
Abstract
Invariant natural killer T cells (iNKT) perform different functions in different diseases. The cells were reported to protect myocarditis. However, the detail relationships between iNKT and Coxsackievirus B3 (CVB3)-induced myocarditis remain unclear. In order to investigate the correlation between the severity of CVB3-induced inflammation infiltration and the proportion of iNKT in the spleen and circulating blood, BALB/c mice were grouped into three groups according to the inflammation infiltration area of heart sections. The proportion of iNKT in CD3-positive cells in the spleen correlated negatively with the inflammation area (linear fit; R2=0.93) and virus capsid protein VP1 (linear fit; R2=0.84) in the myocardial tissue, while the proportion of iNKT in CD3-positive cells in the PBMC positively correlated with the inflammation area (linear fit; R2=0.91) and virus capsid protein VP1 (linear fit; R2=0.93) in the myocardial tissue. The results imply that iNKT might be used as a parameter for the diagnosis of myocarditis in clinical practice.
Introduction
M
Thus far, CVB3-induced myocarditis and the consequent malignant clinical outcomes have been considered for two reasons: virus progeny replication-caused cardiomyocytes damage and virus-triggered secondary immune or inflammatory injury to the myocardium (8). Therefore, it makes sense to seek for novel diagnostic or treatment methods focusing on both mechanisms simultaneously. Unfortunately, clinical drugs cannot now hit the targets simultaneously. Therefore, discovering an agent to reduce both CVB3 and the inflammation in myocardial tissue is a powerful and promising treatment. CD1d expression is essential for pathogenicity of CVB3-induced myocarditis and natural killer T cells (NKT) are CD1d restricted (6).
We discuss here the potential of NKT as a novel target in the diagnosis and/or treatment of CVB3-induced myocarditis. NKT are a heterogeneous group of T cells that combine properties of both natural killer cells (NK) and T cells. Invariant natural killer T cells (iNKT), as the most widely studied subset of NKT, have been reported to regulate the immune response similarly to regulatory T cells (Treg) (14). Therefore, iNKT might theoretically combine functions of both NK and Treg. NK could kill pathogens including virus (15), while Treg could alleviate the immune and inflammatory response (12,13,17), indicating a promising role of iNKT in both virus clearance and immune regulation. In addition, recent reports showed that iNKT had a beneficial effect in various pathologic conditions, such as experimental autoimmune encephalomyelitis (EAE) (18) and systemic lupus erythematosus (1).
Therefore, iNKT might be an ideal target beneficial for the diagnosis and/or treatment of CVB3-induced myocarditis. Recently, NKT were reported to protect CVB3-induced myocarditis (5,11), but no specific research has studied the relationship between iNKT and the disease condition in detail. In this article, we discuss this issue.
Materials and Methods
Animals
BALB/c mice (14–16 g) were purchased from the Joint Ventures Sipper BK Experimental Animal Co. All experiments were performed in compliance with the Animal Care Committee of Fudan University. All mice were randomly divided into two groups: the normal control group and the acute viral myocarditis (AVMC) group. In the normal control group (n=10), mice were injected intraperitoneally (i.p.) with 200 μL of Eagle's minimum essential medium (EMEM), while mice in the AVMC group (n=40) were injected i.p. with 200 μL of CVB3 diluted in EMEM on day 0 to induce acute viral myocarditis after 10 days. The mice were fed in a specific pathogen-free environment.
Virus
CVB3 (Nancy strain) was maintained by passage through HeLa cell cultures and then titered by plaque assay and stored at −80°C. The virus titer was measured by median tissue culture infective dose (TCID50=103, Nancy strain) in 200 μL of EMEM.
Histopathology and immunohistochemistry
Paraformaldehyde-embedded hearts were cut into 5 μm-thick tissue sections and subjected to hematoxylin and eosin (HE) staining to assess myocardial injury. Inflammation areas of the myocardium were counted with Image-Pro Plus 6.0. Slides were graded independently in a blinded manner by two observers, respectively. According to the inflammation area in myocardial tissue, the animals were then divided into three groups as the following: N: normal control group (Inflammation area/Total area=0) M: mild inflammation group (0<Inflammation area/Total area<0.5) S: severe inflammation group (0.5 ≤Inflammation area/Total area ≤1)
Immunohistochemical (IHC) analysis was performed following the methods of our previous study (23).
Flow cytometry analysis
Splenocytes and peripheral blood were withdrawn, and red blood cells (RBC) were lysed by a 5 min incubation in ACK lysis buffer (BD; 555899). Cells (106 cells, 100 μL) were resuspended and stained with PE-α-GalCer /CD1d dimer and APC/Cy7-CD3 (Bio Legend; 100222) for 60 min at 4°C in the dark. PE-α-GalCer /CD1d-dimer (BML-SL232-0100, BD550083,BD 557599, BD550083,BD 553485) was made using the method of Watarai (20). Samples were washed twice with flow cytometry staining buffer (R&D; FC001) and detected on BD LSRFortessa Special Order cell analyzer. The data was analyzed with Flowjo software.
