients with co-infection of HBV or HIV or other associated liver disease were not recruited in the study.
Patients infected with HCV genotype 3a who achieved sustained virological response (SVR) after getting HCV treatment were also recruited from NORI Islamabad. Sustained virological response means undetectable HCV RNA 24 weeks after completing the therapy, and these patients were HCV RNA negative at the time of sampling.
Healthy individuals were selected from National University of Science and Technology Islamabad who served as controls. Controls were screened for HCV, HBV and HIV prior to selection for the current study.
Informed consents were obtained from all the selected patients and healthy participants in written.
Isolation of peripheral blood mononuclear cells
Four mL of venous blood were collected from patients and healthy controls in EDTA-coated vacutainers and was layered over Histopaque®-1077 (Catalogue No.10771, Sigma-Aldrich) for separation of the PBMCs by density centrifugation. PBMCs isolated were washed and suspended in PBS for further utilization (Table 1).
Before treatment.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; IU/mL, International Units/mL; N, number of samples; S.D, standard deviation; SVR, sustained virological response; U/L, units per liter.
All values correspond to the tests performed at the time of sampling, the parameters in treatment-naïve patients were measured after diagnosis before the imitation of the antiviral therapy. Parameters for treatment nonresponders and patients with sustained virological response (SVR) were measured after treatment except the mean HCV viral titer for the patients achieving SVR. HCV RNA was undetectable at the time of sampling in patients with SVR, and the value indicated in the table corresponds to the viral titer before the commencement of anti-HCV therapy.
RNA extraction, quantification, and reverse transcription
Total RNA from the suspended PBMCs was extracted by using TRIzol® reagent (Catalog No.15596-026, Invitrogen) following manufacturer's protocol. The extracted cellular RNA was treated with DNase I (Catalog No.EN0521, Fermentas) to avoid the risk of contamination of genomic DNA. The quality and quantity of extracted RNA was also analyzed by using Eppendrof's Biophotometer Plus prior to synthesis of cDNA. The complementary cDNA strand was synthesized from DNase I-treated RNA by using oligo (dT) 18 primer (Fermentas) and RevertAid™ H minus reverse transcriptase (Fermentas) in the presence of RibolockRNase Inhibitor (Fermentas).
All the equipment used in the process of RNA isolation, quantification, and reverse transcription was either certified by the manufacturer to be RNase free or was made RNase free by treating with 1% DEPC and autoclaving.
Identification of P2X receptors on peripheral blood mononuclear cells
P2X receptors were identified on peripheral blood mononuclear cells by PCR amplification using specific primers for the P2X receptors that were designed by using Primer 3 Software. PCR conditions used for the amplification of the P2X receptors comprised of initial denaturation at 95°C for 10 min, with 35 cycles of amplification [95°C (40 sec), 60°C (40 sec), and 72°C (40 sec)] and final extension at 72°C for 7 min.
Results were validated by using multiple samples to ensure the consistency and high concentration of the template cDNA was used so that any faint expression of the P2X receptor can be detected. PCR amplification products were visualized on 2% Agarose gel stained with ethidium bromide. PCR amplification products were extracted by Silica Bead DNA Gel Extraction Kit (Catalog No: K0513, Fermentas) following manufacturers protocol, which were sent to Macrogen Incorporation South Korea for sequencing to confirm the P2X receptors amplification.
Quantification of the P2X receptors in PBMCs
Quantification of the P2X receptors was done by Applied Biosystems 7300 Real-Time PCR System by using Maxima® SYBR Green/ROX qPCR Master Mix (2X) (Catalog No: K0221, Fermentas). Relative quantification of expression of the P2X receptors in the chronic HCV patients was performed after normalization of the expression of the mRNA by the expression of GAPDH housekeeping gene in chronic HCV patients and healthy controls by using 2−ΔΔC T method.
Statistical analysis of the data was performed by using Graph Pad Prism version 5.02. All values are expressed as mean±SEM. The expression of P2X receptors in the chronic HCV patient samples and healthy controls was compared statistically by unpaired Student t test. Results were considered significant when p value was less than 0.05.
