
Abstract
Although hantaviruses are not generally considered neurotropic, neurological complications have been reported occasionally in patients with hemorrhagic fever renal syndrome (HFRS). In this study, we analyzed innate immune responses to hantavirus infection in vitro in human astrocytic cells (A172) and in vivo in suckling ICR mice. Infection of A172 cells with pathogenic Hantaan virus (HTNV) or a novel shrew-borne hantavirus, known as Imjin virus (MJNV), induced activation of antiviral genes and pro-inflammatory cytokines/chemokines. MicroRNA expression profiles of HTNV- and MJNV-infected A172 cells showed distinct changes in a set of miRNAs. Following intraperitoneal inoculation with HTNV or MJNV, suckling ICR mice developed rapidly progressive, fatal central nervous system-associated disease. Immunohistochemical staining of virus-infected mouse brains confirmed the detection of viral antigens within astrocytes. Taken together, these findings suggest that the neurological findings in HFRS patients may be associated with hantavirus-directed modulation of innate immune responses in the brain.
Introduction
T
HFRS-associated hantaviruses cause vascular leakage and renal dysfunction, whereas HCPS-associated hantaviruses cause acute pulmonary edema (15,47). The mechanisms of hantavirus-induced pathogenesis are not fully understood, but both HFRS and HCPS share several clinical and pathologic features, including high fever, thrombocytopenia, increased capillary permeability and upregulation of tumor necrosis factor-α (TNF-α). Recent reports indicate less well-studied complications of HFRS and HCPS, such as central nervous system (CNS) dysfunction (10,19 –22,24,55). However, hantaviral antigens have not been detected in the brains of patients, so the mechanisms by which hantavirus infection might lead to neurological and neuropsychological complications are almost entirely unknown.
Both antiviral and pro-inflammatory responses to hantavirus infection play an important role in the modulation of host defense and disease manifestation (1,6,18,25,38,46,48). Pathogenic rodent-borne hantaviruses, such as HTNV, evade early innate immune responses by downregulating antiviral responsive gene expression immediately after infection. By contrast, upregulation of antiviral gene expression is observed in nonpathogenic Prospect Hill virus (PHV) infection (35). The pathogenesis of HFRS and HCPS is hypothesized to be mediated by increased pro-inflammatory responses (46). TNF-α, interleukin-1β (IL-1β), interleukin-6 (IL-6), chemokine (C-C motif ) ligand-5 (CCL-5), C-X-C motif ligand-8 (CXCL-8), and C-X-C motif ligand-10 (CXCL-10) are elevated in the kidneys and lungs of HFRS and HCPS patients, respectively, and in hantavirus-infected endothelial cells, macrophages, or epithelial cells in vitro (17,40,41,50). Although the major target organs of HFRS and HCPS are the kidney and lungs, respectively, the possibility that other organs, such as the brain, serve as target sites remains to be determined.
We recently determined that a novel shrew-borne hantavirus, Imjin virus (MJNV) (23), causes rapid and intense innate immune responses in human endothelial cells and macrophages, similar to that of the pathogenic rodent-borne HTNV (50). In this report, we demonstrate that both HTNV and MJNV show similar replication kinetics and efficient viral protein synthesis in human astrocytic cells. Distinct patterns of microRNA (miRNA) expression were also associated with HTNV and MJNV infection, and the top-predicted gene targets were found to play a role in inflammatory cytokine and chemokine pathways and integrin signaling. Moreover, HTNV and MJNV produced profound increases in cytokine and chemokine expression in brains of experimentally infected mice. Therefore, our data suggest that profound innate immune stimulation may be a potential underlying mechanism for CNS dysfunction in HFRS patients.
Materials and Methods
Cells lines
A human astrocytoma cell line (A172), derived from the brain tissue of a 53-year-old man with glioblastoma, was obtained from the Korean Cell Line Bank (Seoul, Korea) and propagated in RPMI-1640 (Lonza) supplemented with 5% heat-inactivated fetal bovine serum (FBS; Lonza), 10 mM HEPES, 2 mM
Viruses
HTNV strain 76–118 and MJNV strain 04–55 were used. Virus stocks and cells were determined to be free of Mycoplasma contamination by polymerase chain reaction (PCR) analysis. Infectivity titers of virus stocks were measured by plaque assay on Vero E6 cell monolayers, as previously described (48).
Animals and virus inoculation
Pregnant outbred ICR mice (Dbl Laboratory, Korea) were maintained in the vivarium at Korea University. Newborn ICR mice (<24 h after birth; 13–15 per group) were inoculated intraperitoneally with 1,000 plaque forming units (PFU) of HTNV or MJNV by the route. Mice were monitored daily for clinical signs, and were euthanized if moribund or at prescribed time points post-inoculation. The Institutional Animal Care and Use Committee approved all experimental animal protocols.
Virus replication kinetics
A172 cells were inoculated with HTNV and MJNV at a multiplicity of infection (MOI) of 0.5. After adsorption for 90 min at 37°C, cell monolayers were washed with phosphate buffered saline (PBS) and maintained in complete medium. Virus infectivity titers of supernatants were determined on each post-infection day by plaque assay (48).
