
Abstract
While the main reproduction aim of semen is the transport of spermatozoa to the female genital tract, seminal plasma is a complex fluid that also carries a broad array of immunologically active molecules. Seminal plasma has been shown to contain a diverse array of anti-inflammatory and pro-inflammatory soluble mediators that regulate immune responses within the female reproductive tract than can facilitate fertilization. Since the natural inflammatory response to semen deposition in the female genital tract may result in recruitment of activated HIV target cells into the female genital mucosa, we discuss the constituents of semen that may increase the risk for HIV infection in women.
Introduction
H
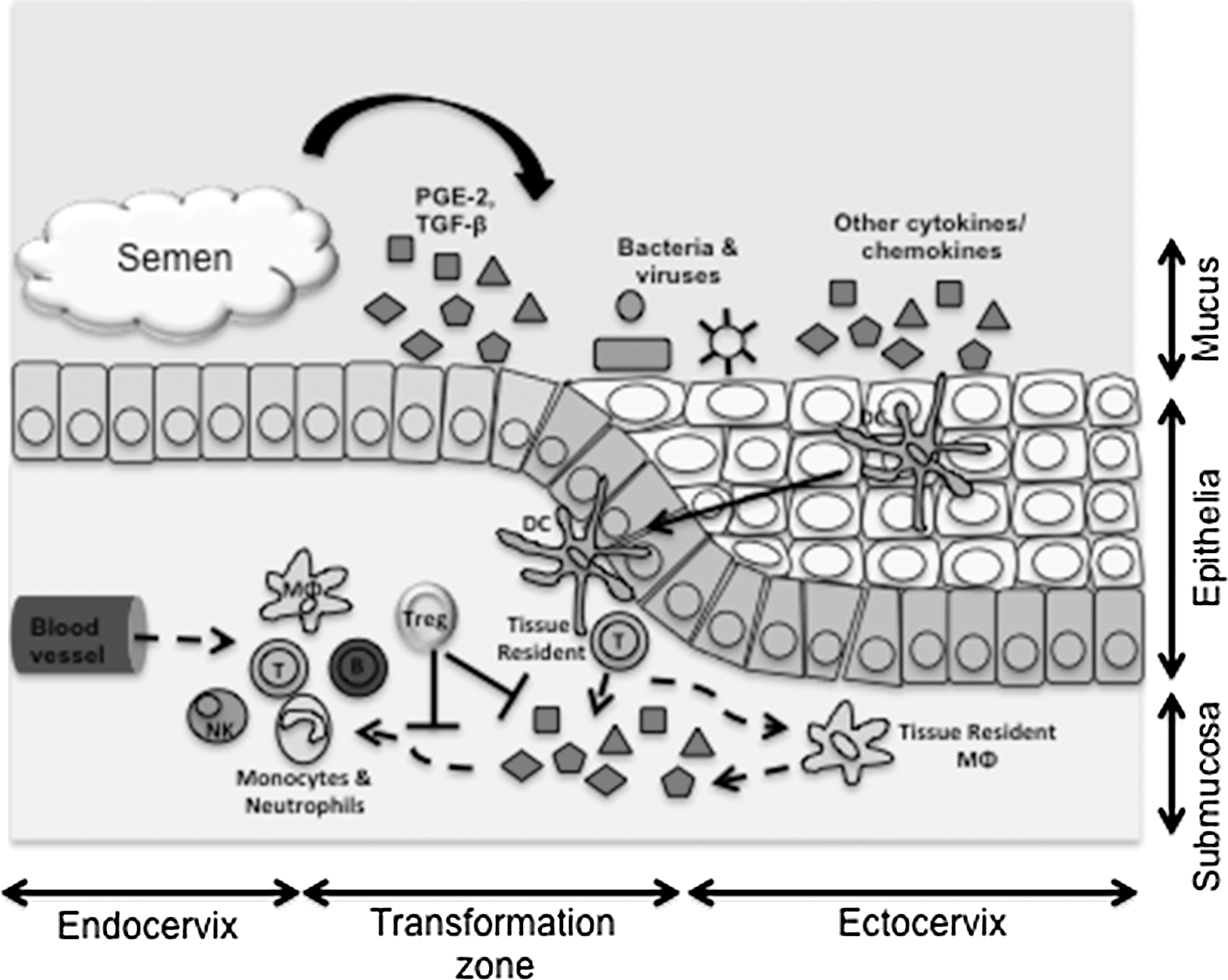
The effect of semen on HIV target cell recruitment. Semen deposition in the female genital tract has been shown to result in the release of multiple factors that have either direct or indirect immunomudulatory effects upon the genital epithelium. Sentinel dendritic cells (DC) are known to sample bacterial and viral products constantly that are presented to tissue resident CD4 T-cells. In turn, T-cells produce cytokines that activate tissue resident macrophages (MΦ) to produce cytokines and chemokines that causes a gradient-dependent migration/influx of T-cells, monocytes, neutrophils, NK cells, and B-cells. Mucosal resident T-regulatory cells (Treg) have been suggested to function as dampeners of this response.