Results
Identification of the P2X receptor expression in PBMCs
Specific primers were used to identify the P2X receptor expressed in the peripheral blood mononuclear cells. The PCR amplification results showed the expression of four out of seven isoforms of P2X receptors in the PBMCs. P2X1, P2X4, P2X5, and P2X7 mRNA were detected in PBMCs, while the transcriptsfor P2X2, P2X3, and P2X6 were not found in PBMCs. The PCR amplification products were sequenced to confirm the transcript expression of the P2X receptors. The sequencing PCR results confirmed the presence of P2X1, P2X4, P2X5, and P2X7 transcripts in the PBMCs. Sequences obtained [GenBank: JX471560, JX471561, and JX471562] showed maximum resemblance with the reported sequence when searched for homology using BLAST tool in GeneBank (Fig. 1).
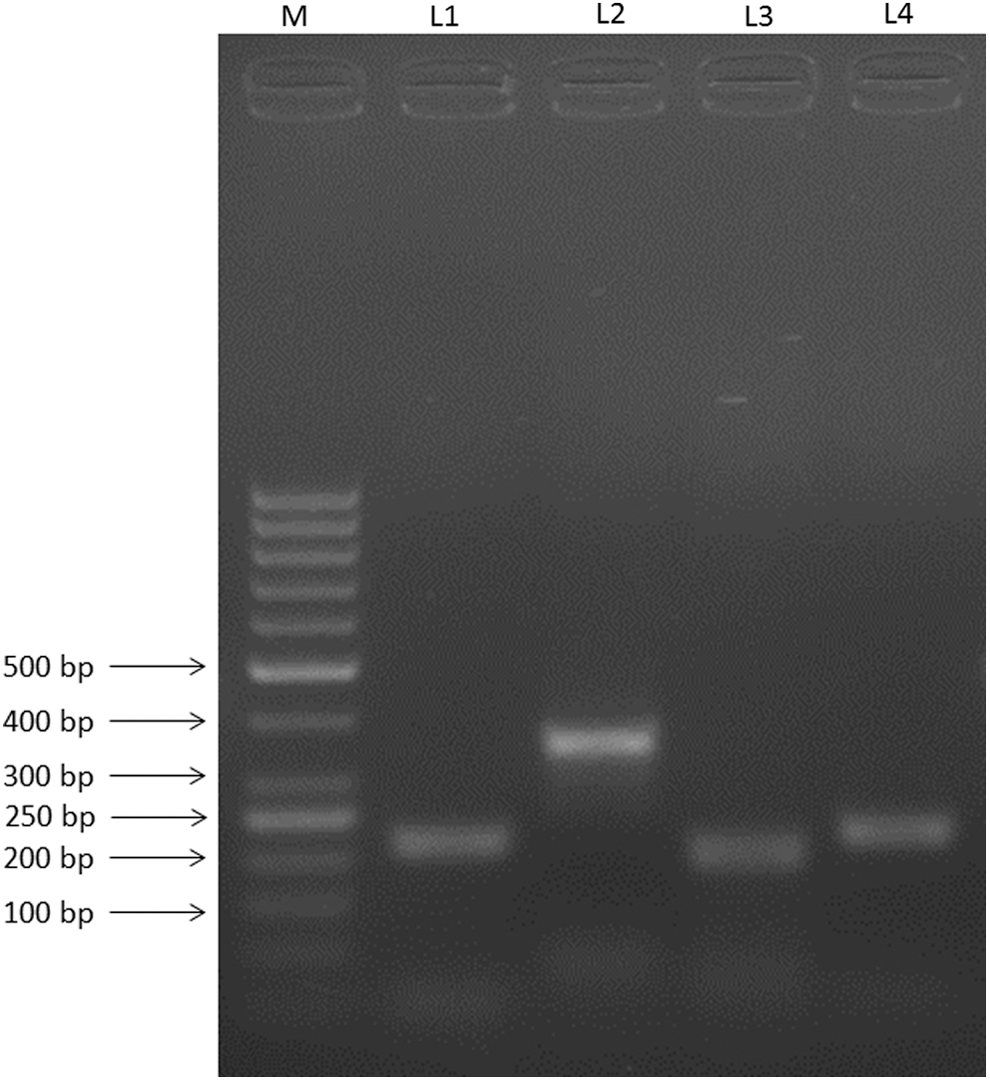
Identification of P2X transcripts in PBMCs. Lane M shows 50 bp DNA Marker. Lanes 1, 2, 3, and 4 shows the expression of P2X1 (213 bp), P2X4 (355 bp), P2X5 (193 bp), and P2X7 (220 bp) in PBMCs, respectively.
