
Abstract
An immune role of neural stem/progenitor cells (NSPCs) has been proposed in many recent studies; however much still remains to be elucidated. In the current investigation, we report that NSPCs possess the ability to convert encephalitogenic T cells into CD4+ -CD25+-FOXP3+ regulatory T cells (Treg). Encephalitogenic and nonencephalitogenic T cells isolated from sham and Japanese encephalitis virus (JEV) infected animals were co-cultured with mouse NSPCs. Post co-culture, significant increase in the number of Tregs was observed from encephalitogenic T cells co-cultured with NSPCs. This increased conversion was found to be dependent on direct contact between T cells and NSPCs. Neutralization of TGF-β and IFN-γ in NSPC cultures abrogated this increased conversion of encephalitogenic T cells into Tregs. Flow cytometric, quantitative RT-PCR, and immunoblot analysis of both T cells and NSPCs revealed surface and intracellular changes post co-culture. Co-stimulatory molecules (B7) and ICAM-1 were increased on NSPCs post co-culture; levels of TGFβ, IFNγ, and TGFβR1 were also increased in NSPCs. This study provides a basic insight into the interaction between CNS-infiltrating encephalitogenic T cells and NSPCs during viral encephalitis. Conversion of encephalitogenic T cells into CD4+ -CD25+-FOXP3+ Tregs through interaction with NSPCs indicates an attempt in regulation of excessive inflammation in the CNS.
Introduction
T
Regulatory T cells (Treg) are a subset of the CD4+ T cells that are involved in regulating excessive immune responses. Being the regulator of immune responses, it is possible that these cells may have important roles in modulating CNS inflammation in viral encephalitis. An in vitro study involving bone marrow dendritic cell culture showed that when the cells were infected with JE virus (JEV), they resulted in expansion of Tregs, post co-culture with allogenic naïve T cells (3). Tregs were also shown to be involved in controlling the development of symptomatic West Nile encephalitis in both mouse and humans (20). Using murine model of human immunodeficiency virus encephalitis (HIVE), a recent study showed excessive ingression of Tregs in the inflamed CNS when compared to non-inflamed peripheral tissues such as spleen and liver (15). In another study using HIVE, activated Tregs were shown to attenuate astrogliosis and microglial inflammation with concomitant neuroprotection, which was due to increased neurotrophic factors in brain (21). Another investigation focusing on retroviral and herpes simplex virus infection revealed Tregs to be involved in evasion of infections by abrogation of cytotoxic CD8+ T cell responses (11,39). Adoptive transfer of Tregs into coronavirus-infected mice also resulted in decreasing T cell proliferation, dendritic cell activation, and proinflammatory cytokine/chemokine production (35). A recent study by Reuter et al. also showed peripheral Tregs to be involved in modulating viral persistence by decreasing the virus-specific CD8+ effector T cells in the brain (32).
In the current investigation we hypothesize about a novel role of NSPCs in combating CNS inflammation. We report for the first time that NSPCs have the inherent capacity to induce conversion of JEV-primed T cells (henceforth referred to as encephalitogenic T cells) into Tregs, which may be crucial in combating CNS inflammation post neuroviral infections. In many cases, survivors of neuroviral diseases suffer from various degrees of cognitive deficits resulting from neuronal death or arrest of neurogenesis. A better understanding about the ways of controlling the inflammation that leads to this may hence lead us to the development of novel therapeutic measures that may prove beneficial in the future.
Materials and Methods
Ethics statement
All animal experiments were approved by the Institutional Animal and Ethics Committee of the National Brain Research Centre (approval no. NBRC/IAEC/2008/41) and (approval no. NBRC/IAEC/2011/66). The animals were handled in strict accordance with good animal practice as defined by the Committee for the Purpose of Control and Supervision of Experiments on Animals, Ministry of Environment and Forestry, Government of India. Care was taken to minimize pain or discomfort to the animals that were used for the experiments.
Virus generation
JEV was propagated in suckling mice brain and its infectivity was determined by plaque assay, as described earlier (27).
Animal model and treatment schedule
Four- to six-week-old BALB/c mice of either sex were randomly distributed into three groups: Sham, JEV-infected and Ovalbumin (OVA)-treated. Only animals belonging to JEV-infected groups were infected with 3×105 plaque forming units (PFU) of JEV (GP78 strain). Animals of Sham group received an equal volume of filtered MEM, whereas animals of ovalbumin-treated group received 30 μg ovalbumin intravenously (30). Animals of all groups were sacrificed following the first appearance of symptoms (4 dpi) in JEV-infected group. Spleens from animals of all the treatment groups were dissected out for isolation of T cells (Supplementary Fig. S1; Supplementary material is available online at
Minocycline administration in animal model of JE
Mice were randomly assigned to three groups: Mock-infected, JEV-infected, and JEV and minocycline-treated group (JEV+M). In animals belonging to the JEV+M group, minocycline 45 mg/kg body weight was administered intraperitoneally twice daily post JEV-infection according to the previously published report (25). Virus infection was given to JEV-infected and JEV+M groups as described above.
Generation of neural stem/progenitor cells
BALB/c mouse pups (postnatal day 7) were decapitated and SVZ was dissected out aseptically in PGM buffer (phosphate buffer with 1 mM MgCl2 and 0.6% glucose). The tissue was then dissociated in a solution of 2 mg/mL papain with 50 μg/mL DNaseI at 37°C for 10 min and neutralized with DMEM containing 10% FBS. Following two washes with DMEM containing 10% FBS, the cell pellet was resuspended in Dulbecco's modified Eagles media/F12 (DMEM-F12) containing B27 supplement and 50 mg/mL gentamycin (all from Gibco, CA, USA). The suspension was passed through a 40 μm screen and then centrifuged at 300 g for 6 min. The cells were plated at density of 3×104 cells/cm2 in DMEM F12 containing B27 and gentamycin, supplemented with 20 μg/mL EGF (epidermal growth factor) and 10 μg/mL FGF (fibroblast growth factor; R&D Systems, Minneapolis, Mn). Fresh media was added after every 2 days. All in vitro experiments with neurospheres were carried out after a minimum of two passages and under cell density of 1.5×106 cells/100 mm in petri dishes. Neurospheres generated were then plated on to PDL-coated 60 mm culture dishes and 6-well plates (1×106) to get a monolayer of adherent NSPCs (Supplementary Fig. S2) (6).
Isolation of T cells and co-culture with NSPCs
Splenocytes isolated from spleens dissected from animals belonging to treatment groups described above were enriched for T cells by magnetic bead separation using T cell enrichment kit according to manufacturer's instructions (BD Biosciences, San Jose, CA). Isolated T cells were then co-cultured with NSPCs adhered to 60 mm culture dish (107cells per plate) (Supplementary Fig. S1). For transwell experiments, T cells (107) from different treatment groups were added on to NSPCs grown in PDL-coated 6-well plates carrying 0.4 μm cell culture inserts (BD Biosciences). 107 T cells isolated from sham and JEV-infected splenocytes were also co-cultured with adherent NSPCs in 60 mm plates in which either TGF-β or IFN-γ or both (1 μg/mL) were neutralized using monoclonal antibodies (R&D Systems) specific to them.