In addition to viral load, several in vivo and in vitro studies have proposed a causal relationship between an individual's immune activation status during HIV infection and their risk of transmitting HIV to their partners (34,36). Given that semen can induce inflammatory responses and activate HIV susceptible cells within the female genital tract (65), it is plausible that, in addition to semen being the carrier of virus particles, its chemical composition may directly modulate HIV transmission risks. This review explores the impact that semen can have on aspects of female genital tract biology that might conceivably influence the odds of sexual HIV transmission.
The Female Genital Tract and Inflammation
The lower female genital tract consists of discrete anatomical compartments, including the vulva (consisting of keratinized stratified squamous epithelium similar to skin); the vagina (nonkeratinized stratified epithelium, which is aglandular); the ectocervix (a mucosal layer that is histologically similar to the vagina); the endocervix (consisting of simple columnar epithelium with a thickness of 10–30 μm, interspersed with pseudoglands); and the cervical transformation zone (defined as an abrupt transition between the endocervix and ectocervix) (53). Within the lower genital tract, physical, innate, and adaptive immune mechanisms protect against invading pathogens while tolerating commensal microorganisms (6).
The mediators of cellular immunity (including neutrophils, T-cells, antigen-presenting cells, B-cells and NK cells) in these regions have been well characterized in healthy women (53). CD8+ T-cells are more common in female genital tissues than CD4+ T-cells (44). In the absence of inflammation, the highest concentrations of these and antigen-presenting cells in the lower female genital tract are found within the cervix, with particularly high T-cell concentrations occurring in the transformation zone and surrounding tissues (53). Although the vagina has a larger surface area than the cervix, the epithelial tissues in this part of the genital tract have lower concentrations of T-cells and antigen-presenting cells than those of the cervix (53). Consequently, the cervix is considered as the primary inductive and effector site for immune responses within the female genital tract (27). The lower reproductive tract in women comprises the largest surface area (∼87.5 cm2) that is exposed to infectious HIV in semen (8,49).
Inflammation in the female genital tract may have both physiologic and pathologic consequences. Many of the normal reproductive processes in women display hallmarks of inflammation (23,63). Ovulation, menstruation, implantation, and parturition all induce tissue remodeling and injury of the genital tract mucosa that trigger inflammatory cascades (23,63). All of these events are associated with the upregulation of inflammatory mediators, including cytokines, chemokines, growth factors, and lipids, which collectively can alter the mucosal vascular components and impact immune function (23,63). These processes act in concert to facilitate the tissue remodeling and repairs that are necessary for the genital tract to carry out its reproductive functions.
Mucosal immune systems act as barriers against the outside world. They have to identify persistently and reliably the most appropriate targets among the large number of antigenic molecules that they encounter. While pathogen-derived antigens have to be targeted, antigens derived from commensal microbes and semen should ideally be tolerated. Most mucosal tissues achieve a steady-state regulatory balance that, while not harming the host, remains poised to mediate host immunity rapidly whenever necessary.