P2X receptor transcript analysis in treatment-naïve chronic HCV patients
P2X receptor transcript analysis in treatment-naïve chronic HCV infected patients and healthy controls revealed that the expression of P2X1 and P2X7 receptors isoforms was increased significantly in PBMCs of treatment naïve chronic patients.
P2X1 gene expression in peripheral blood mononuclear cells increased significantly by 2.2 fold, while the expression of P2X7 isoform was increased significantly by 2.5 fold as a result of chronic HCV infection. Expression of rest of the P2X receptors was not altered in these patients (Fig. 2).
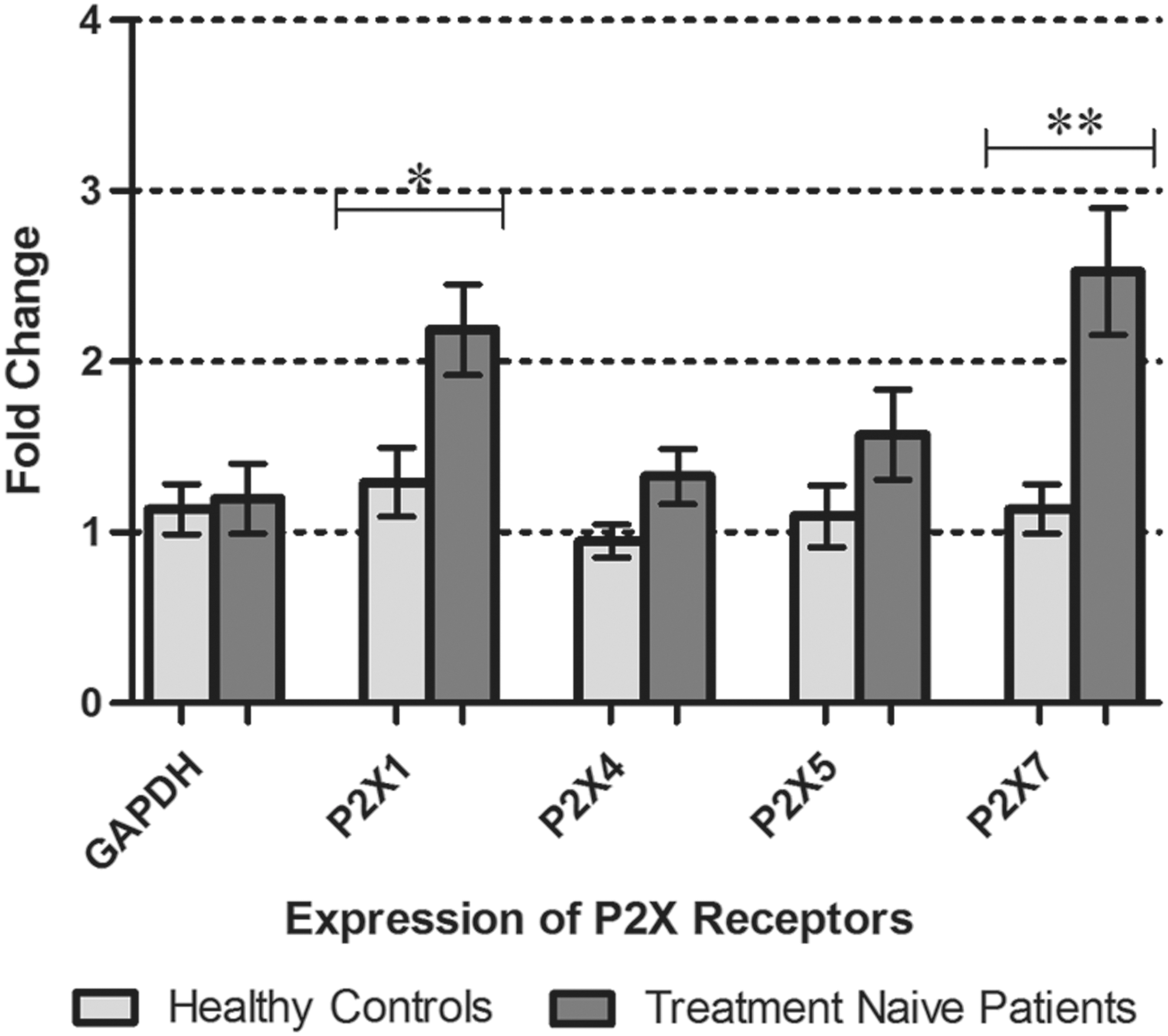
P2X receptor gene expression in PBMCs of Treatment Naïve chronic HCV patients compared to that of healthy controls. Graph showing significant increase in expression of P2X1 and P2X7 receptors by 2.2 and 2.5 fold, respectively. All values are expressed as mean±SEM; p values<0.05 are considered significant. Results represented are from fifteen individual studies with samples run in triplicate. *p<0.05; **p<0.01.