Western blot analysis
Total proteins of the heart tissues were achieved as our earlier paper described (23). Samples containing 30 μg protein were separated with 10% SDS-PAGE and transferred to polyvinylidene difluoride (PVDF) membranes. Membranes were blocked for 1 h at room temperature with 5% BSA, and reacted with mouse anti-VP1 (1: 200; NCL-ENTERO; Leica Microsystems) at 4°C overnight. They were washed 10 min for three times with TBST and then incubated with peroxidase-affinipure goat anti-mouse IgG (1:5000; JacksonImmuno Research; 115-035-003) and HRP-GAPDH (1: 5000; Kangchen Biotech) for 2 h at room temperature. Then they were washed 6 min for three times with TBST. Finally, blots were detected with ECL (TIANGEN; PA112) on Bio-Rad ChemiDoc XRS+ systems.
Statistical analysis
The results were presented as mean±SD. Comparisons of the groups were made by one-way analysis of variance (ANOVA).The correlation between the factors was analyzed with linear fit. The statistical package employed was OriginPro 9.0.
Results
Myocardium pathological score of CVB3-induced myocardial injury
No mice died in the normal control group, while 19 mice died in the AVMC group. All mice alive were sacrificed and assessed at day 10 and the dead mice were excluded from the study. HE staining showed there was no inflammation infiltration in the myocardium of normal control group. But inflammation lesions were distributed widely in the myocardial tissue of the AVMC group (Fig. 1A).
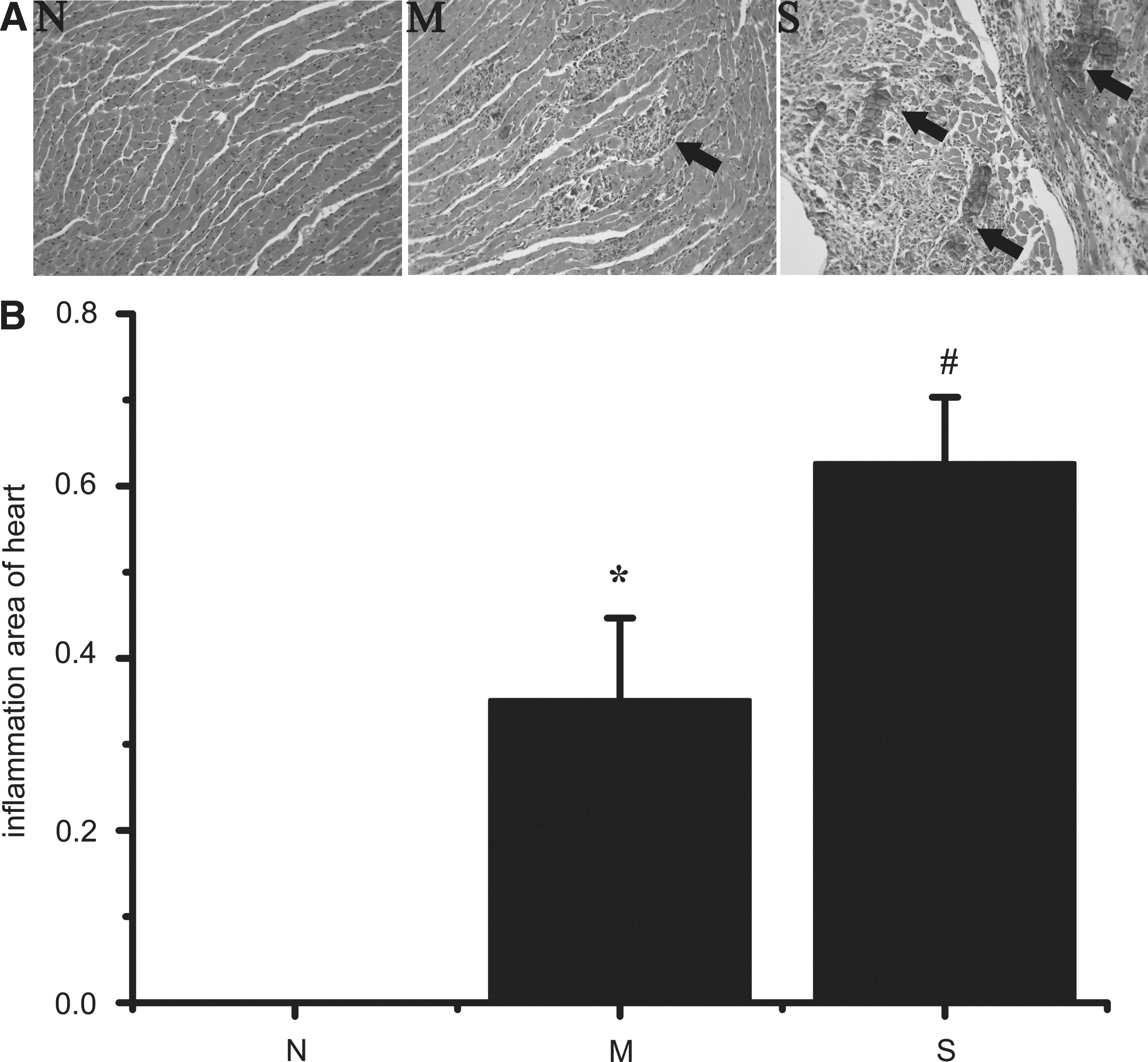
Pathological changes in myocardial tissue. All mice alive were sacrificed and assessed at days 10 after CVB3 infection; the dead mice were excluded from the study. The heart sections were stained with hematoxylin and eosin (HE). In each slice, four different microscopic fields (200X) were randomly captured, and then the ratios of inflammation area/total area were measured with Image-Pro Plus 6.0. According to the ratios, 10 mice were grouped into normal control group (n=10), 11 mice were grouped into mild inflammation group (n=11), and 10 mice were grouped into severe inflammation group (n=10).