The female reproductive tract has a specialized mucosa that is required to both facilitate reproduction and provide protection against infection. Specifically, the mucosa of the female reproductive tract are required to accommodate prolonged exposure to HLA-mismatched developing embryos such that its immune functions require more extreme tolerance mechanisms than those of the gut and lung mucosa. For example, during the menstrual cycle, peak regulatory T-cell activity occurs around the time of ovulation (16)—the time that fertilization is most likely to occur. There are also changes in the Th1/Th2 balance during the menstrual cycle (14). Faas et al. (14) reported that immune responses in females become Th2-biased (defined by increased IL-4 production) during the luteal phase of the ovarian cycle. In the event of fertilization occurring, pregnancy is associated with additional immune regulation, including a bias toward Th2/Treg responses and away from the more inflammatory, graft-rejecting Th1 responses (76).
Female Genital Inflammation and HIV Risk
Heterosexual sex is the main mode of HIV transmission globally. Women are twice as likely as men to become infected with HIV during heterosexual sex (70), with the risk for HIV transmission for both sexes increasing with increasing plasma and genital viral loads (4,15). Although transmission rates are strongly dependent on plasma viral loads, local mucosal factors such as genital inflammation are significant predictors of an increased risk of infection via the genital mucosa (10,35,64). Further, although activation of the immune environment within the female genital tract is crucial for protective immunity, it may also facilitate HIV transmission by either disrupting the epithelial barrier and/or providing the virus with a stable supply of susceptible target cells (34).
To establish an infection via the female genital mucosa, HIV within semen must overcome substantial immunologic and physiologic barriers to cross the epithelial barrier and infect underlying HIV target cells (8). The three major targets of HIV-1 infection within the female reproductive tract are thought to be CD4+ T-cells, macrophages, and dendritic cells, which all express the CD4 receptor and the CCR5 co-receptor that are required for HIV infection (2,8,24). Although previous studies have argued that an intact squamous epithelial layer provides an impenetrable barrier against HIV infection, Carias et al. (8) demonstrated that through simple diffusion into the epithelium through interstitial spaces between cells, HIV particles are capable of penetrating intact vagina and ectocervical squamous epithelial layers up to a depth of 2–10 μm. HIV penetration has also been shown to be dependent on viral concentrations in the inoculum (8). Breaches in the epithelial barrier are most likely to be the most direct way for HIV to enter the genital tract, and this is evidenced by a number of studies demonstrating the effect of ulcerative STIs such as HSV-2 (29). the use of barrier-disrupting spermicides such as nonoxynol-9 (11), or vigorous penetrative sexual intercourse (48) in increasing risk of HIV transmission. Therefore, factors that alter the quality or integrity of the epithelial barrier, cause inflammation, or recruitment of activated HIV target cells into the mucosa may increase a women's risk of acquiring HIV.
Composition of Semen
Semen is a mixture of spermatozoa and seminal fluid, which is produced by the accessory glands along the male genital tract (prostate, seminal vesicles, and bulbourethral glands) (38). The pH of normal semen ranges from 7.2 to 7.8, and is more alkaline than the acidic environment in the female genital tract, which normally ranges in pH from 4.0 to 6.0 (22,75). In adults, semen ejaculate volumes vary from 0.1 to 10 mL (55).
Seminal fluid is a complex liquid comprised of proteins that are often antigenically distinct from the partner into which it is deposited. Amongst other constituents, it contains immunologic factors (including immunoglobulins, cytokines, chemokines, and growth factors), prostaglandins, enzymes, and angiogenic factors (69). The protein content of seminal plasma is between 35 and 55 mg/mL, and includes more than 900 distinct proteins (20,51). During adolescence, ejaculation is known to occur within 12 months after the onset of puberty, with first semen volumes being significantly lower than those measured from adult males (ranging from 0.5 to 1 mL). Early adolescent semen fails to liquefy and is jelly-like compared to that of adults. Liquefaction is an important process for fertility, when gel formed by proteins from the seminal vesicles is broken up and the semen becomes more liquid to facilitate sperm motility. As males age, their semen develops more mature characteristics, increasing in volume, containing increased quantities of normal sperm, and liquefying after a shorter period of time. Within 24 months of the first ejaculation, the semen volume and the quantity and characteristics of the sperm it contains match that of adult semen (30).