Immunofluorescence microscopy
A172 cells, seeded on coverslips in 24-well plates, were inoculated with HTNV or MJNV at a MOI of 0.5 for 90 min at 37°C. Thereafter, fresh cell culture medium was added and incubated for 72 h. Slides were prepared as described previously (50) and examined by fluorescence microscopy, using a Zeiss Axioplan 2 microscope. Images were captured with a digital CCD camera (Hamamatsu).
Western blot analysis
Protein lysates were prepared as described previously (50). Briefly, HTNV- and MJNV-infected A172 cell lysates were separated on 10% SDS-PAGE and blotted onto PVDF membranes. HTNV and MJNV N proteins were detected with mouse monoclonal N protein-specific antibodies followed by anti-mouse horseradish peroxidase (HRP)-conjugated secondary antibody.
Quantitative real time reverse transcription PCR
RNA isolation and cDNA synthesis were described previously (50). Briefly, total RNA (1 μg), isolated with Trizol reagent (Invitrogen), was reverse transcribed using the RT system (Promega) for 1 h at 42°C. The resulting cDNA was used as template for real-time PCR quantification. Using the power SYBR Green supermix (Invitrogen), mRNA expression of human and mouse genes was measured. Human primers were previously published (14,50,60), and mouse primers are provided (Supplementary Table S1; Supplementary Data are available online at
Cytokine secretion measurement
Supernatants of HTNV- and MJNV-infected A172 cells were harvested at indicated times post-infection and stored at −80°C until testing. Cytokine levels were measured on a Magpix using a human cytokine Milliplex Kit (Millipore). Data analysis was performed with Bio-Plex Manager software (Bio-Rad Laboratories). The Magpix experiments were performed in duplicate.
miRNA microarray analysis
Total RNA was isolated followed by quality checks of both total RNA and small RNA using a 2100 Bioanalyzer and software capable of determining 28S and 18S ribosomal RNA ratio, total RNA Integrity Number, and small RNA and miRNA concentrations. Only samples with adequate total RNA and miRNA were used in the study. miRNA expression profiling was performed using the Agilent Human miRNA 8 X 60K (V16). The microarray was designed based on Sanger miRBase (release 16.0) and contained probes for 1,205 human and 144 human viral miRNAs. One hundred ng of total RNA was labeled using the Agilent miRNA Complete Labeling and Hybridization Kit, according to the manufacturer's instructions. For target gene functional annotation, we chose to use GO categories, canonical Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways maps, and Panther protein classification tools. Ingenuity Pathways Analysis (IPA; Ingenuity Systems, Inc.) was used to analyze miRNA expression data in the context of known biological response, regulatory networks, and specific pathways.
Immunohistochemistry
Brain tissues were fixed in 4% paraformaldehyde and embedded in paraffin. Paraffin-embedded, 5 μm thick sections were deparaffinized with xylol, rehydrated using decreasing concentrations of ethanol (100%, 95%, 80%, and 70% for 5 min each). Following antigen retrieval in 10 mM sodium citrate buffer, pH 6.0 at 95–100°C for 20 min, peroxide-blocking solution was applied to slides for 20 min at room temperature. Antigen detection was performed using anti-HTNV and anti-MJNV monoclonal mouse IgG antibodies followed by antibody enhancer and polymer AP (GBI Polink-2 Plus kits; GBI Lab); subsequent color development was performed with AP+red (GBI Labs). For double staining of glial fibrillary acidic protein (GFAP), preblocking solution (10% normal goat serum) was applied to the slides and incubated for 10 min at room temperature. Anti-GFAP primary antibody, at the recommended concentration (1:500; Millipore), was incubated for 2 h at room temperature. Slides were washed in 1× TBST four times for 5 min and antibody enhancer and polymer HRP detection kit was applied (GBI Polink-2 Plus kits). After washing the slides four times, color development was performed with Emerald Green (GBI Labs). The slides were mounted with aqueous mounting media.
Statistical analysis
The paired differences of the experimental groups were compared using the nonparametric Mann–Whitney U-test. A p-value of <0.05 was considered statistically significant (Graph-Pad; Prism software).
Results
Replication kinetics of HTNV and MJNV in A172 human astrocytic cells
To study the immunological responses in vitro, we first determined if the A172 human astrocytic cell line could be infected with hantaviruses. Virus-specific fluorescence was detected in A172 cells at 72 h following HTNV or MJNV infection at a MOI of 0.5 (Fig. 1A). Infectivity titers of HTNV and MJNV in A172 cells, at a MOI of 0.5, attained 103–104 PFU/mL at day 2 and persisted until day 5 post-infection (Fig. 1B). Nucleocapsid (N) protein synthesis in HTNV- and MJNV-infected A172 cells, as measured by Western blot analysis, was elevated early and persisted until day 5 post-infection (Fig. 1C).