P2X receptor transcript analysis in treatment nonresponder chronic HCV patients
Patients recruited in this group had a complete course of standard therapy but they failed to respond to this treatment. The transcript analysis of P2X receptors in these patients showed that the expression of the P2X receptors in PBMCs of these patients remained unaltered despite the chronic infection and therapy (Fig. 3).
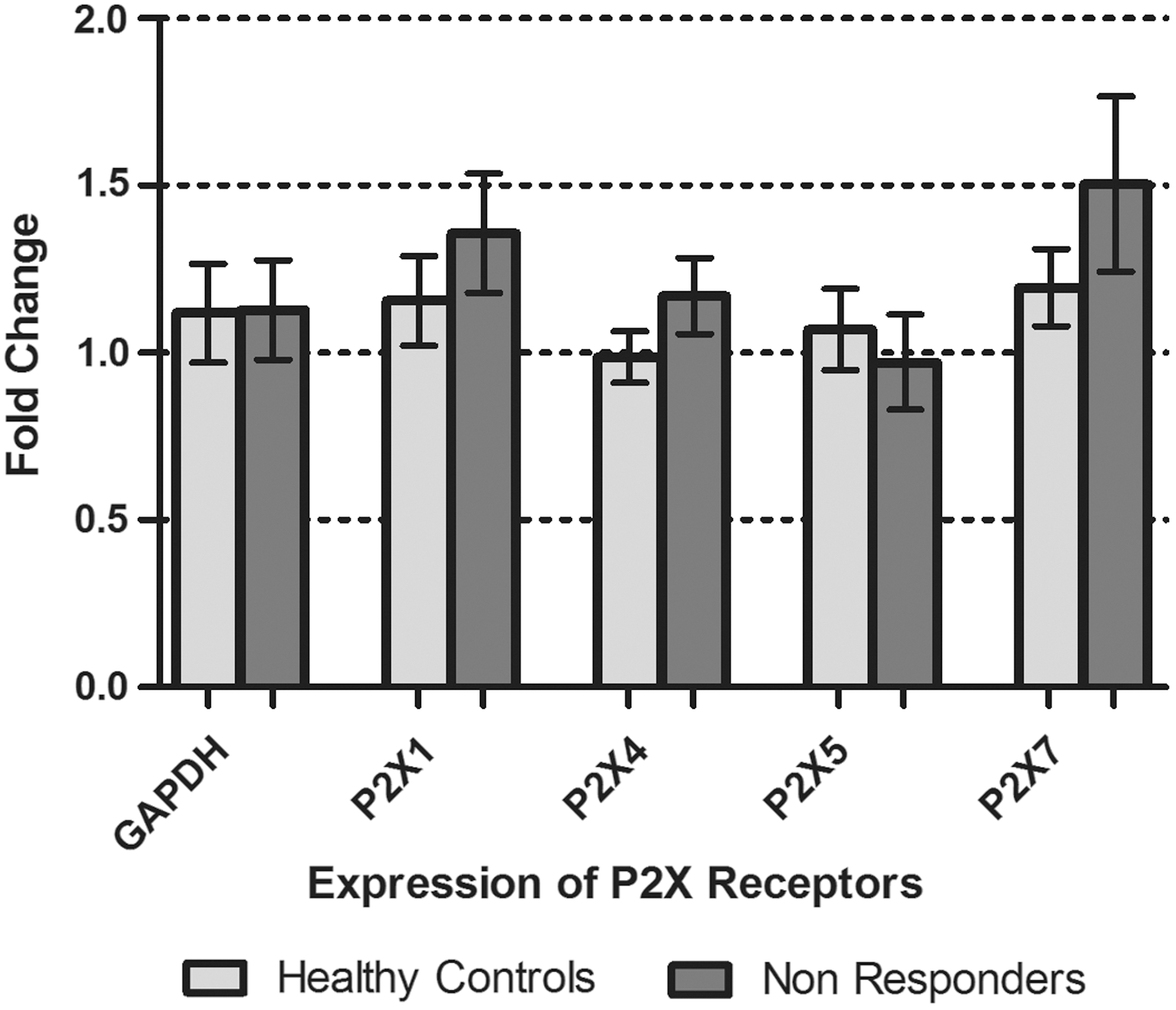
P2X receptor gene expression in PBMCs of treatment nonresponders compared to that of healthy controls. All values are expressed as mean±SEM with p values<0.05 considered as significant. Results represented are from fifteen individual studies with samples run in triplicate.
