
Abstract
The adaptive immune system plays an important role in response to interferon plus ribavirin treatment of hepatitis C virus (HCV) infection. Cytokines play a significant role in the adaptive immune system. The production of cytokines may be regulated by single nucleotide polymorphisms (SNPs). This study was designed to examine the correlation of some important SNPs of cytokines with interferon plus ribavirin treatment of HCV infection in the Pakistani population. We followed 140 chronic HCV-infected patients in our study. All of these patients had completed their planned course of interferon plus ribavirin treatment. We also considered 120 healthy subjects as controls. The detection of interleukin-18 (IL-18) SNPs was performed by tetra-primers amplification-refectory mutation system polymerase chain reaction, while for genotyping of osteopontin (OPN), transforming growth factor beta (TGFβ), and N-acetylgalactosaminyltransferase 8 (GALNT8) SNPs, allele-specific polymerase chain reaction was performed. The distribution of the IL-18 −607AA genotype varied significantly between healthy control and patient groups. Its distribution was significantly high in healthy subjects than HCV patients (p=0.031), signifying its potential involvement in the natural clearance of HCV infection. The occurrence of the −607AA genotype of IL-18 was also significantly higher in the sustained virological group (SVR) than in the nonresponder (NR) group (p=0.046), highlighting its protective involvement in the treatment outcome of chronic HCV infection. The frequency of the OPN −442TT genotype was higher in the SVR group than in the NR group (p=0.034), indicating a significant possible role of this genotype in therapy for HCV infection. No important association was found between TGFβ and GALNT8 genotypes and the natural clearance and treatment response of HCV infection. IL-18 −607AA and OPN −442TT genotypes can be used as positive predictive markers of interferon plus ribavirin treatment of HCV infection in the Pakistani population.
Introduction
H
Interferon-based therapy is mostly effective against HCV genotype 3, which is the most highly distributed genotype in the Pakistani population (2). Interferon-based therapy decreases serum HCV-RNA levels in a biphasic pattern. The rapid decline of HCV-RNA levels within 1–2 days might be attributed to the direct interaction of interferon on HCV production. On the other hand, the decline of HCV-RNA levels in the second phase might be attributed to cell-mediated immune response (35). The decline of HCV-RNA levels in SVR patients is more prominent than non-SVR patients, indicating that the efficacy of interferon-based therapy is not only dependent on viral factors but also on host factors (54).
Previously it has been shown that vulnerability to HCV infection and response to interferon plus ribavirin therapy is regulated at different function levels, such as the production of cytokines by various cells, single nucleotide polymorphisms (SNPs) of different genes, and antigen presentation (11,28,47). Cytokines are secreted by most of cells, for example immune cells, endothelial cells, fibroblasts, and a range of stromal cells. Cytokines are important regulators of immune responses. There are inter-individual variations in cytokines production, which may be attributed to SNPs in the DNA of these cytokines. These variations influence the balance between pro-inflammatory and anti-inflammatory cytokines, and might affect the severity as well as the treatment outcome of HCV infection (6).
Interleukin-18 (IL-18) is an inflammatory cytokine that is largely secreted by monocytes, macrophages, and immature dendritic cells. IL-18 synergizes the effect of IL-12 and induces a Th-1-mediated immune response, while in the absence of IL-12, it induces a Th2-mediated immune response. It also activates the secretion of two other important cytokines: tumor necrosis factor alpha (TNF-α) and interferon gamma (IFN-γ). TNF-α regulates the secretion of IL-18 by stimulating the secretion of IL-18 binding protein (IL-18BP) (53). Interferon therapy of chronic HCV patients leads to a 3–24-fold increased production of IL-18BP to regulate inflammation and fibrosis development (31). Generally, the production of IL-18 is considerably higher in chronic HCV patients, suggesting its critical function in disease progression (53). The enhanced production of IL-18 is associated with hepatic injury (48) and poor treatment response to interferon therapy (3). It has been established that IL-18 production remains low in SVR patients during treatment (52). Enhanced secretion of IL-18 and IL-18BP has been reported in Chickungunya virus patients as compared with controls. Thus, virus infection leads to increased production of IL-18 and IL-18BP (9). Earlier studies have reported that IL-18 production is mainly dependent on IL-18 genetic variants −607C/A (rs1946518) and −137G/C (rs187238). These SNPs are linked with interferon plus ribavirin therapy of HCV (30), HBV (24,51), and human immunodeficiency virus (HIV) infection (41). We selected these two SNPs to find their correlation with natural clearance and treatment outcome of HCV infection in our population.