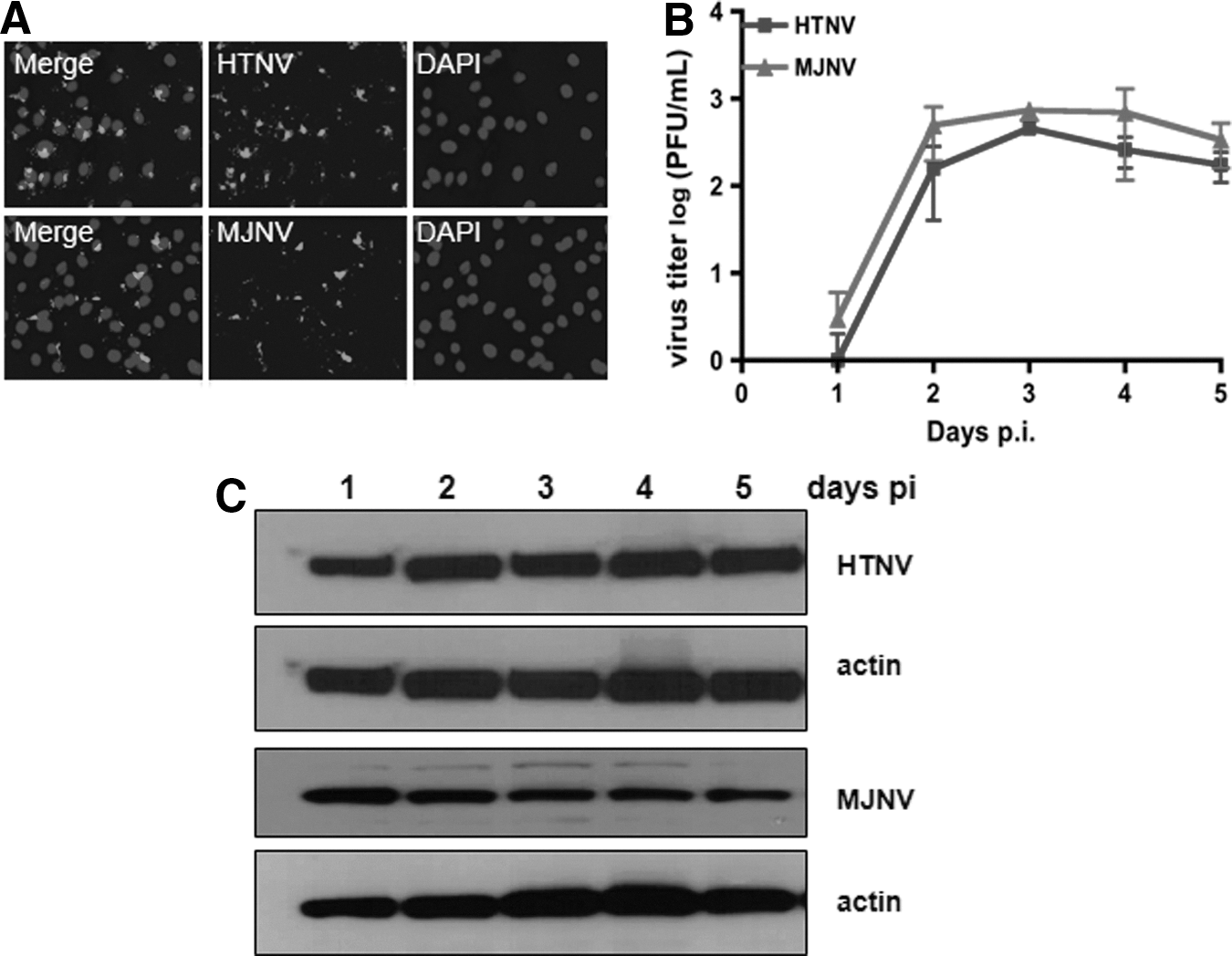
Hantavirus infection in human astrocytic cells.
Antiviral and pro-inflammatory responses in HTNV- and MJNV-infected A172 cells
To determine if astrocytes can induce antiviral responses to hantavirus infection, transcripts for myxovirus resistance gene A (MxA), IFN-β, IL-28 (IFN-λ2), and IL-29 (IFN-λ1) were quantitated by real-time reverse transcription PCR (qRT-PCR) at 1, 3, and 5 days post-infection. MxA and IFN-β gene expression peaked at day 3 post-infection, and decreased at day 5 post-infection in cells infected with HTNV or MJNV (Fig. 2A and B). In contrast to the low expression levels of MxA and IFN-β at day 1 post-infection, the induction of type III IFN (both IL-28 and IL-29) preceded induction of MxA and IFN-β by 2 days (Fig. 2C and D). Thus, these data indicate that type III IFN (IFN-λ) expression is induced prior to type I IFN (MxA and IFN-β) during HTNV and MJNV infection.