According to the pathologic scores, 11 mice out of 21 in the AVMC group were included in the mild inflammation (M) group (0<Inflammation area/Total area<0.5), and 10 mice were in the severe inflammation (S) group (0.5 ≤Inflammation area/Total area ≤1). The difference between the groups was statistically significant (Fig. 1B; p<0.01).
Virus capsid protein VP1 expression and myocardial injury severity
CVB3 capsid protein VP1 has been used as a parameter to detect CVB3 infection in the myocardium and the severity of virus infection (9,10,22,23). Therefore, in order to evaluate the severity of CVB3 infection, VP1 was detected to measure the amount of CVB3 in the myocardial tissue with both IHC and Western blot.
Compared with the Normal group, VP1 expression increased significantly in both the Mild and Severe groups (Fig. 2; p<0.01). The Severe group also showed a significant increase compared with the Mild group (Fig. 2C; p<0.01), indicating the consistency of VP1 expression with the disease severity.
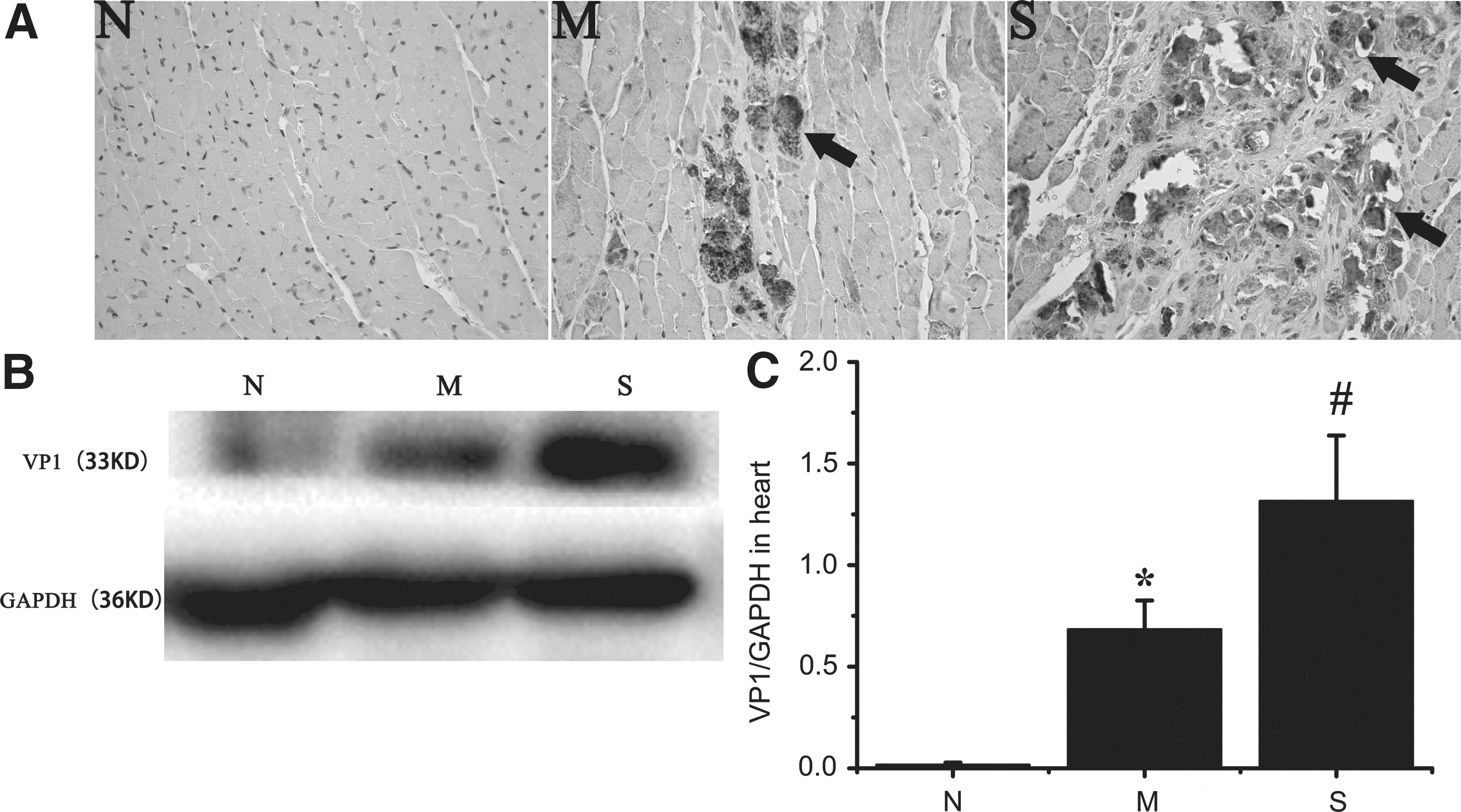
Virus capsid protein VP1 expressions in the myocardial tissue. All mice alive were sacrificed and assessed at days 10 after CVB3 infection; the dead mice were excluded from the study. VP1 was detected to measure the amount of CVB3 in the myocardial tissue with both IHC
iNKT alteration induced by CVB3 infection in the spleen
iNKT express invariant T cell receptors and are α-GalCer/CD1d dimer-dependent. Therefore, CD3 and α-GalCer/CD1d dimer were considered the specific markers of iNKT (20).