Isolation of CD4+ CD25+ and FOXP3 T-regulatory (Treg) cells
Both pre and post co-culture with adherent NSPCs, isolated T cells were subjected to Treg isolation using Dynabeads® FlowComp™ Mouse CD4+CD25+ Treg cells isolation kit (Invitrogen, Carlsbad, CA) according to manufacturer's guidelines.
Immunoblot
In order to obtain total cellular extracts, T cells and NSPCs were lysed in buffer containing 1% Triton-X-100, 10 mM Tris-HCl (pH 8.0), 150 mM NaCl, 0.5% Nonidet P (NP-40), 1 mM EDTA, 0.2% EGTA, 0.2% sodium orthovanadate and protease inhibitor cocktail (Sigma, St. Louis, MO). After 15 min on ice, lysates were centrifuged at 12,000 g for 10 min at 4°C. Supernatants were collected and stored at −30°C. Protein levels were estimated by the bicinchoninic assay (BCA) method. Western blot analysis was performed to study modulation in expression of T cell and NSPC proteins post co-culture, from total cellular extract using primary antibodies against phosphoZAP70, phosphoSMAD2/3, FOXP3, and TGFβR1 (Santa Cruz Biotechnology, Santa Cruz, CA) at 1:1000 dilutions. After multiple washes with PBS–Tween, blots were incubated with appropriate peroxidase-conjugated secondary antibodies (Vector Laboratories, Burlingame, CA). The blots were processed for development using chemiluminescence reagent (Millipore, Temecula, CA). The images were captured and analyzed using Chemigenius, Bioimaging System (Syngene, Cambridge, UK). To determine equivalent loading of samples, the blots were stripped and reprobed with anti-β-tubulin (Santa Cruz Biotechnology).
Immunohistochemistry
For immunohistochemical staining, brains from sacrificed animals were excised following repeated transcardial perfusion with ice-cold saline and fixed with 4% paraformaldehyde. Twenty micron thick cryosections were made with the help of Leica CM3050S cryostat and processed for immunohistochemical staining to detect presence of CD4+ and FOXP3+ T cells in SVZ. Sections were incubated overnight at 4°C with rabbit anti-FOXP3 (Santa Cruz) and FITC-conjugated anti-CD4 (BD Biosciences) antibodies, respectively. After washes, slides were incubated with appropriate fluorochrome-conjugated secondary antibodies (Vector Laboratories Inc.) and following final washes, sections were coverslipped after mounting with 4′–6-diamidino-2-phenylindole (DAPI, Vector laboratories Inc.). The slides were observed either under a Zeiss Axioplan 2 fluorescence microscope or a Zeiss Apotome microscope (Zeiss, Gottingen, Germany).
Flow cytometry
NSPCs were plated in 60 mm culture plates and were treated as described above. After 24 h of co-culture, cells were collected by washing with chilled sterile 1X PBS. After two washes with 1X PBS, cells were first fixed with BD Cytofix solution (BD Biosciences) and permeabilized by resuspending in permeabilization buffer (BD Cytoperm plus; BD Biosciences). Cells were then washed twice in wash buffer (PBS containing 1% bovine serum albumin) and finally resuspended in wash buffer at 1×106 cells per 100 μL. They were then treated with primary antibody (B7.1 and B7.2; BD Biosciences; 1:100 dilutions) for 30 min at room temperature. Cells were then washed and pelleted by centrifugation, followed by incubation with FITC-conjugated secondary antibody for 30 min. After final wash, cells were resuspended in 400 μL FACS buffer and analyzed on a FACS Calibur. The mean fluorescence intensities of B7.1 or B7.2 were calculated using Cell Quest Pro Software (BD Biosciences).
Quantitative Real Time-PCR (qRT-PCR)
Total RNA from NSPCs and T cells from different treatment groups and from subventricular zones of Sham and JEV-infected animals were extracted using Tri reagent (Sigma, USA). Random hexamer primers were used for cDNA synthesis using Advantage RT-PCR (Clontech, Mountain View, CA). 500 ng of cDNA was used as a template for performing qRT-PCR using SYBR Green Supermix (Bio-Rad, Hercules, CA, USA) on Rotor-Gene Q (Qiagen, Alameda, CA, USA). The real time PCR results were analyzed using instrument software and normalized with those from glyceraldehyde 3-phosphate dehydrogenase (GAPDH) internal control. The real time PCR results were analyzed by ΔΔCt method and either expressed as fold-changes over mock-infected or 2−ΔCt.
Statistical analysis
Values were calculated as the mean±S.D. of n number of independent experiments (each n from different cell preparations). Differences between mock-infected/control and experimental samples for the accumulated data were evaluated using one-tailed Student's t test. Statistical significance was set at p<0.05 for all analyses.
Results
Minocycline administration results in decreased CD4+ and FOXP3+ cells in SVZ of JEV-infected animals
Immunohistochemical staining of brain sections from animals of JEV+M groups (both 4 days and 8 days) revealed no CD4+ and FOXP3+ cells in SVZ when compared to SVZ of animals belonging to only JEV-infected group. However, there was significant increase in CD4 and FOXP3 double positive cells in SVZ of JEV-infected groups (8 days) when compared to only JEV-infected group of 4 days post-infection. Mock-infected group showed no CD4+ and FOXP3+ cells in SVZ (Fig. 1 a–b).