Kinetics of antiviral and pro-inflammatory gene expression during hantavirus infection. A172 cells were infected with HTNV (black) or MJNV (light grey) (MOI of 0.5) or mock infected. RNA was extracted at days 1, 3, and 5 post-infection. The mRNA expression of
To determine the effect of HTNV and MJNV infection on the pro-inflammatory responses, mRNA expression of interferon gamma-induced protein-10 (IP-10), Regulated upon Activation, Normal T-cell Expressed, and Secreted (RANTES), vascular endothelial growth factor (VEGF), and interleukin-8 (IL-8) were measured at 1, 3, and 5 days in A172 cells infected with either HTNV or MJNV. Both HTNV and MJNV led to elevated levels of IP-10, RANTES, VEGF, and IL-8 at 3 days post-infection (Fig. 2E–H). In general, HTNV-infected cells show decreased detection of IP-10, RANTES, and IL-8 at day 5 post-infection. However, MJNV-infected cells generally show slight increases in detection of these markers at day 5 post-infection. Next, the levels of secreted cytokine and chemokine proteins were measured in supernatants of HTNV- or MJNV-infected A172 cells. Consistent with the qRT-PCR results, the protein secretion levels of IP-10 and VEGF increased upon infection with HTNV and MJNV (Fig. 3).
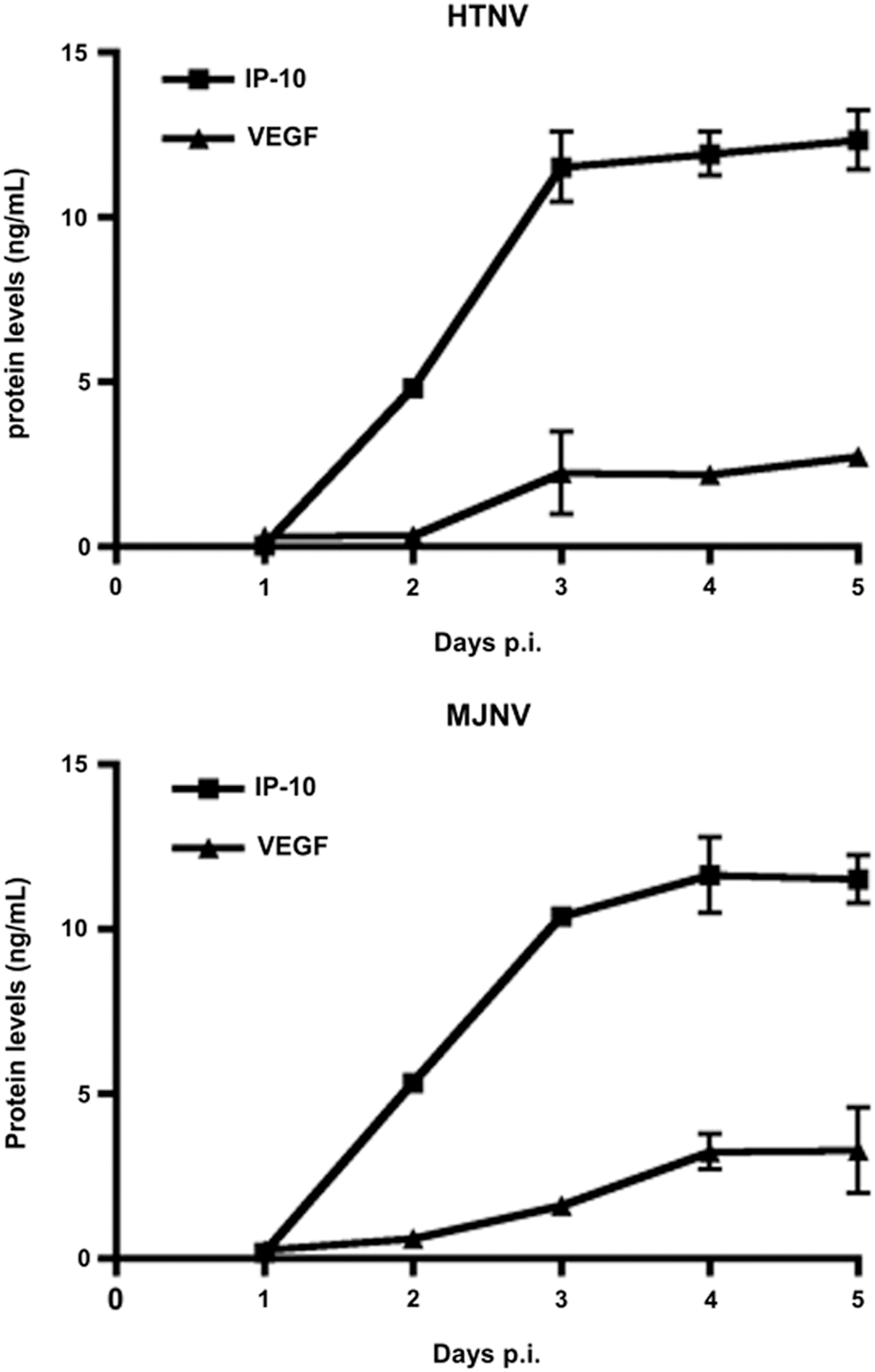
Cytokine secretion following HTNV or MJNV infection in A172 cells. Supernatants of HTNV- and MJNV-infected A172 cells were harvested at indicated times post-infection, and inflammatory molecule levels of IP-10 and VEGF were determined by Magpix analysis in supernatants of A172 cells infected with HTNV or MJNV at a MOI of 0.5, or mock infected and harvested from days 1 to 5 post-infection. The Magpix experiments were performed in duplicate, and the results are representative of two independent experiments.