P2X receptor transcript analysis in patients with sustained virological response
Transcript analysis of P2X receptors in PBMCs of patients who achieved the sustained virological response after the treatment showed that the expression of only P2X7 receptor was significantly raised by 2.7 fold as compared to healthy individuals, while rest of the P2X receptors had almost similar expression in both the groups (Fig. 4).
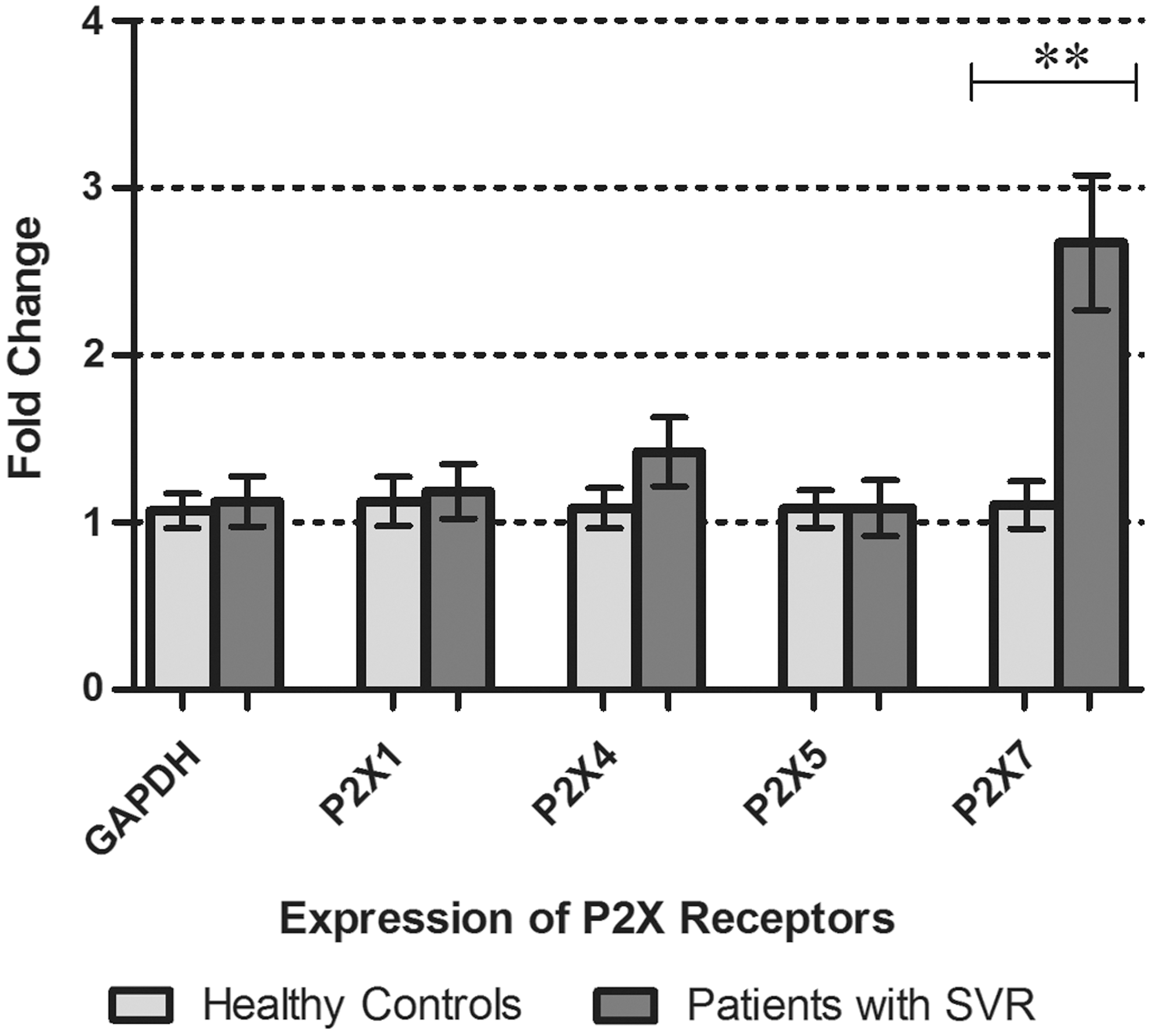
P2X receptor gene expression in PBMCs of patients with sustained virological response compared to that of healthy controls. Graph showing significant increase in expression of P2X7 receptors by 2.7 fold. All values are expressed as mean±SEM with p values<0.05 considered as significant. Results represented are from fifteen individual studies with samples run in triplicate; **p<0.01.