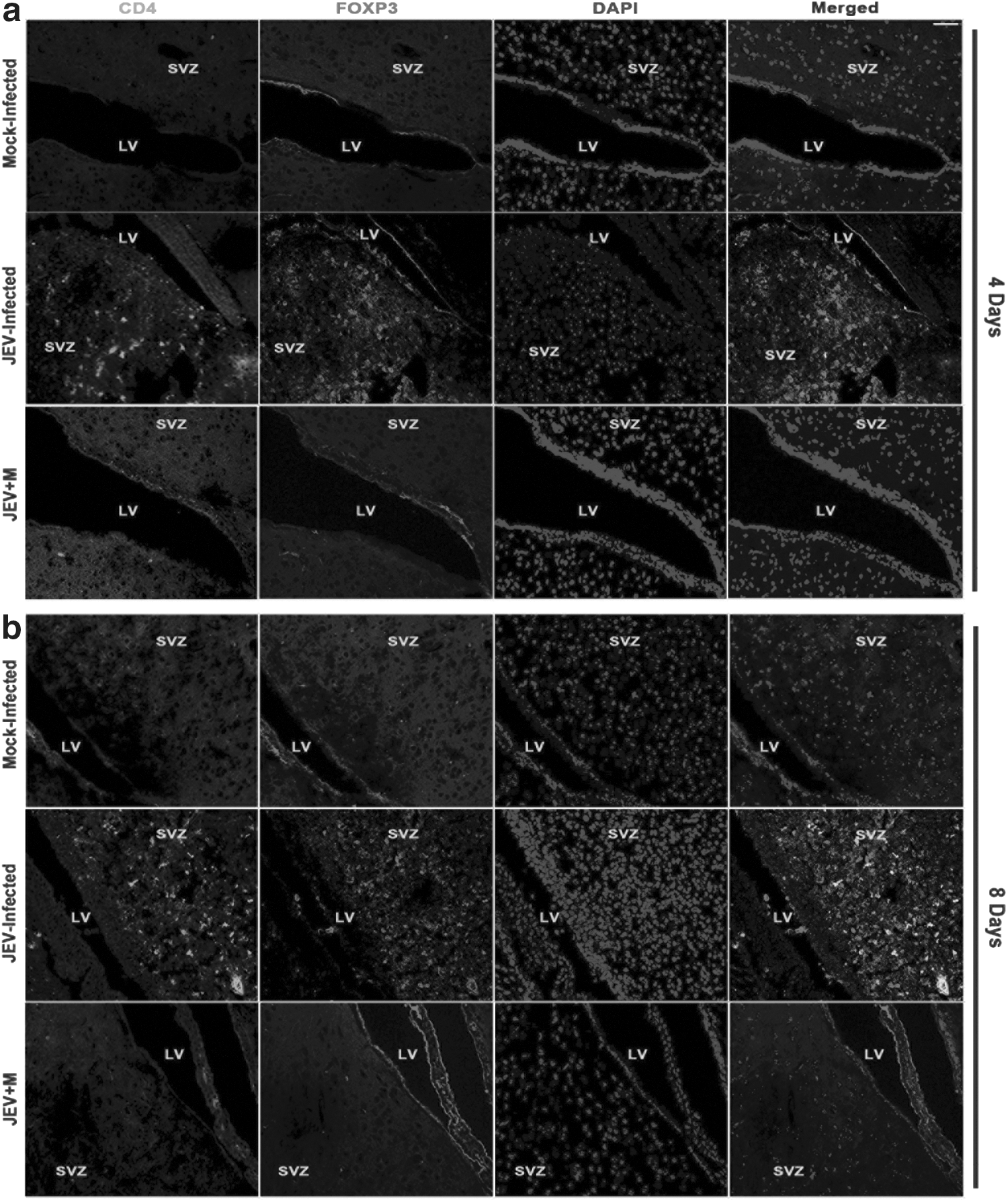
Minocycline treatment results in abrogation of CD4+ and FOXP3+ cells in SVZ after JEV infection. Immunohistochemical staining from brain sections of Mock-infected, JEV-infected, and JEV-infected minocycline-treated mice showing infiltration of CD4+ FOXP3+ cells in SVZ after 4 days
Minocycline administration downregulates B7.1, B7.2, ICAM-1, TGF-β, and IFN-γ in SVZ
Quantitative real time PCR analysis revealed that the expression of co-stimulatory molecules B7.1 and B7.2 in SVZ of JEV-infected animals showed significant increases when compared to SVZ of mock-infected and JEV+M animals both 4 and 8 days post infection (p<0.05) (Fig. 2c-d). Similarly, the expression of ICAM-1 was found to be increased (p<0.05) (Fig. 2e). Also, the levels of both TGF-β and IFN-γ were significantly increased in SVZ of animals belonging to JEV-infected groups when compared to mock-infected and JEV+M animals, both 4 and 8 days post infection (p<0.01) (Fig. 2d and e). Significant decreases in levels of B7.1, B7.2, ICAM-1, TGF-ß, and IFN-γ was observed in SVZ of minocycline administered and JEV-infected animals when compared to only JEV-infected groups both 4 and 8 days post infection (p<0.05) (Fig. 2a–e).
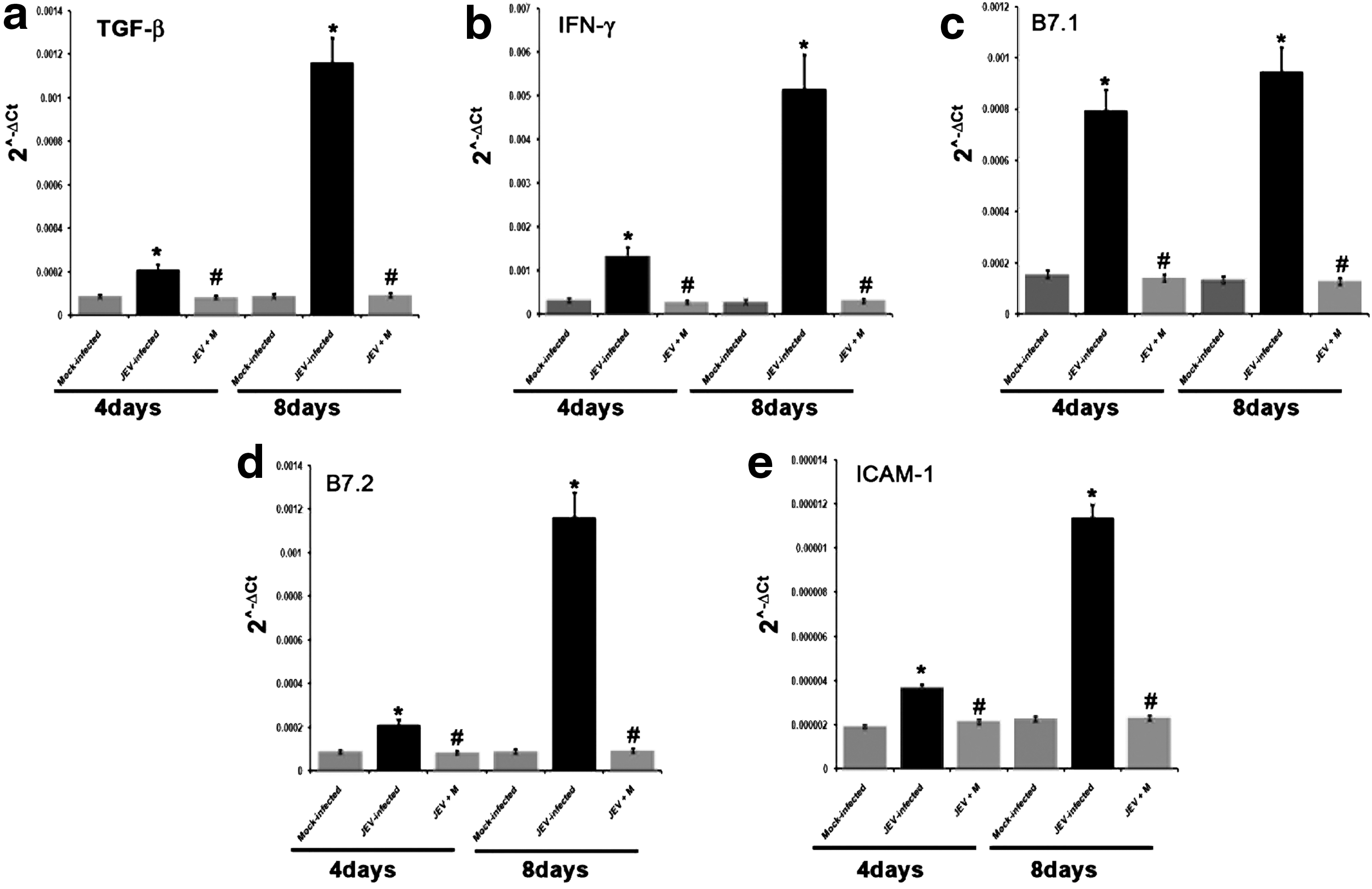
Decreased expression of co-stimulatory molecules (B7), ICAM-1, TGF ß, and IFNγ in SVZ of JEV-infected mice after minocycline treatment. Graphs representing qRT-PCR analysis of B7.1, B7.2, ICAM-1, TGF-β, and IFN-γ from RNA isolated from SVZ dissected out of Mock-infected, JEV-infected, and JEV-infected minocycline-treated mice; 4 days and 8 days represents duration of minocycline treatments post JEV-infection,
NSPCs induce conversion of encephalitogenic T cells into Tregs