TGF-β is an anti-inflammatory cytokine that suppresses the development and cytotoxicity of natural killer (NK) cells. It blocks the secretion of IFN-γ and IL-12. SNPs of TGF-β are responsible for its variable production (18), liver cirrhosis (37), and natural virus clearance (25). We considered the correlation of TGF-β functional variants at codon 10 T/C (rs1982073) and codon 25 G/C (rs1800471) with HCV infection and treatment outcome.
OPN is an extracellular matrix (ECM) that is mainly produced by bone and kidney. It has a wide range of biological activities, which involve cell adhesion, proliferation, and migration (16,44). Recently, the involvement of OPN promoter variant in HCV infection has been investigated. It was found that elevated levels of OPN mRNA and protein expression were associated with hepatocellular carcinoma (20). Similar results for HBV infection were also found (45). It was concluded that increased plasma level of OPN is a prognostic marker of hepatocellular carcinoma, in both HCV and HBV infection (49,55).
OPN plays a crucial role in liver fibrosis during the process of tissue remodeling (7). ECM is mainly deposited in subendothelial space by hepatic stellate cells (HSC) (8,40). It has been demonstrated that OPN is significantly involved in the activation of HSC, as incubation of OPN with HSC leads to cell proliferation and migration. Additionally, it has been shown that OPN directly induces the production of collagen and type II TGFβ receptor from HSCs, both at transcriptional and translational levels. Thus, OPN has a key function in hepatic fibrosis and injury (29). We studied the influence of OPN promoter polymorphism −442C/T (rs11730582) on natural clearance and interferon-based therapy of HCV infection.
Probable polypeptide GALNT8 is considered to catalyze the early reaction in the biosynthesis of O-linked oligosaccharide. It transfers an N-acetyl-D-galactosamine residue to the protein receptor. A single nucleotide variant in intron 6 of GALNT8 (rs10849138) is shown to be involved in the treatment response of combinational therapy for HCV infection in the Japanese population (34). We studied the association of this particular polymorphism of GALNT8 with HCV infection in our population.
The main objective of this study was to find the distribution of important SNPs of cytokines regarding chronic HCV infection, their association with natural clearance, and treatment response in the Pakistani population.
Materials and Methods
Patients and healthy controls
We considered 140 patients in our study. Patient blood sampling was performed in Saidu Group of Teaching Hospital (Khyber Pakhtun khwa) and the Gambat Institute of Medical Sciences (Sindh). Seventy-five patients were from Swat and its vicinity, and 65 patients were from Gambatt and its vicinity. The inclusion criteria were the detection of HCV-RNA by reverse transcription polymerase chain reaction (RT-PCR). HCV-RNA quantification was performed with the MyiQ2 two-color real time PCR detection system (BioRad) by using the Gene Proof Hepatitis C Virus PCR Kit designed for the detection of HCV-RNA. Patients with other infectious diseases, for example HBV, HIV, tuberculosis (TB), malaria, or autoimmune liver disease, were not included in this study. Chronic HCV subjects possessing genotypes 1, 4, and co-infection were administered treatment of interferon alpha 2a and ribavirin for 1 year, while all others were mostly administered treatment for 6 months. Patients who failed to complete a planned course of treatment for any reason were not included in the study. We also enrolled 120 unrelated healthy adult subjects in this study as controls. All these subjects were from Islamabad, having no previous history of infectious HBV or HCV infection. The study was agreed by the Ethic Committee of the institute, and written permission was taken from all enrolled subjects for the detection of genetic variations of different cytokine genes.