The cytokine environment within semen is quite distinct from both other mucosal sites and blood plasma. Specifically, it is dominated by high concentrations of TGF-β and PGE2, which both have potent immunosuppressive properties (26,32), including the suppression of immune cells such as neutrophils, NK cells, macrophages, and dendritic cells. While PGE2 levels in human semen are ∼100 μg/mL, it also contains higher concentrations of both the active and inactive forms of TGF-β (∼1 ng/mL and ∼80 ng/mL respectively) than any other body fluid (58,60 –62).
In addition to immunomodulatory agents, semen also contains a wide array of inflammatory, regulatory, adaptive, and hematopoetic cytokines (47). Relative to blood, semen from healthy HIV− men generally contains fivefold higher concentrations of MCP-1, IL-8, Fractalkine, GMCSF, IL-7, and IL-15 (47). At sites of inflammation, these cytokines are involved in the recruitment, maturation, and proliferation of monocytes, T-cells, B-cells, dendritic cells, and NK cells (1,9,17,37,41,68). Thus, semen passing through the male genital tract contains high concentrations of immune mediators involved in the recruitment, maturation, and survival of immune cells at this site.
Sexually transmitted infections (STIs) have an important influence on cytokine concentrations in the male genital tract (33,52,58). STIs are associated with elevated concentrations of G-CSF in seminal plasma. In turn, G-CSF concentrations correlate positively with those of the β-chemokines MIP-1α and MIP-1β, which have been implicated in the migration of potential HIV target cells into tissues of the male genital tract. G-CSF also potentially influences T-cell function and activates dendritic cells (56).
Semen and Female Genital Tract immunomodulation
The ability of semen from healthy males to inhibit various components of the immune response is important for human reproduction in that it allows the safe passage of foreign spermatozoa along the female genital tract (25,32,52,58,60). Given the evolutionary importance of reproduction, it is to be expected that regulation happens through several overlapping mechanisms. Conversely, seminal fluid is able to elicit pro-inflammatory responses in the female genital tract (5,12,33,42,58 –60,65), which are attributable either to the extended function of pro-inflammatory cytokines within semen or female immune responses to non-self antigens, histo-incompatibility, or allogeneic proteins within semen (5,42,59,60,65,66). Interactions between seminal fluid and female epithelial cells alter multiple gene expression pathways (65,66), with more than 700 genes being differentially expressed within ectocervical tissue biopsies from women who had had intercourse within 12 hours of the biopsy compared with those who had not recently had intercourse (65). The major gene pathways affected were those involved in immune cell trafficking, immune responses, pro-inflammation antigen presentation, and chemokine signaling (65,66). Analyses of semen-induced alterations in gene expression have been particularly informative regarding the mechanistic framework of the phenomenon known as the “seminal fluid-induced leucocyte-reaction” (31,46,59,65). Experiments in mice have shown that seminal fluid-induced alterations in inflammatory gene expression within the reproductive tract precede leukocyte recruitment (39,57,61). In vitro and human in vivo studies have further provided evidence that the seminal fluid component of semen is important for the induction of pro-inflammatory cytokines and chemokines such as GM-CSF, IL-6, IL-8, MCP-1, MIP-3α, and IL-1α (65).
One of the major cytokines in semen, the active form of TGF-β, causes a 70–99% decrease in basal transepithelial electrical resistance (a sensitive indicator of epithelial barrier integrity). Exposure of genital tissue to TGF-β apparently modifies the genital tissue distribution patterns of the tight junction proteins occludin and claudin-7 (50). The high concentrations of TGF-β in seminal plasma may act on tight junctions in the female reproductive tract to allow deeper penetration of HIV particles between interstitial spaces (8). This would potentially ensure closer contact between penetrating HIV particles and submucosal target cells.
The two major components of semen that have been suggested to be the main modulators of inflammatory signaling in female genital epithelial cells are TGF-β and PGE2 (7,67,71). By inducing an anti-inflammatory state in the female genital tract, TGF-β within semen is apparently particularly important for conditioning the maternal immune system to tolerate paternal antigens (58). In addition, the presence of TGF-β in semen has been shown to recruit CD4+CD8+FOXP3+regulatory T-cell (Treg) populations to the genital tract (3,26). Treg cells may further limit the female's allogeneic immune response, thereby promoting tolerance to paternal antigens and preventing the rejection of embryonic implantation (Fig. 1).