T cells isolated from spleens of animals belonging to different treatment groups were subjected to Treg isolation using a commercially available kit, as described above. The purity of isolated T cells was assessed using flow cytometry following CD3 staining (>98% CD3+ purity; data not shown). Isolated Tregs were counted by the trypan blue dye exclusion technique, using a hemocytometer. Population of Treg was found to be significantly greater amongst total T cells belonging to JEV-infected groups in comparison to that of both sham and OVA-treated groups (p<0.01); no significant difference was observed in Treg numbers between sham and OVA-treatment groups (Fig. 3a). The splenic T cells of animals belonging to different treatment groups were then co-cultured with adherent NSPCs. Post 24 hours of co-culture, these T cells were then subjected to Treg isolation and counted. There was significant increase in the number of Treg when T cells of JEV-infected animals were co-cultured with NSPCs as compared to those belonging to sham or OVA-treated animals (p<0.01). However, no significant difference between Treg numbers was found when T cells of sham and OVA-treated animals were co-cultured with NSPCs (Fig 3b).
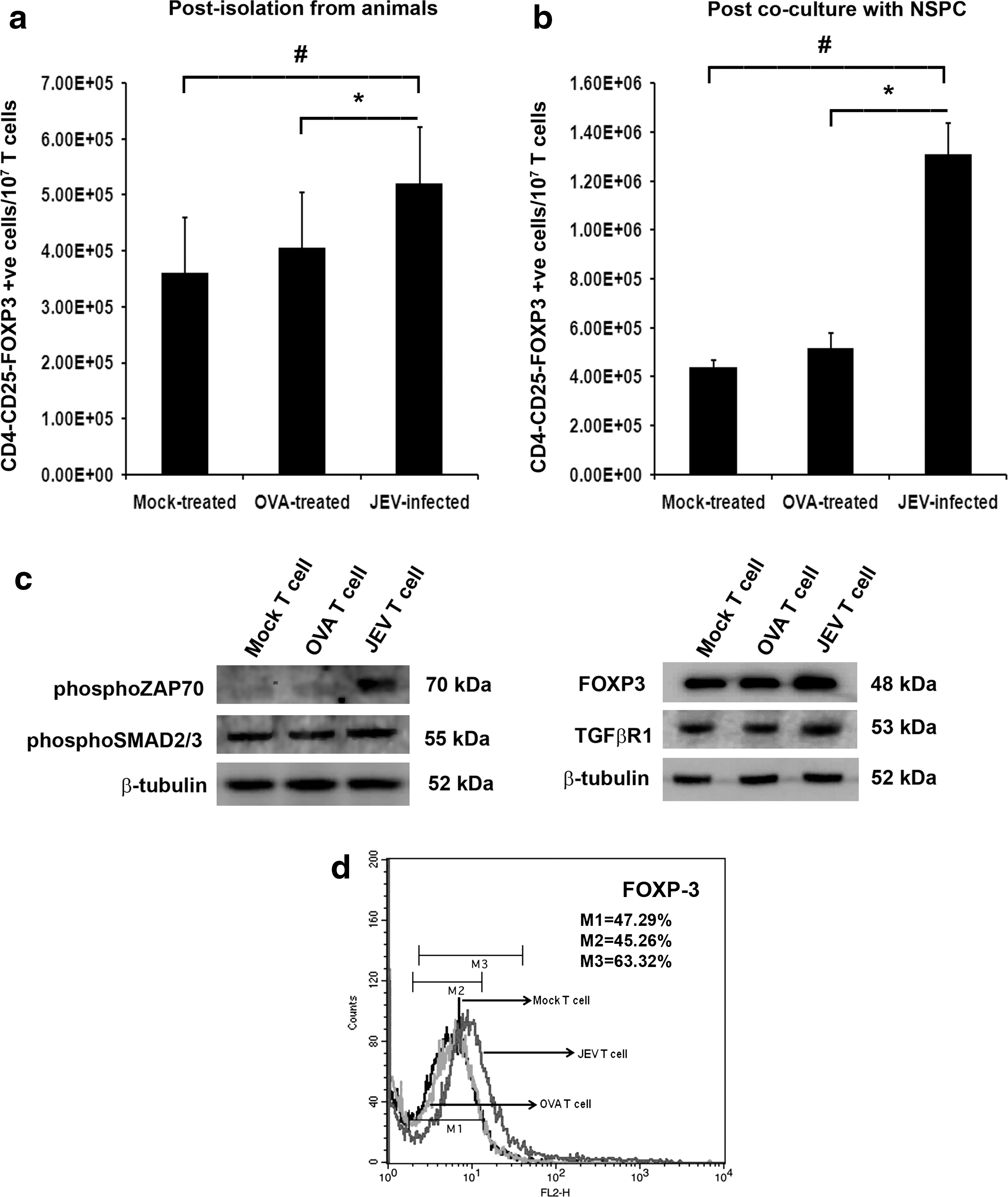
Increased CD4+CD25+and FOXP3+ Treg induction and expressions of FOXP3, TGFβR1, phosphoSMAD2/3, and phosphoZAP70 in T cells post co-culture with NSPCs. Graphical representation of number of CD4+CD25+, and FOXP3+ Treg cells from spleen of Mock-infected, OVA-treated, and JEV-infected mice per 107 isolated T cells
Increased expression of phosphoZAP70, PhosphoSMAD2/3, TGF-βR1, and FOXP3 in encephalitogenic T cells post co-culture with NSPCs
PhosphoZAP70, PhosphoSMAD2/3, TGF-βR1, and FOXP3 levels were significantly increased in T cells belonging to JEV-infected groups post co-culture with NSPCs when compared to T cells belonging to sham and OVA-treatment groups co-cultured with NSPCs (p<0.05) (Fig. 3c-d).