Distinct cellular miRNA expression patterns in HTNV- and MJNV-infected A172 cells
To identify differentially expressed miRNAs associated with HTNV and MJNV infection, we compared global cellular miRNA expression profiles in HTNV- and MJNV-infected A172 cells at matching time points. miRNAs that were altered at least twofold were considered significant. Heat maps showed the top 10 up- and downregulated miRNAs induced by both HTNV and MJNV-infected samples (Fig. 4A and Supplementary Table S2). Analysis of 1,205 human miRNAs revealed that HTNV upregulated the expression of 90 miRNAs and downregulated expression of six miRNAs by twofold, whereas MJNV upregulated expression of 64 miRNAs and downregulated expression of two miRNAs by twofold (Fig. 4B). A total of 3,364 target genes were upregulated, whereas 2,803 target genes were downregulated by selected miRNAs. To analyze more fully the top biological functions associated with identified targets, we performed a Gene Ontology (GO) term and KEGG pathway annotation, using the DAVID gene annotation tool. Our pathway-based analysis suggested that the top three putative targets of miRNAs regulated by HTNV and MJNV infection were involved in Wnt, inflammatory cytokine and chemokine production, and integrin signaling (Table 1).
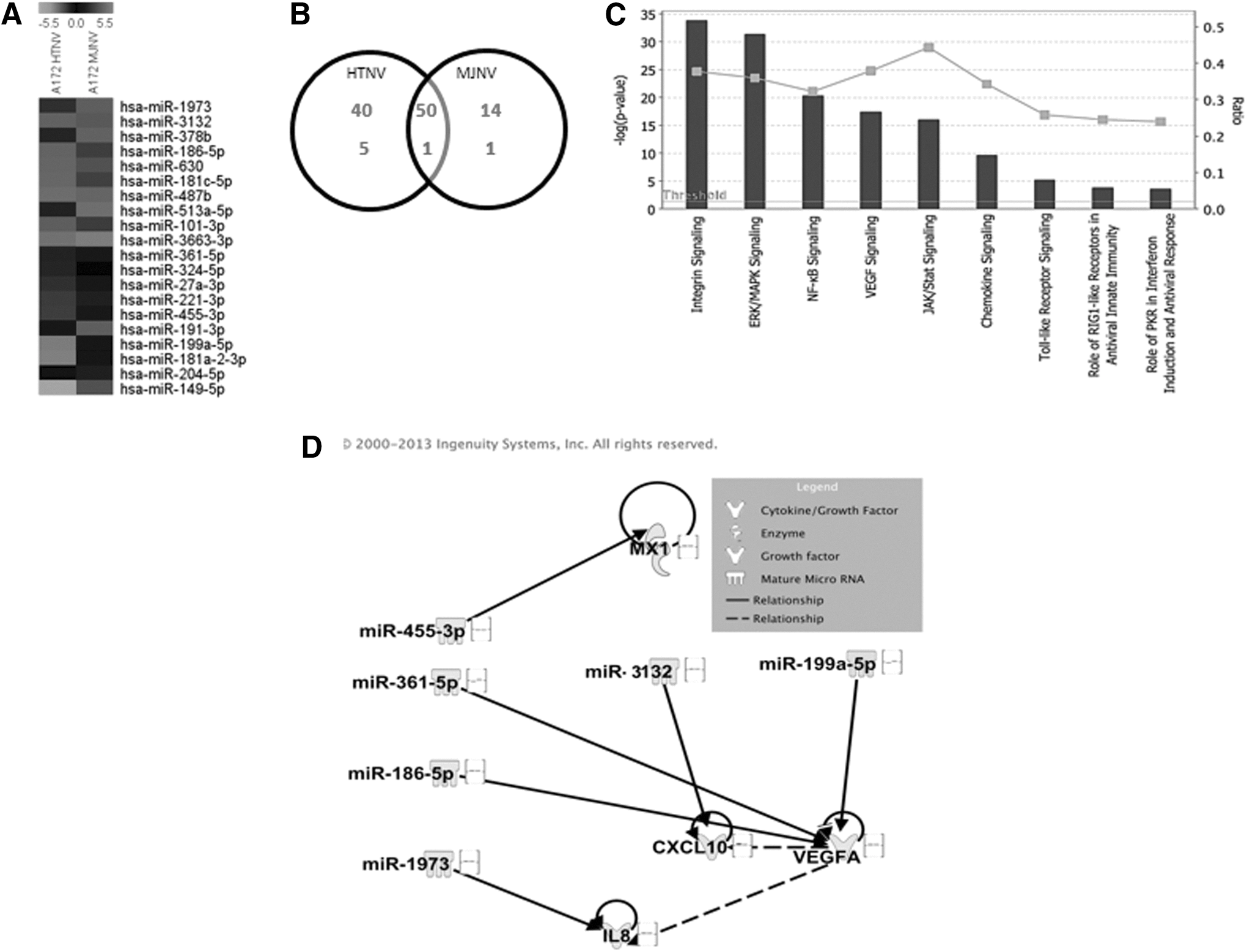
Cellular miRNA expression patterns in HTNV- and MJNV-infected human astrocytic cells.