Compared with the Normal group, the frequency of iNKT out of CD3-positive cells was significantly lower in the Mild group (Fig. 3A and B; p<0.01). The proportion of iNKT in CD3-positive cells in the Severe group was also significantly lower than in the Mild group (Fig. 3A and B; p<0.01). Besides, more severe inflammation infiltration and virus capsid protein VP1 expression in the myocardial tissue were accompanied by a lower proportion of iNKT in CD3 positive cells in the spleen.
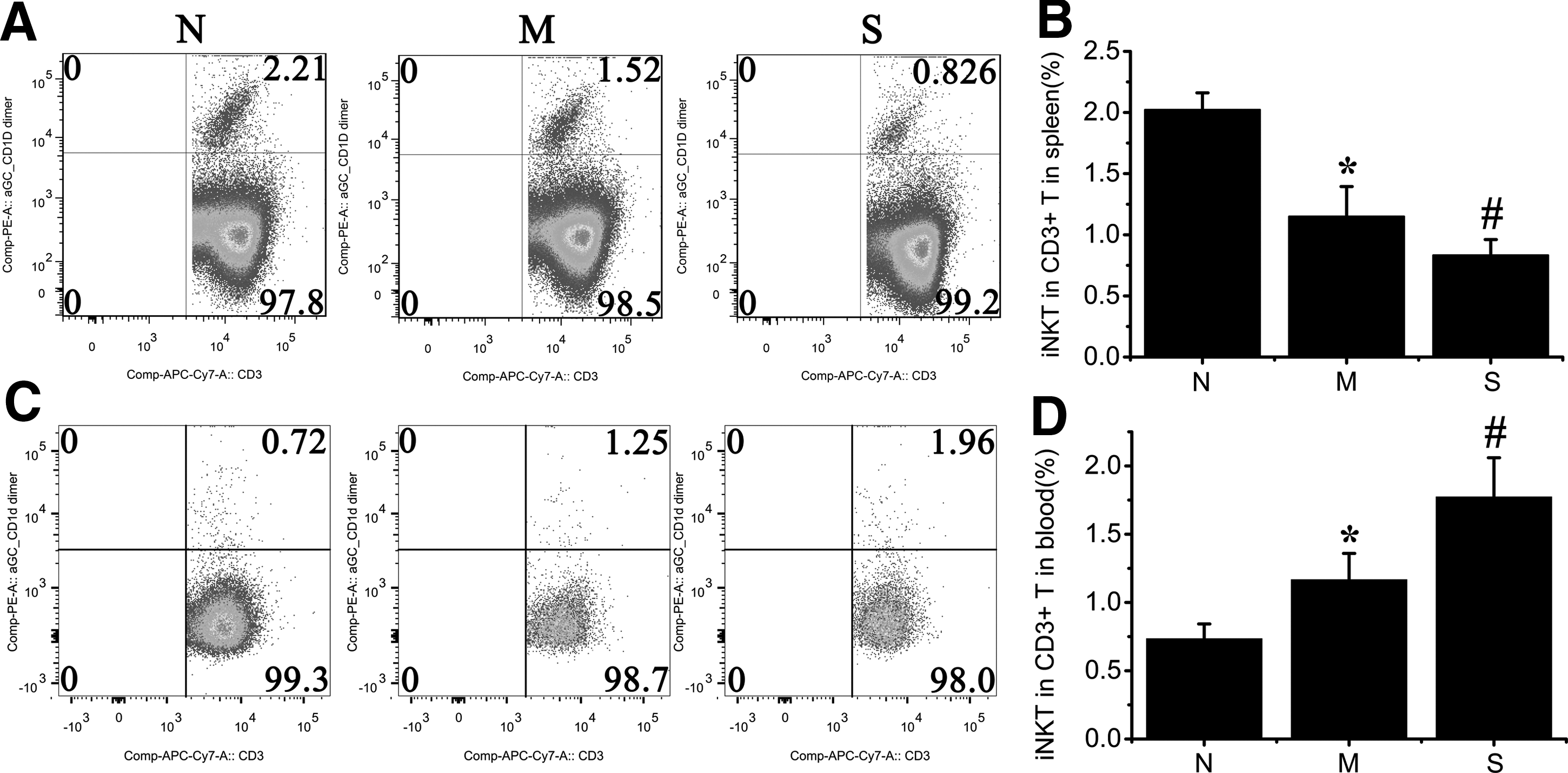
iNKT in the spleen and PBMC. All mice alive were sacrificed and assessed at days 10 after CVB3 infection; the dead mice were excluded from the study. The red blood cells in the spleen and circulating blood were lysed and then stained with PE-α-GalCer/CD1d dimer and APC/Cy7-CD3. The samples were detected with BD LSRFortessa Special Order cell analyzer. The CD3-positive cells in the lymphocytes group were gated and analyzed with α-GalCer/CD1d dimer and CD3