Encephalitogenic T cells induces upregulation of B7.1, B7.2, ICAM-1, TGF-βR1, TGF-β, and IFN-γ in NSPCs
Flow cytometric analysis of B7.1 and B7.2 revealed significant increases in their surface expression on NSPCs co-cultured with T cells isolated from JEV-infected animals when compared to NSPCs that were co-cultured with T cells belonging to sham group (Fig. 4a, b). qRT-PCR carried out to analyze the transcript levels of ICAM-1, TGF-βR1, TGF-β, and IFN-γ showed significant increases in NSPCs co-cultured with encephalitogenic T cells when compared to their levels in NSPCs co-cultured with T cells belonging to either sham or OVA-treated groups (p<0.05) (Fig. 4c-f ). Immunoblot of TGF-βR1 also showed increased level in NSPCs co-cultured with encephalitogenic T cells when compared to NSPCs co-cultured with T cells belonging to either sham or OVA-treated groups (p<0.05) (Fig. 4g).
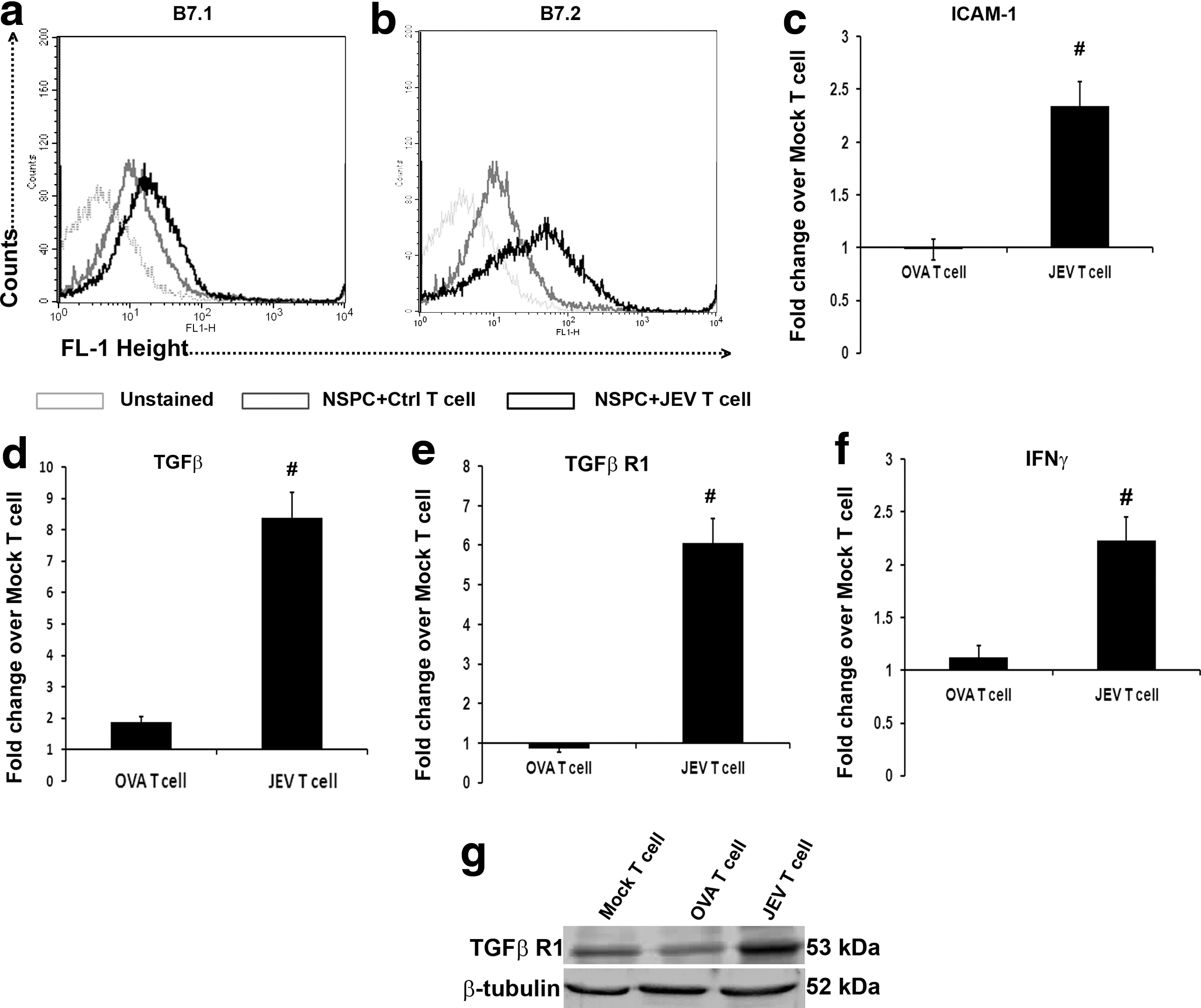
JEV-activated T cells upregulate B7.1, B7.2, TGF-β, IFN-γ, TGFβR1, and ICAM-1 on NSPCs. Flow cytometric analysis of surface staining for the expression of B7.1
Direct contact between NSPCs and encephalitogenic T cells is essential for Treg conversion
To ascertain whether direct contact between NSPCs and encephalitogenic T cells was necessary for their conversion into Tregs, we co-cultured T cells isolated from animals of different treatment groups as described above with adherent NSPCs culture, carrying 0.4 μm cell culture inserts. No significant changes in Treg numbers were observed in encephalitogenic T cells when compared to T cells from sham and OVA-treated groups co-cultured with NSPCs carrying inserts (Fig. 5a).