GO, gene ontology.
To understand better the roles of miRNAs expressed in HTNV- and MJNV-infected cells, Ingenuity Pathway Analysis (IPA) was used to generate gene lists known to have direct or indirect functional relationships with these miRNAs. Using this approach, we were able to show that HTNV- and MJNV-modulated miRNAs and their targets regulated the innate immune response, antiviral response, and pro-inflammatory signaling pathways (Fig. 4C). Furthermore, our network analysis demonstrated a potential correlation between HTNV- and MJNV-modulated miRNAs and mRNA expression data of antiviral or inflammatory molecules (such as MxA, IL-8, IP-10, and VEGF; see Fig. 2E–H) induced by these viruses in the same cell type and same time after infection, suggesting their role in inducing inflammation after HTNV and MJNV infection (Fig. 4D). Moreover, the similar expression profiles of specific miRNAs in HTNV- and MJNV-infected A172 cells suggested that these two viruses might cause similar cellular pathology.
Immunological responses in the brains of HTNV- and MJNV-infected suckling ICR mice
Suckling ICR mice (<24 h of age), inoculated intraperitoneally with 1,000 PFU of HTNV or MJNV, developed rapidly progressive, severe disease, characterized by weight loss, ruffled fur, and reduced activity, followed by neurological signs, including paralyses and seizures. The onset of disease was more rapid with MJNV than HTNV. That is, clinical signs appeared at 8–9 days post-inoculation in MJNV-infected mice, compared to 12–13 days in HTNV-infected mice, and death ensued within 2 days of disease onset (Table 2).
Two independent experiments were performed for each group.
HTNV, Hantaan virus; MJNV, Imjin virus; PFU, plaque forming units; PBS, phosphate buffered saline.
The transcriptional levels of cytokines and chemokines that might play a role in attracting inflammatory cells to affected tissues was assessed by measuring the expression of antiviral responsive genes, including myxovirus (influenza virus) resistance-1 (Mx1), IFN-β, IFN-λ, 2′-5′-oligoadenylate synthetase (OAS), and pro-inflammatory response genes, including IP-10, RANTES, macrophage/monocyte chemotactic protein-1 (MCP-1), and TNF-α in the brain, kidney, lungs, liver, and spleen of HTNV- and MJNV-infected suckling mice, using qRT-PCR. All four antiviral genes were significantly elevated in the brains of HTNV-infected (p<0.05), and IFN-β and IFN-λ gene expressions were also significantly increased in the brains of MJNV-infected (p<0.05) compared with mock-infected control mice. IFN-β and IFN-λ expression was elevated by ∼1 log in the virus-infected groups, whereas Mx1 and OAS expression was 2–3 logs higher in the virus-infected groups (Fig. 5A–D). Along with upregulation of antiviral responsive genes, the levels of IP-10, RANTES, MCP-1, and TNF-α were significantly increased in brains of HTNV-infected mice (Fig. 5E–H). In examining the innate immune responses in other tissues of HTNV- and MJNV-infected mice, we found that the expression of antiviral and pro-inflammatory genes was increased, but the level of induction was not as high as that in the brain (Fig. S1). For example, a 4–6 log-fold higher elevation of IP-10 and RANTES was found in the brain compared to <1–2 log increase in the kidney, lungs, liver, and spleen.