Semen and the Commensal Microbiota in the Female Genital Tract
Although bacterial, viral, and fungal pathogens are readily transferred from male to female partners via semen during sexual intercourse, the impact of semen exposure on the delicate balance of bacterial commensals in the female genital tract remains unclear. The most common medical condition that involves a perturbation in the vaginal microflora is known as bacterial vaginosis (BV) and is characterized by the change in the genital tract microbiome from predominantly Lactobacilli species to an ecology of other Gram-negative and positive bacteria (19). Rather than being a sexually transmitted disease, BV is generally thought to be a “sexually-enhanced” disease (73). Accordingly, sexual activity is the main factor associated with the occurrence of BV (21,73). In a study conducted by Eschenbach et al. (13), a significant increase in Escherichia coli and enteric Gram-negative rods was observed in the genital tract following a single episode of unprotected vaginal intercourse. This did not, however, lead to changes in vaginal Lactobacilli. Transient increases in vaginal E. coli colonization following intercourse have also been observed in other studies (18,28). However, increases in vaginal enteric Gram-negative rods (although not E. coli) have also been observed in women whose partners use condoms, suggesting that such intercourse-associated vaginal microflora changes rather than being attributable to seminal influx may be due to mechanical transfers of perineal enteric bacteria that is expected to occur irrespective of condom use (13,73).
Despite these findings, condom use might influence the microflora of the female genital tract in other ways. Relative to intercourse with a lubricated condom, intercourse without a condom may transfer more bacteria from the male periurethral area, the urethra, and under the foreskin to the vagina (13). Semen contains a diverse array of bacterial species derived from the urethra during ejaculation (74). In support of the notion that sharing of bacterial species between partners may occur, E. coli genotypes found in males have been known to cause urinary tract infections in their female partners (18).
In addition to the mechanical effects of sexual intercourse, semen exposure has been suggested to result in alkalinization of the vagina so as to promote a shift from Lactobacilli-dominated microflora to BV-type microflora. However, under normal circumstances, vaginal pH is only marginally increased following unprotected intercourse (13), and it is unclear what influence alkalinization has on BV. Another mechanism by which semen might contribute to the development of BV is by blocking the microbicidal activity of extracellular H2O2 that is produced by vaginal Lactobacilli. By shielding BV-causing microbes from H2O2, semen may promote their outgrowth (45).
Use of the long-acting injectable hormone contraceptive depot medroxyprogesterone acetate (DMPA) by women has been associated with an increased risk of HIV acquisition (43). In addition to causing epithelial thinning and drying, studies have also shown that women using DMPA have higher concentrations of pro-inflammatory cytokines in their genital secretions (including RANTES, IL-1β, IL-6, IL-8, MIP-3α, VEGF, and SLPI) (40). It is therefore possible that the impact of semen on the female genital tract may be further influenced by the contraceptive choice of the women. While several studies have suggested that DMPA use may increase risk of HIV infection (43) and influence the inflammatory environment in the female genital tract (40), DMPA has been shown to reduce BV incidence, possibly through hormone-dependent elevation of Lactobacilli concentrations (72).
Conclusion
Male seminal fluid may substantially alter the immunologic environment within the female mucosa. Besides inducing the expression of cytokines and chemokines that recruit and activate T lymphocytes, macrophages, and dendritic cells, seminal fluid may also promote an immune environment that is more tolerant of foreign antigens such as those found within semen itself and those expressed by the semi-allogeneic fertilized egg to allow implantation. More broadly, however, this altered immune environment may also impact both physiologic processes such as tissue remodeling during the reproductive cycle and pathophysiologic processes such as those involved in the breaching of the mucosal barrier by viruses such as HIV.
Footnotes
Acknowledgments
L.L., S.N., and J.P. received training in the United States as part of the Columbia University-Southern African Fogarty AITRP Program.
Author Disclosure Statement
No competing financial interests exist.