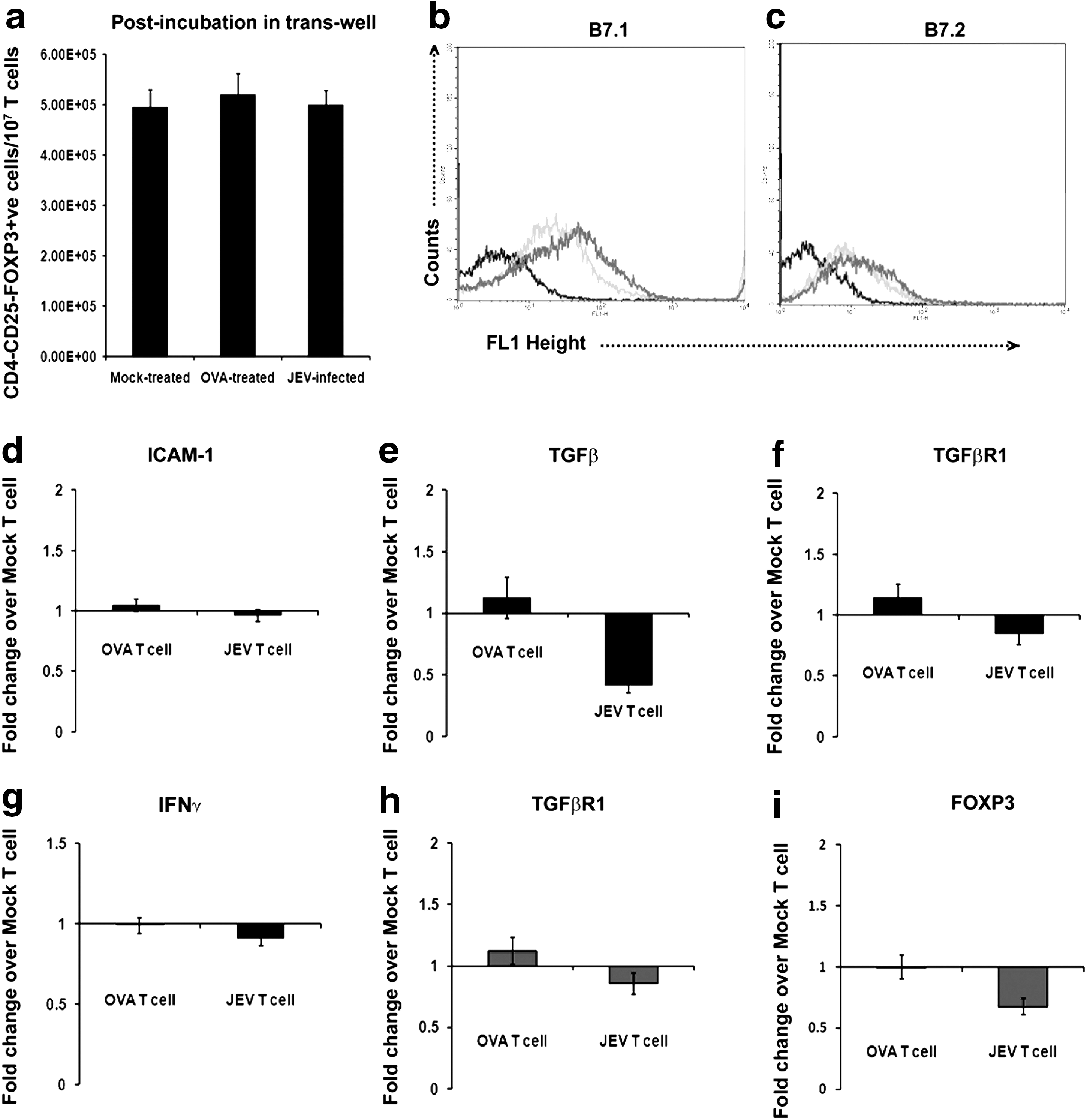
Increased CD4+CD25+and FOXP3+ Treg induction and molecular changes in NSPCs and T cells post co-culture is dependent on direct contact. Graphical representation of number of Tregs from pool of T cells of Mock-infected, OVA-treated, and JEV-infected animals, post co-culture with NSPCs carrying 0.4 μ cell culture inserts, so as to prevent direct contact between the T cells and NSPCs
Changes in NSPCs and T cells following co-culture is dependent on direct cell to cell contact
Flow cytometric analysis showed no significant changes in B7.1 and B7.2 levels on adherent NSPCs carrying cell culture inserts and co-cultured with encephalitogenic T cells when compared to those co-cultured with T cells belonging to sham group (Fig. 5b, c). qRT-PCR results also showed no significant changes in the levels of ICAM-1, TGF-βR1, TGF-β, and IFN-γ in NSPCs carrying cell culture inserts and co-cultured with encephalitogenic T cells when compared to those co-cultured with T cells belonging to sham or OVA-treated group (Fig. 5d-g). The levels of TGF-βR1 and FOXP3 in encephalitogenic T cells when co-cultured with NSPCs carrying inserts did not show significant changes when compared to T cells belonging to sham and OVA-treatment groups co-cultured with NSPCs carrying insert (Fig. 5h-i).
TGF-β and IFN-γ are required for NSPC-mediated conversion of encephalitogenic T cells into Tregs
As indicated from above results that co-culture with encephalitogenic T cells resulted in increased expression of TGF-β and IFN-γ in NSPCs, we wanted to know whether these two cytokines play important roles in conversion of encephalitogenic T cells into Tregs. NSPCs were first treated with neutralizing monoclonal antibodies against either TGF-β or IFN-γ or both for 24 h, and then co-cultured with encephalitogenic T cells. Neutralizing IFN-γ alone resulted in significant decrease in number of Treg in encephalitogenic T cells when compared to encephalitogenic T cells co-cultured with NSPCs without IFN-γ neutralizing antibody (p<0.01). Decrease in Treg number was also observed in encephalitogenic T cells co-cultured with NSPCs containing TGF-β neutralizing antibody, but this decrease was not statistically significant. Neutralizing both TGF-β and IFN-γ together in NSPCs resulted in decreased Treg numbers of encephalitogenic T cells co-cultured with them when compared to encephalitogenic T cells co-cultured without any neutralizing antibodies (p<0.01) (Fig. 6a).
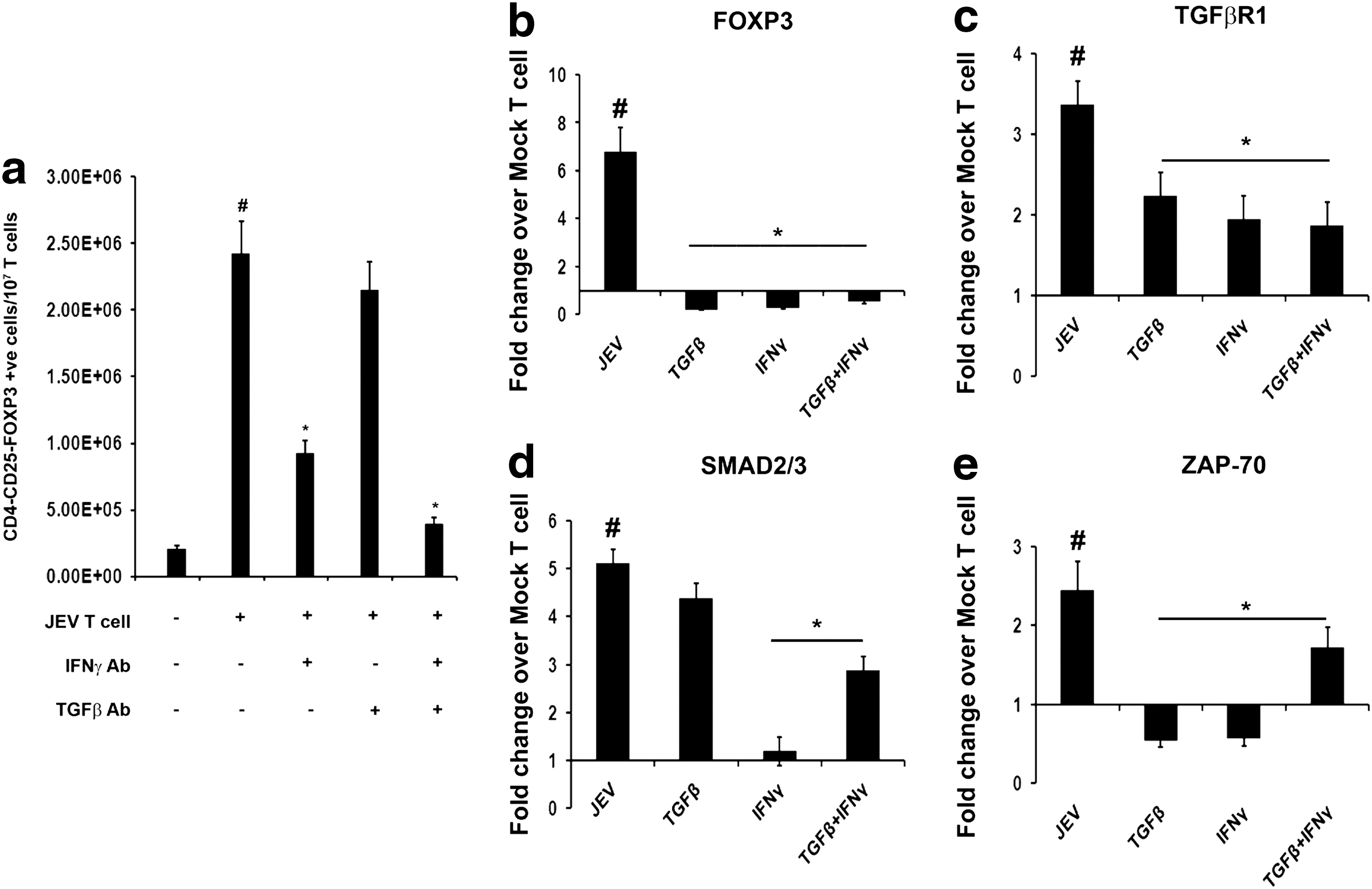
TGF-β and IFN-γ released from NSPCs are essential for Treg induction and molecular changes in T cells post co-culture. Graphical representation of number of T cells converted to Tregs isolated from mock-infected and JEV-infected animals' T cells, post co-culture with NSPCs that were treated with neutralizing antibodies against either TGF-β or IFN-γ or both
TGF-β and IFN-γ released from NSPCs are important for molecular changes in both NSPCs and T cells post co-culture
In order to observe the effects of TGF-β and IFN-γ neutralization on the surface expression of B7.1 and B7.2 on NSPCs following co-culture with T cells, flow cytometry was performed. Their levels were found to be significantly decreased in TGF-β neutralized NSPCs co-cultured with encephalitogenic T cells when compared to NSPCs without any neutralization that were similarly co-cultured with encephalitogenic T cells. Their levels were also significantly reduced in IFN-γ neutralized NSPCs and TGF-β+IFN-γ neutralized NSPCs when compared to NSPCs without any neutralization co-cultured with encephalitogenic T cells (Fig. 7a, b). qRT-PCR of ICAM-1 and TGFβR1 also revealed decreases in their levels in NSPCs (TGF-β neutralized) co-cultured with encephalitogenic T cells when compared to NSPCs (without any neutralization) co-cultured with encephalitogenic T cells (p<0.05). Similarly, their levels were also significantly reduced in NSPCs (IFN-γ neutralized) and NSPCs (TGF-β+IFN-γ neutralized) when compared to NSPCs