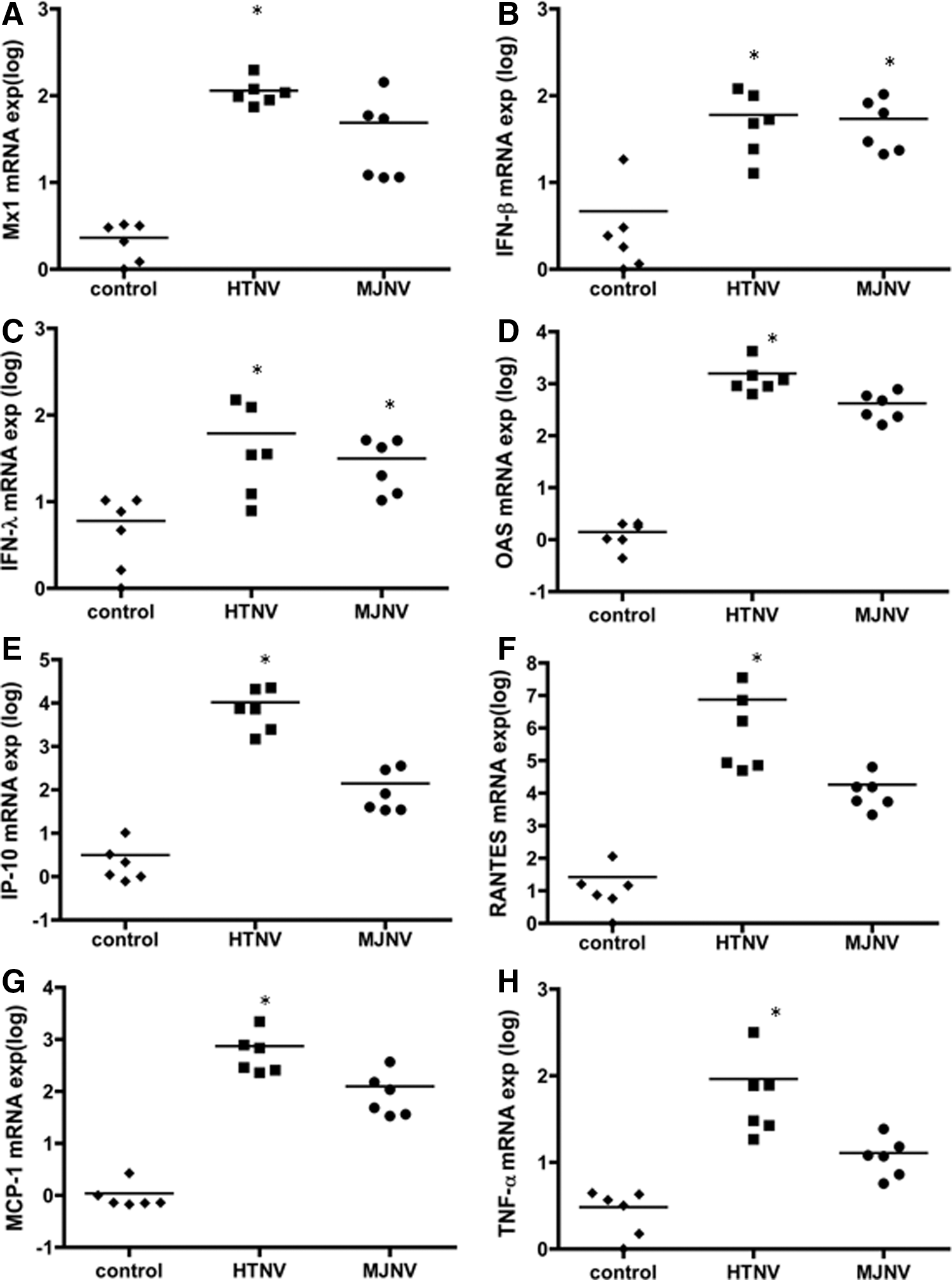
Hantavirus-induced antiviral and pro-inflammatory gene expression patterns in vivo. Suckling ICR mice were injected intraperitoneally with HTNV or MJNV and sacrificed 9 (MJNV) or 13 (HTNV) days after infection. Total RNA was extracted from the brain, and the expression of
To ascertain which cell types were targeted in the brain of hantavirus-infected mice, antibodies against GFAP were used to identify astrocytes. Also, antibodies against HTNV and MJNV N protein were used to localize virus in the brain of HTNV- and MJNV-infected mice. Viral N protein was detected extensively throughout the brain and appeared to be specifically associated with GFAP-staining astrocytes (Fig. 6).
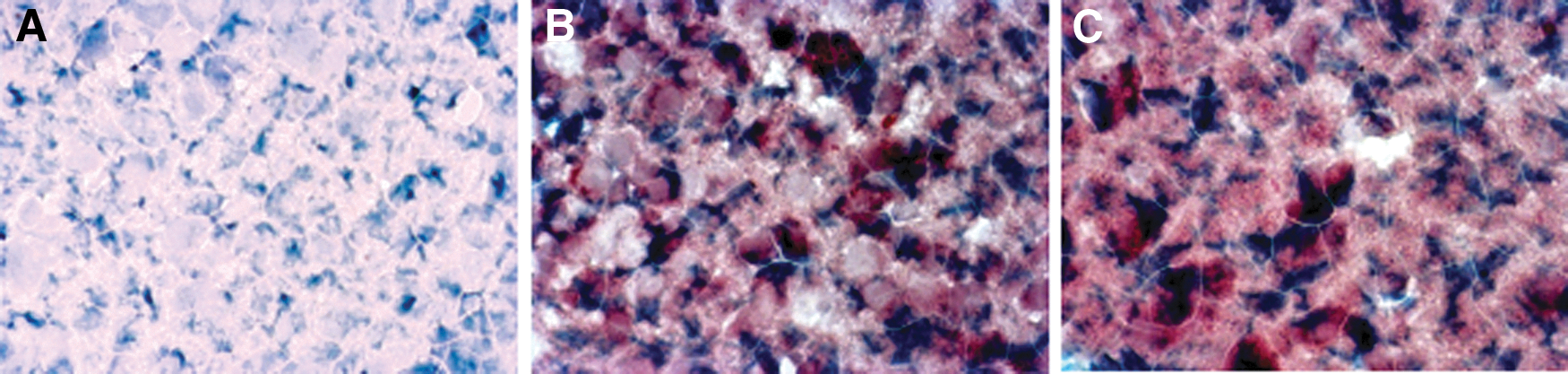
Immunohistochemical detection of GFAP and viral N protein in the brain of HTNV- and MJNV-infected mice. Robust cytoplasmic staining of GFAP and hantavirus N protein was found in brains of mice experimentally infected with HTNV and MJNV.