iNKT alteration induced by CVB3 infection in the circulating blood
Unlike in the spleen, the frequency of iNKT out of CD3-positive cells in the Mild group was significantly higher than in the Normal group (Fig. 3C and D; P<0.01). The frequency in the Severe group was also much higher than in the Mild group (Fig.3C and D; p<0.01). More severe inflammation infiltration and virus capsid protein VP1 expression in the myocardial tissue were accompanied by a higher proportion of iNKT out of CD3-positive cells in the circulating blood.
But the proportion of iNKT out of all peripheral blood mononuclear cells (PBMC) was much lower in the virus group than in the Normal group (Fig. 4; p<0.01). Unfortunately, it was not correlated with the severity of myocarditis.
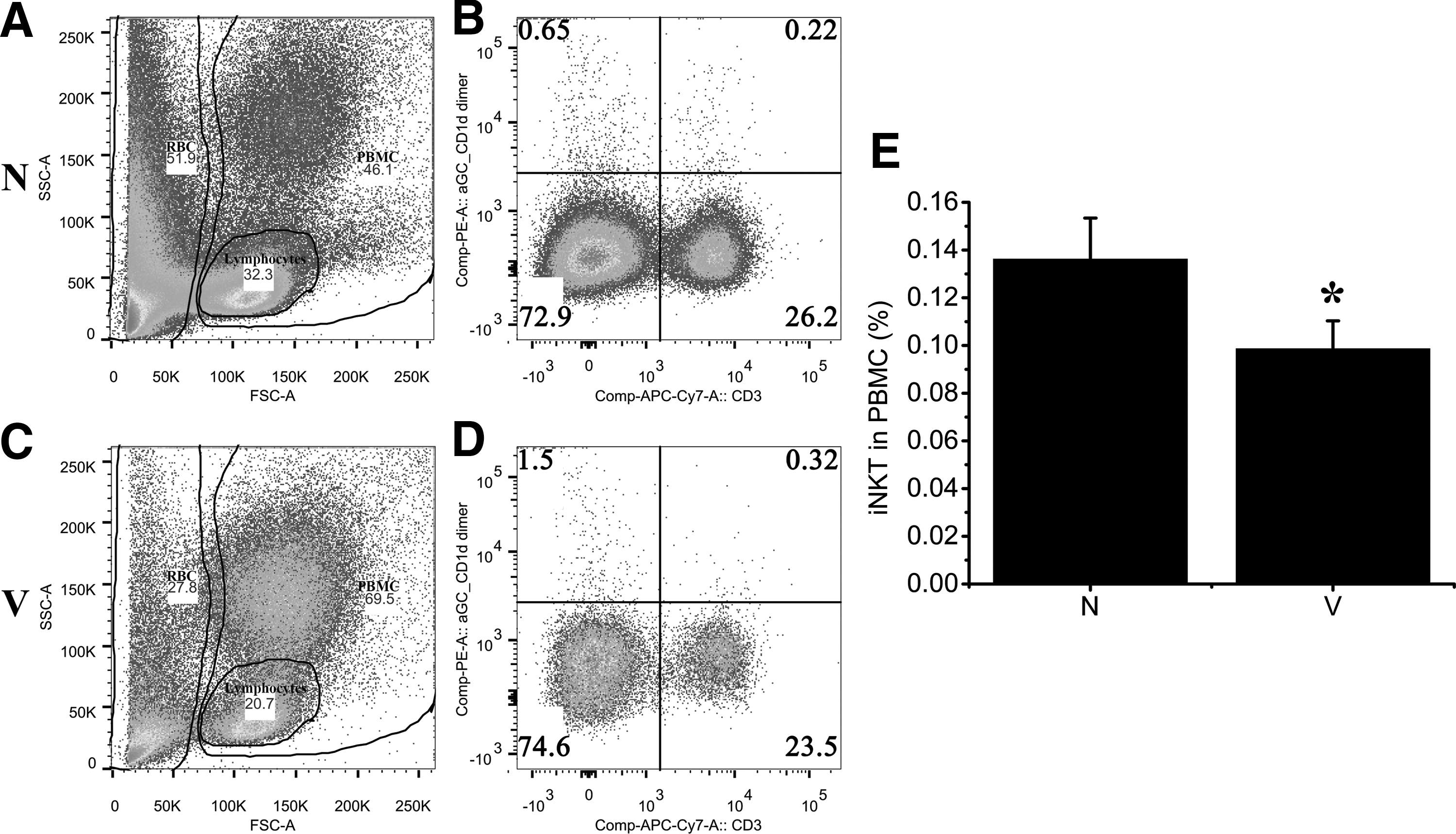