without any neutralization, co-cultured with encephalitogenic T cells (p<0.05) (Fig. 7c, d). The levels of FOXP3, TGFβR1, SMAD2/3, and ZAP70 were significantly decreased in encephalitogenic T cells co-cultured with NSPCs containing TGF-β neutralizing antibody when compared to similar T cells co-cultured with NSPCs without any neutralization (p<0.05); similar decreases were also observed in encephalitogenic T cells co-cultured with NSPCs (IFN-γ neutralized) and NSPCs (TGF-β+IFN-γ neutralized) when compared to encephalitogenic T cells co-cultured with NSPCs without any neutralization (p<0.05) (Fig. 6b–e).
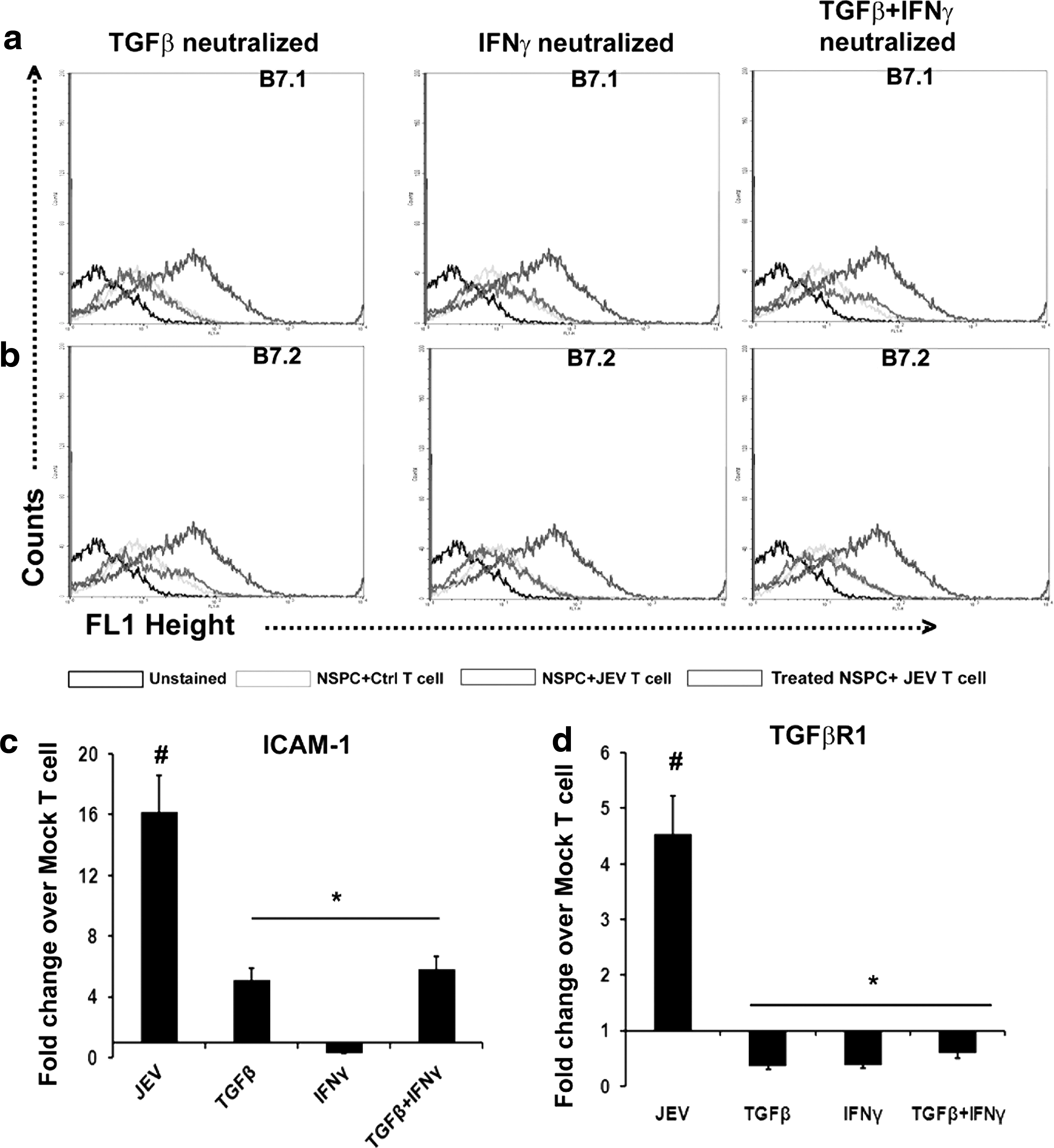
Molecular changes in NSPCs post co-culture with T cells is dependent on TGF-β and IFN-γ released from them. Flow cytometric staining for surface expression of B7.1
Discussion
Infiltrations of peripheral T cells into the CNS have been reported in many inflammatory diseases, including viral encephalitis such as JE (1,9,34). The brains' resident immune cells viz. the microglia along with the astrocytes form a complex immune interaction with these infiltrating cells (2,24), the result of which varies. On one hand there is exacerbation of the inflammation in the CNS, but on the other hand intrinsic mechanisms to control the inflammation are also activated. In an earlier study, we had reported that post infection with JEV, NSPCs in the brain acquire immune-competency and contribute to both the innate and adaptive immune axes; also there is significant decrease in their proliferation (5). It was also reported that these NSPCs release soluble factors that could induce T cell proliferation in the brain (8). In this current investigation, we show that noninfected NSPCs are capable of inducing conversion of antigen-primed T cells to a regulatory subset, which is indicative of anti-inflammatory behavior of these cells. In a well-characterized mouse model of JE, we observed presence of CD4+ and FOXP3+ cells in the SVZ region of the brain and the number was found to be increased after 8 days of infection compared to 4 days, probably to combat excessive inflammation. In order to ascertain whether these CD4+ and FOXP3+ cells in SVZ has any preventive role, we looked for these cells in SVZ of JEV-infected animals recovered from minocycline administration. To our surprise we did not find any CD4+FOXP3+ in their SVZ, probably because of no infiltration of peripheral T cells in CNS due to intact blood-brain barrier (36). Since the SVZ region of the brain is a known neurogenic niche, we were interested to study interactions between NSPCs and T cells in general or Tregs.
Our initial in vitro study where we cultured NSPCs in vitro and then co-cultured them with splenic T cells, either from mock-infected animals or from JEV-infected animals, showed very interesting results. We used splenic T cells as we had reported earlier that JEV is detected in the spleen by 4 days post infection (9). Thus, T cells isolated at this time point would be adequately primed with the virus in vivo. Here, we are referring to these cells as encephalitogenic T cells. We found that the number of Tregs was significantly higher when these encephalitogenic T–cells were co-cultured with naive NSPCs in comparison to that observed when those NSPCs were co-cultured with T cells, either from mock-infected or from OVA-treated animals. This increase was true, even considering the initial percentage of Tregs from all groups of animal prior to their co-culture with NSPCs. Increase in Tregs has also been reported in earlier studies using an EAE model. In one such study, myelin-specific regulatory T cells accumulated in CNS as a regulatory mechanism to control excessive inflammation (18). Increase in expression of co-stimulatory molecules and ICAM-1 was observed in NSPCs post co-culture with activated T cells. This observation was somewhat similar to the findings of an earlier study where such increases were reported in neurons, post co-culture with encephalitogenic T cells (16,22). Expression of co-stimulatory molecules are important for effective signaling with T cells. Thus, their overexpression is a clear indicator of a bi-directional interaction between the two cell types.