Discussion
While renal insufficiency is the principal target-organ manifestations of HFRS, viral antigens can be found in the brains of animal models (59) and CNS-associated clinical signs and symptoms have been reported in some HFRS patients (10,19 –22,24,55). Our data demonstrate that HTNV and MJNV efficiently replicate and induce N protein synthesis in human astrocytic cells and similarly cause the induction of innate immune responsive genes. Also, in suckling ICR mice, HTNV or MJNV elicited production of high levels of pro-inflammatory cytokines and chemokines in the brain, which were associated with an acute, fatal neurological disease. Collectively, these data suggest that the neurological complications associated with HFRS may be triggered by innate immune stimulation in the brain, following infection with either known pathogenic rodent-borne hantaviruses or newfound still-orphan shrew-borne hantaviruses.
We recently reported that HTNV and MJNV replicated to high titer in both human umbilical vein endothelial cells (HUVEC) and macrophages (THP-1) and was associated with increased expression of antiviral responsive genes in a time-dependent manner (50). As in HUVEC and THP-1, HTNV and MJNV replicated in A172 cells with similar kinetics and efficiency. Hantavirus infection in A172 cells was accompanied by type I and type III IFN responses and increased production of pro-inflammatory mediators, such as IP-10, RANTES, and VEGF. It has been previously demonstrated that hantaviruses induces the expression of type I IFN (α/β), type III IFN-λ, and interleukin-28 and -29 (IL-28 and IL-29) (56). Type I IFN has been demonstrated to play a central role in the regulation of innate immune responses and in the control of hantavirus infection in vitro (1,35). Interestingly, type III IFN was upregulated more rapidly at 1 day post-infection than type I IFN in A172 cells, although this was not sufficient to control viral replication at early time points.
Mounting evidence suggests that specific miRNA expression profiles may be associated with clinical outcomes or treatment responses in human diseases (13). We also previously reported distinct changes in miRNA expression profiles in a hantavirus species-specific manner from human endothelial, epithelial, and macrophage cells (49). In this study, we demonstrated that similar miRNAs were up- or downregulated in HTNV- and MJNV-infected A172 cells, and the top three putative targets of miRNAs appeared to be involved in Wnt, inflammatory cytokine/chemokine production pathways, and integrin signaling, which were previously shown to be dysregulated in pathogenic hantavirus infection (1,16). Moreover, expression-pairing analysis using Ingenuity demonstrated that HTNV- and MJNV-modulated miRNAs can target inflammatory genes induced by these viruses in A172 cells. Thus, our miRNA expression analysis confirms previous reports and these dysregulated innate immune responses may be attributed in part to the miRNA-mediated expression changes of target genes.
In their natural rodent reservoir hosts, hantaviruses cause asymptomatic and persistent infections with no apparent pathology. However, suckling ICR mice, inoculated with HTNV develop lethal CNS infection (31). Thus, we utilized suckling ICR mice to study innate immune responses. Both HTNV and MJNV infection caused similar pathology in suckling mice, but there were minor differences in disease kinetics. That is, MJNV caused disease and death more rapidly than HTNV. Furthermore, higher induction of pro-inflammatory responsive genes was found in the brains of HTNV-infected mice compared to MJNV-infected mice. It is possible that the more pronounced cytokine and chemokine response in HTNV-infected mice may have been protective in delaying pathology.
In conclusion, this study indicated that MJNV triggered distinct activation of innate immune responses, resembling that of HTNV, in human astrocytic cells and mice brain. The differentially expressed miRNAs detected by microarray analysis provide potential candidates of viral infection-related host cellular miRNAs. Although the disease kinetics was accelerated in MJNV-infected suckling mice, the severity of CNS-mediated paralysis was similar to HTNV. Taken together, these findings suggest that distinct innate immune responses occur in the brain following hantavirus infection and may account for the neurological signs and symptoms in some HFRS patients.
Footnotes
Acknowledgments
This work was supported by a grant from the Agency for Defense Development (UE134020ID), and by grants from Korea University, the National Institutes of Health (R01AI075057, P20GM103516), and the Armed Forces Health Surveillance Center, Division of Global Emerging Infections Surveillance and Response System (AFHSC-GEIS).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.