iNKT in total PBMC. All mice alive were sacrificed and assessed at days 10 after CVB3 infection; the dead mice were excluded from the study. The red blood cells in the circulating blood were lysed, and then stained with PE-α-GalCer/CD1d dimer and APC/Cy7-CD3. The samples were detected with BD LSRFortessa Special Order cell analyzer. Lymphocytes, RBC, and PBMC were respectively gated out
Correlation analysis
To determine the consistency of HE staining and VP1 detection in the evaluation of myocardial inflammation infiltration severity, the correlation between myocardial inflammation infiltration area and virus capsid protein VP1 was analyzed with linear fit (Fig. 5A; R2=0.94), indicating that both parameters could be used to evaluate the disease severity.
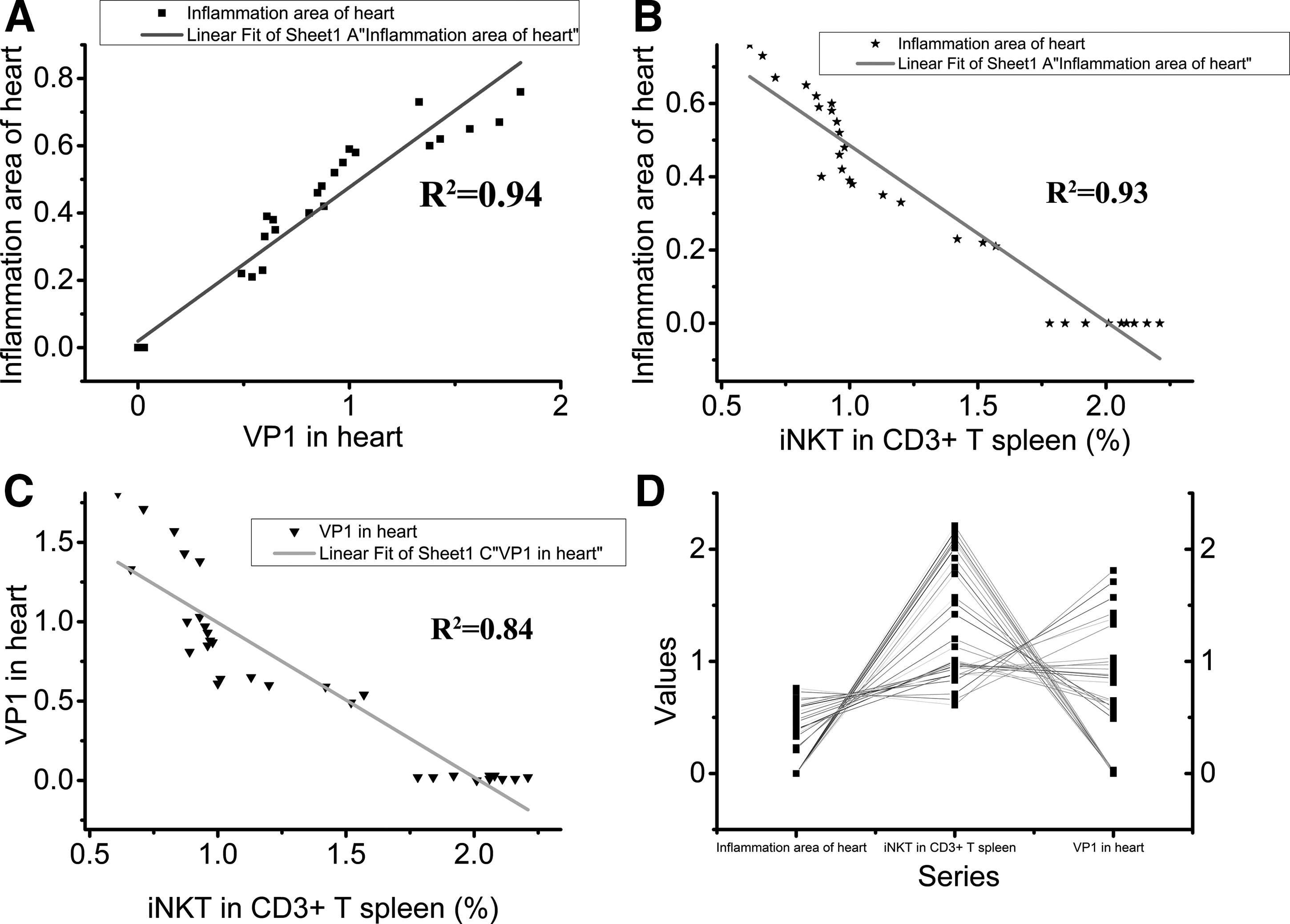
Relationships among the inflammation area, VP1, and percentage of iNKT out of CD3 positive cells in the spleen. The parameters in each individual mouse (n=31) were analyzed with OriginPro 9.0.