TGF-β and IFN-γ are well-known inducers of Tregs (13,37). Increase in TGF-β and IFN-γ was observed in NSPCs post co-culture with encephalitogenic T cells and their essentiality in Treg induction was demonstrated when neutralization of TGF-β or IFN-γ in NSPCs before co-culture resulted in decreased induction in Tregs. This induction in Treg from the encephalitogeic T cells post co-culture with NSPCs was found to be dependent on direct contact between either cell types. Prevention of direct contact using cell culture inserts resulted in no such changes in either NSPCs or activated T cells which were observed in experiments involving direct contacts indicating that functional co-stimulation was required in this process. Increase in the TGFβR1 levels was observed in NSPCs as well as on activated T cells. This increase in TGFβR1 levels in T cells was dependent on both direct contact and the levels of TGF-β and IFN-γ released from NSPCs, as neutralizing these cytokines in NSPCs before co-culture resulted in decreased TGFβR1 expression on T cells. TGFβR1 on NSPCs showed similar results, and this increase was found to be dependent on direct contact between both the types of cells and presence of TGF-β and IFN-γ. Levels of phosphoSMAD2/3, a signal transduction molecule involved in TGF-β signaling, was also increased in the encephalitogenic T cells post co-culture with NSPCs, and this increase could be dependent on TGF-β and IFN-γ levels in NSPCs. This increase in phosphoSMAD2/3 levels indicated towards increased TGF-β signaling in activated T cells post co-culture with NSPCs. This observation was in accordance with an earlier study that showed involvement of SMAD2/3 in induction of Treg through TGF-β (4). ZAP70 is an adaptor molecule involved in T cell receptor (TCR) signaling. Increase in the levels of phosphoZAP70 was observed in activated T cells post co-culture with NSPCs and this was further dependent on the levels of TGF-β and IFN-γ in NSPCs. This increase in ZAP70 levels may be due to increased TCR signaling post co-culture with NSPCs, but this observation needs to be investigated further, especially because we have seen earlier that NSPCs cultured from murine neurospheres do not express MHC-II (8).
Taken together our data suggest that NSPCs in inflamed CNS, where there is excessive infiltration of activated T cells, may try to suppress the inflammatory activity by inducing conversion of activated T cells into CD4+ -CD25+ -FOXP3+ Tregs. There have been several past efforts in characterizing the role of neural stem cells for their use in therapeutic purposes for the treatment of CNS inflammatory diseases (8,23,31,38). Our study adds a new vista to the current knowledge whereby it has been shown that NSPCs are capable of mounting an endogenous anti-inflammatory response. However, further studies are necessary to ascertain the degree of effectiveness of this process or their significance in vivo, before a therapeutic role of these cells is brought under consideration.
Footnotes
Acknowledgments
The authors would like to thank Manish Kumar Dogra and Kanhaiya Lal Kumawat for their technical assistance. AN is a recipient of the Senior Research Fellowship from the CSIR; KD is a recipient of a Research Associateship in Biotechnology and Life Sciences from the Department of Biotechnology (DBT), Government of India. AB is a recipient of the National Bioscience Award for Career Development-2010 from DBT. The authors have no conflict of interest to declare. The study conception and design was made by AN, KD, and AB. AN, IAM, and KK carried out experimental manipulations. Data were analyzed by AN and KD. The manuscript was written by AN, KD, and AB.
Author Disclosure Statement
No competing financial interest exists.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.