Discussion
HCV is well recognized for its ability to escape the immune system and persist as chronic infection, predisposing patients to steatosis, fibrosis, cirrhosis, and hepatocellular carcinoma. Though many insights have been provided by the scientific community to apprehend the molecular mechanisms involved in pathogenesis and progression of this disease, the picture is still obscure (44).
The current study was designed to compare healthy controls with the expression of the P2X receptor on the PBMCs of treatment naïve chronically HCV infected patients, patients not responding to standard antiviral therapy for HCV, and patients achieving sustained virological response (SVR). PBMCs including lymphocytes, monocytes, and macrophages are responsible for protecting the body from pathological insults and microbial invasions are now also known to host the HCV as well (35,40). Many lymphotropic HCV strains have been identified that have presented the B lymphocytes and T lymphocytes as potential reservoirs of HCV in the body (24).
Results of the current study confirmed the presence of four isoforms of P2X receptors (P2X1, P2X4, P2X5, and P2X7) on the PBMCs, which is consistent with the finding of many studies (5,51). P2X receptors are expressed on majority of the immune cells and are known to be involved in many cellular functions of PBMCs, including their activation, chemotaxis, release of cytokines, and amplification of the antigenic stimulatory signals (26,49).
Analysis of P2X receptor expression in PBMCs of different categories of chronic HCV patients revealed exciting results. This study showed a significant increase in the expression of the P2X7 receptors by 2.5 fold and 2.7 fold in the PBMCs of the treatment naïve chronic HCV patients and patients achieving SVR after treatment, respectively, compared to healthy controls. The expression of the P2X7 receptors in the treatment-nonresponders remained unaltered when statistically analyzed Fig. 5.
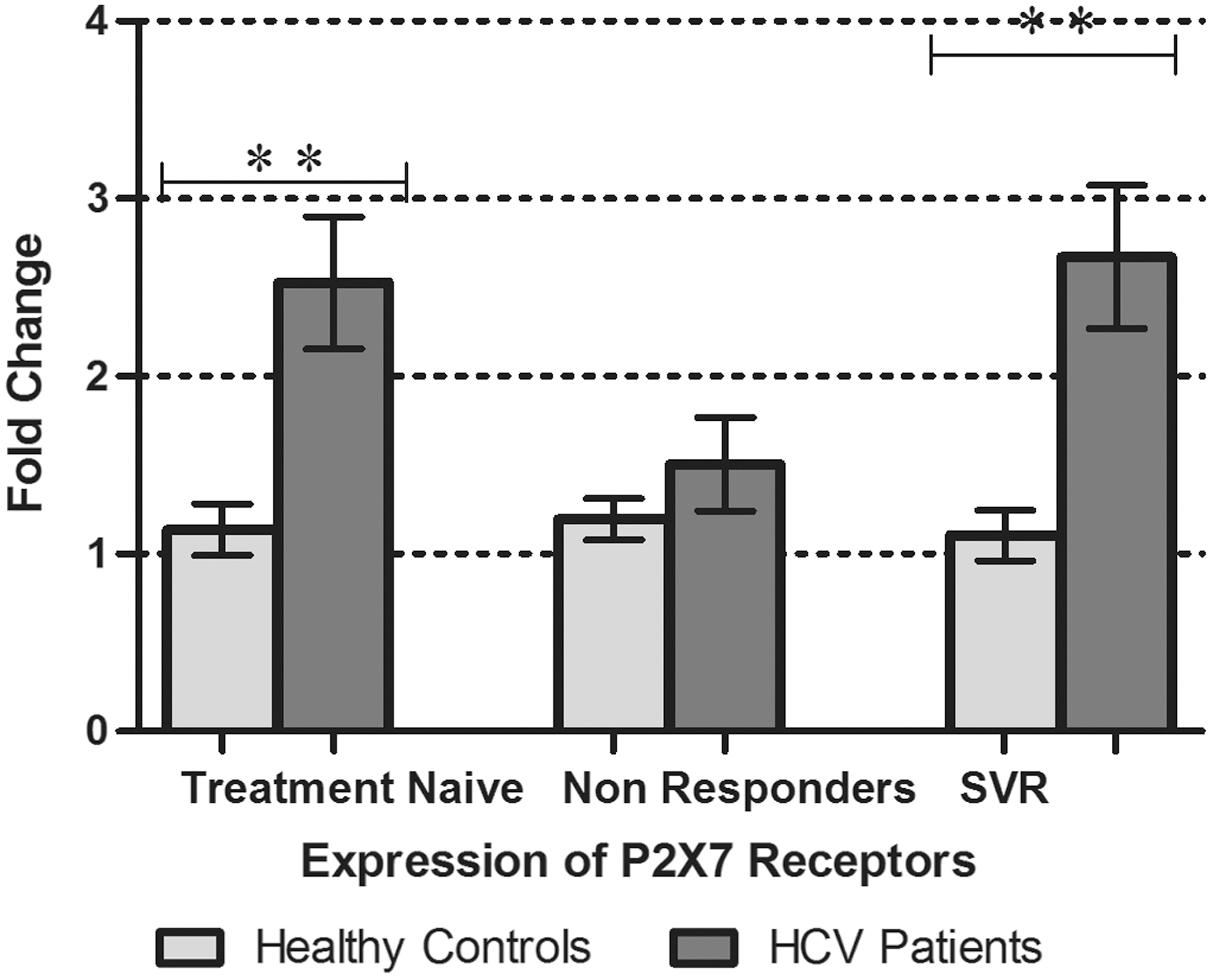
P2X7 receptor gene expression in PBMCs of treatment naïve, treatment nonresponders, and patients with sustained virological response (SVR), along with expression in healthy controls. All values are expressed as mean±SEM with p values<0.05 considered as significant.