In order to observe the relationship between iNKT in CD3-positive cells in the spleen and in the PBMC, these two variables were analyzed using linear fit (Fig. 6A; R2=0.80).
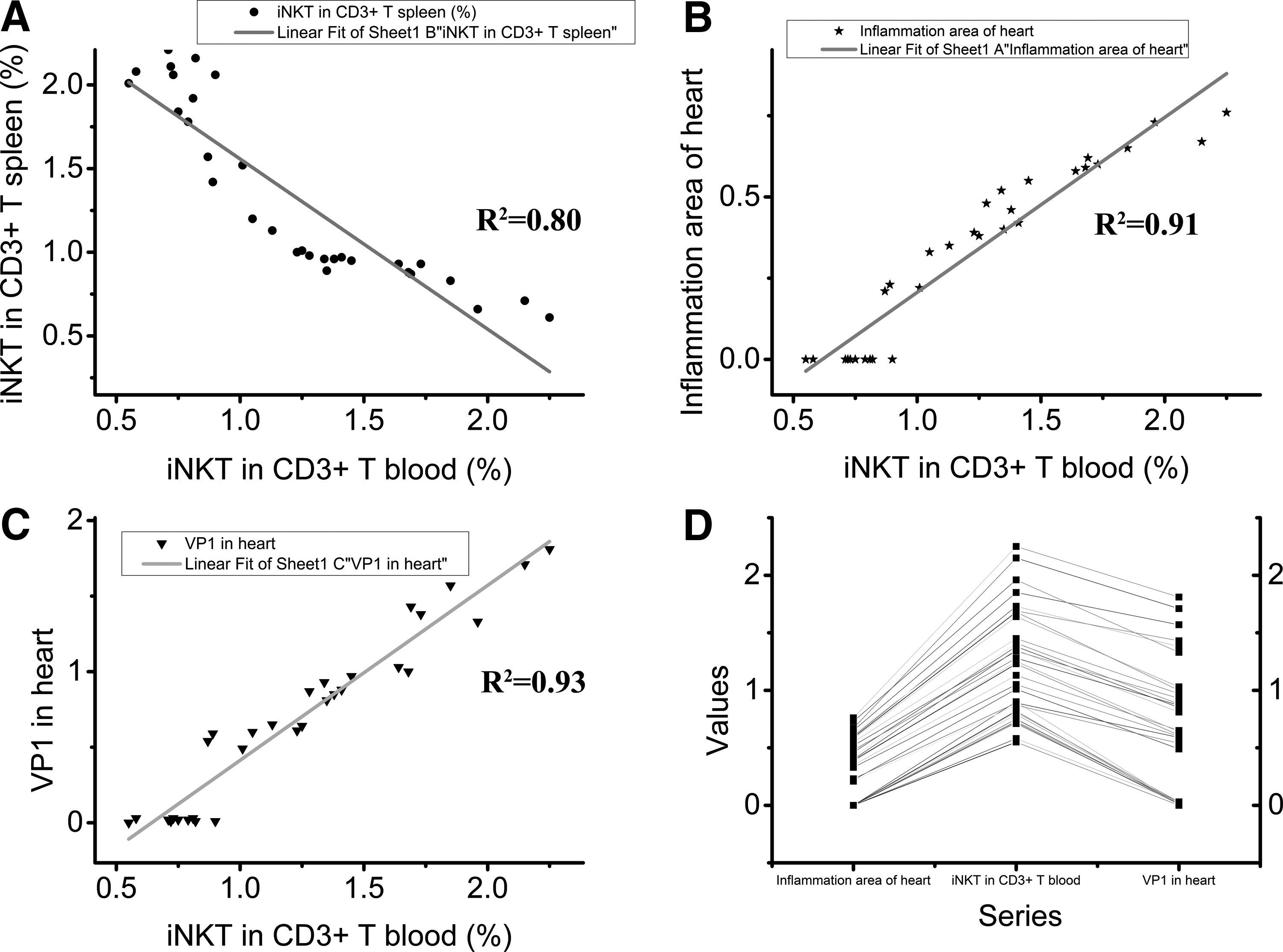
Relationships among the inflammation area, VP1 and percentage of iNKT out of CD3 positive cells in the PBMC. The parameters in each individual mouse (n=31) were analyzed with OriginPro 9.0.