P2X7 receptor has been represented as a key player in inflammatory responses by the immune cells in many studies (20). Its abundant expression on immune and epithelial cells, along with extra 200 amino acids at its cytoplasmic C-terminal, differentiates it with the rest of the members of P2X receptors family (37). It has lower affinity for the ATP than other P2X receptors and requires higher concentration of ATP for its activation, which suggests its role to sense eATP as a danger signal in pathological conditions (53). On brief exposure to ATP, it causes opening of cationic channels similar to other P2X receptors, but on prolonged stimulation with eATP, it leads to opening of a large pore permeable to molecules of 0.9 KDa, which induces apoptosis in the infected cells. P2X7 receptor activation has been reported to stimulate the immune system by inflammasome formation, resulting in production of pro-inflammatory cytokines IL-1β and IL-18, activation of MAP kinase pathways leading to production of reactive oxygen and nitrogen species from the immune cells, and activation of phospholipase D which augments phagosome–lysosome fusion to lyse intracellular or phagocytized pathogens (29). Activation of P2X7 receptors on T lymphocytes results in influx of Na+ and Ca+2 with efflux of K+ through the nonselective cationic channel leading to activation of various molecular pathways in the downstream signaling resulting in increased IL-2 levels and activated B and T cells production (2,33,54). It has been demonstrated in many studies that clearance of HCV in acute phase is associated with vigorous effector T cells response with concomitant higher levels of IFNγ and IL-2, while progression of infection to chronic state is associated with impaired effector T cells function with decreased capacity to produce IFN and IL-2 (19). So, increased P2X7 receptor expression and activation may be helpful to provoke the immune cells to fight and clear the virus from the body in chronic infection.
P2X7 receptor-mediated signaling is also known to tilt the balance of T cell differentiation in favor of effector T cells instead of immunosuppressive CD4+CD25+Foxp3+T regulatory cell (Tregs) (1,52). Increased Ca+2 influx as result of activation of P2X7 receptor leads to stimulation of extracellular signal-regulated kinases (ERK) dependent signaling pathway that suppresses the transcription of Forkhead box P3 (FOXP3) and induces retinoic acid receptor-related orphan receptor-γt (RORγt) expression (41). This compromises the stability and function of Tregs, which are upregulated in chronic HCV infection, favoring viral persistence by dampening the immune response of effector T cells (6). Increased expression of P2X7 receptor favors the differentiation of CD4+ T cells into TH17 subsets and thus may be responsible for aggressive immune response against the HCV and later resolution of the infection.