To explore the correlation between myocardial inflammation infiltration and the quantity of iNKT in CD3-positive cells in the spleen/PBMC, these two variables were analyzed using linear fit (Fig. 5B, R2=0.93 and Fig. 6B, R2=0.91). Similarly, the linear fit was also performed to observe the correlation between virus capsid protein VP1 in the heart and iNKT in CD3-positive cells in the spleen/PBMC (Fig. 5C; R2=0.84 and Fig. 6C; R2=0.93). The line series of the myocardium inflammation infiltration area, iNKT in the spleen/PBMC and virus capsid protein VP1 in the heart showed that the proportion of iNKT out of CD3-positive cells in the spleen correlated negatively with inflammation infiltration area and virus capsid protein VP1 in the myocardial tissue (Fig. 5D), while the proportion of iNKT in CD3 cells in the blood showed an opposite trend (Fig. 6D). The results demonstrated that the worse the disease severity was in the myocardium, the lower proportion of iNKT out of CD3 positive cells was in the spleen (Fig. 5) and the higher it was in the PBMC (Fig. 6).
Discussion
CVB3-induced myocarditis is a common cardiovascular disease. Lack of specific symptoms sometimes led to its initial appearance in the malignant form of sudden death or persistent progression into dilated cardiomyopathy (2). Even perceived, its treatment mainly relies on supportive care, especially for the fulminant myocarditis cases (4).
iNKT performed inconsistent immune functions in different diseases. They were reported to be involved in accelerating the severity and incidence of collagen-induced arthritis (CIA) and in perpetuating the disease progression (19). But recently, they were reported to alleviate many autoimmune diseases such as experimental autoimmune encephalomyelitis (EAE) (18) and systemic lupus erythematous (13). It is still not understood why iNKT show converse functions in different autoimmune diseases. CD1d, which is essential for pathogenicity of CVB3-induced myocarditis (6), could restrict NKT, indicating that NKT might play an important role in myocarditis. Recently iNKT was reported to protect myocarditis induced by CVB3 (5,11). Interestingly, α-galactosylceramide, which was used to mobilize and activate iNKT (16,19), was reported to protect mice against the development of CVB3 induced-myocarditis (21). In addition, it is proved that α-galactosylceramide could not only stimulate NKT but also result in a block of Th17 differentiation after intranasal immunization in mice (24). Our previous study reveals that Th17 could aggravate CVB3-induced myocarditis (22). These observations imply that NKT could indeed alleviate myocarditis through block of Th17 differentiation or their immunomodulatory functions just as Treg (14).
In our study, we found that the proportion of iNKT out of CD3 cells in the spleen/PBMC altered with different inflammation levels in the myocardial tissue: the more inflammation infiltration area in the myocardial tissue, the lower proportion of iNKT in CD3-positive cells in the spleen (Fig. 5B; R2=0.93) and the higher in the PBMC (Fig. 6B; R2=0.91). This indicated that the proportion of iNKT to CD3 could reflect the inflammation severity in the heart and might provide a novel diagnostic method to assess myocarditis severity. Furthermore, the proportion of iNKT was also closely correlated with virus capsid protein VP1 expression in the myocardial tissue: more virus capsid protein VP1 in the myocardial tissue was accompanied with lower proportion of iNKT out of CD3-positive cells in the spleen (Fig. 5C; R2=0.84) and with higher proportion of iNKT out of CD3-positive cells in the PBMC (Fig. 6C; R2=0.93). This implies the potential of iNKT in the spleen/PBMC as a marker of CVB3 replication in the myocardium. In other words, if the proportion of iNKT in CD3-positive cells is low in the spleen or high in the PBMC, it might imply that CVB3 was replicating in the myocardial tissue and corresponding treatment would be needed.
iNKT was reported to be a beneficial factor in myocarditis (5,11). In our study, we found that the percentage of iNKT in CD3-positive cells in PBMC increased in the Virus group compared with the Normal control group. The results seemed to be in conflict with the previous study. Although the percentage of iNKT in CD3-positive cells in PBMC increased, the percentage of iNKT out of the total PBMC decreased in the Virus group compared with the Normal control group (Fig. 4; p<0.01).This result was consistent with the previous study that iNKT could relieve myocarditis. But unfortunately, the frequency of iNKT out of the total PBMC was not correlated with the severity of myocarditis.
In addition, iNKT might not only play an important role in myocarditis diagnosis but also might provide novel insights in the treatment of myocarditis. This requires a follow-up study in the future.
Footnotes
Acknowledgments
This project was supported by grants from the National Natural Science Foundation of China (No. 31070786; 81010007), the three-year action plan of Traditional Chinese Medicine from Municipal Health Bureau of Shanghai (No. ZYSNXD-CC-ZDYJ027) and Program of Shanghai Subject Chief Scientist (13XD1401500).
Author Disclosure Statement
No competing financial interests exist.