Expression of the P2X7 receptor varies with the cytokines present in the ambient environment. Pro-inflammatory cytokines such as IL-2, IL-4, IL-6, TNFα, and IFNγ, and bacterial products such as lipopolysaccharides (LPS) induce the expression of P2X7 receptors on peripheral blood mononuclear cells and human epithelial cells, whereas TGFβ1 prevents upregulation of P2X7 receptors in inflammation, limiting its role in immunity (14,55). So, there is a possibility that either the expression of P2X7 receptor is intrinsically increased in the patients achieving SVR like treatment naïve patients or it may be linked to the interferon that is administered as a major component of anti HCV therapy in these patients.
Besides P2X7 receptor playing a vital role in inducing the immune response, its pro-inflammatory effects can be detrimental as well and may contribute to pathological effects induced by HCV infection. The expression of P2X7 receptors on PBMCs has been observed to be upregulated in type 2 diabetes patients with moderate and deficient metabolic control as compared to patients with adequate metabolic control or healthy individual (15). Increased expression of P2X7 receptors in type 2 diabetes suggests that chronic inflammation may be contributing towards the loss of insulin function. Chronic HCV infection has already been identified for its tendency to predispose the patients to metabolic syndrome by developing the insulin resistance that is often characterized by increased levels of TNFα in various studies (45). Increased P2X7 receptors transcripts in chronic HCV infection may provide a further insight into the HCV-induced insulin resistance and loss of metabolic functions.
Transcript expression analysis of P2X receptors in PBMCs of the treatment-naïve chronic HCV patients also showed significant increase in expression of the P2X1 receptor. Limited scientific data are available regarding the roles of the P2X1 receptors in peripheral blood mononuclear cells. In T lymphocytes, P2X1 receptors are involved in autocrine purinergic signaling along with the pannexin-1 hemichannel P2X4 and P2X7 receptors to amplify antigenic stimulatory signals for activation of T lymphocytes (50). Most of the work on P2X1 is in reference to their expression on smooth muscle cells, cardiovascular system, central nervous system, and peripheral nervous system. The expression of P2X1 receptors is upregulated in atria in dilated cardiac myopathies (3). But in reference to immune cells, little is known about their function.
P2X receptor expression in PBMCs of treatment nonresponder chronic HCV patients is quite surprising. Expression of none of the isoforms of the P2X receptor is found responsive in these patients when statistically analyzed, despite having complete course of therapy. This predicts that maybe the immune system has failed to combat the disease because of the failure in efficient activation of the immune system. It is reported that increased eATP in small concentrations in extracellular milieu for a long time may lead to receptor internalization (downregulation) and receptor desensitization leading to decreased immune response by the cells (22). Increased eATP in chronic infection is also known to activate the immunosuppressive P1 (A2A) and P2Y (P2Y11) purinergic receptors that modulate and subsequently decrease the P2X receptor-mediated immune activation (50).
Conclusion
This report indicates that P2X receptors help the immune system to combat HCV infection, because the increased expression of P2X receptors, specifically P2X7 receptor, has been found in SVR and treatment-naïve patients. Their expression is unaltered in the treatment-nonresponder group. The current study illustrates the need to investigate the roles of P2X receptor-mediated signaling in PBMCs, especially in chronic inflammation and HCV persistence, to unveil the molecular pathways abrogated by these events, leading to failure of proper immune response. This knowledge can be useful for designing therapeutic interventions to treat and prevent chronic HCV infections by modifying and monitoring immune responses.
Author Disclosure Statement
The authors declare that they have no competing interests.
Footnotes
Acknowledgments
We are highly thankful to National University of Sciences and Technology (NUST), H-12, Islamabad, Pakistan, for providing the financial support and facilities to conduct this research.
Author contributions: SM conceived and designed the study. WA performed all experiments. WA searched the literature and drafted the manuscript. JA, QLA, MK, MT, MI, HA helped WA in sample collection and data analysis. SM critically reviewed and edited the manuscript. SM supervised the study, gave final approval, and helped WA in performing the work and data analysis. All authors read and approved the final version of the manuscript.