PCR
DNA was extracted from all the collected blood samples by the use of a Genomic DNA mini kit (Cat no. 1820-02; Invitrogen) in accordance with the manufacturer's protocol. DNA was quantified by Nanodrop (Eppendorf Biophotometer) and stored at −20°C before use. Amplification-refectory mutation system PCR (ARMS-PCR) for IL-18 and allele-specific PCR (AS-PCR) for TGFβ, OPN, and GALNT8 were performed by using the list of primers shown in Table 1. Both the ARMS-PCR (19) and AS-PCR (10) techniques are well established for the determination of SNPs, and both produce dependable results.
PCR amplification for IL-18 was carried out in 27 μL of PCR mixture, while for all other genes, it was carried out in 25 μL of PCR mixture comprising 100–500 ng DNA. Additionally, the PCR mixture contained 11 μL NF H2O, 200 mM dNTPs (Fermentas), 2.5 mM MgCl2 (Fermentas), 2×PCR buffer (Fermentas), and 2 units of Taq DNA polymerase (Fermentas). Cycling conditions were as follows: long denaturation at 95°C for 5 min, followed by 35 cycles of short denaturation at 95°C for 30 sec. Primers annealing of IL-18 −137 (rs1946518), OPN −442 (rs11730582), and GALNT8 (rs10849138) genetic variants was carried out at 62°C for 30 sec, while primers annealing of IL-18 −137 (rs187238), TGFβ 10 (rs1982073), and TGFβ 25 (rs1800471) genetic variants was carried out at 66°C for 30 sec. Cycling extension was performed at 72°C for 30 sec. The final extension was carried out at 72°C for 10 min. PCR amplification was run on 2% agarose gel to observe under UV light of the gel doc system (Wealtec) and compared with 50 bp DNA marker (Fermentas) as shown in Figure 1, 2 and Supplementary Figures S1–S12; Supplementary Data are available online at
Demographic representation of IL-18 −607 ARMS-PCR amplified products. Lane M represents 50 bp ladder. Lanes 1 and 2 show a heterozygous subject, while lanes 3 and 4 represent a homozygous subject with IL-18 −607 genotypes.
AS-PCR amplification of OPN. Lane M specifies 50 bp ladder. Lanes 1 and 2 specify a heterozygous subject, lanes 3 and 4 specify a homozygous subject, while lanes 5 and 6 specify a heterozygous subject with OPN −442 polymorphism.
Statistical analysis
Statistical analysis of the data was performed using SPSS v13 software (SPSS, Inc.). The Hardy–Weinberg principle was applied to all healthy subjects and patients to find the expected prevalence of different alleles and genotypes of studied cytokine genetic variants in our population. Pearson chi square analysis was performed to reveal any important variations in the prevalence of IL-18, OPN, TGFβ, and GALNT8 genotypes in treatment responders and nonresponders. A two-tailed p-value of≤0.05 was regarded as statistically significant.
Results
IL-18 gene polymorphism
The IL-18 −607 gene polymorphism was tested in 260 individuals. The demographic and clinical profile of all individuals is shown in Table 2.
Age is shown as mean±SD.
Viral load is shown as median and ranges.
SVRs, sustained virological groups; NRs, nonresponders.
Regarding IL-18 −607 gene polymorphism, there were a total of 87, 113, and 60 individuals for CC, CA, and AA genotypes respectively. All subjects were categorized into three groups: healthy controls, SVRs, and NRs. The distributions of IL-18 −607 genotypes among these groups are shown in Table 3.
p-Values were calculated by Pearson chi square test for SVR and NR groups. The boldface typed p-values represent significant values. Thus, IL-18 –607AA and OPN –442TT genotypes were positively associated with interferon-based therapy of HCV infection.
The distributions of all IL-18 −607 genotypes were in accordance with the Hardy–Weinberg principle in all three groups. The IL-18 607AA genotype was more prevalent in healthy individuals (29%) than chronic HCV patients (17.8%), signifying its possible protective function in the spontaneous clearance of HCV infection (p=0.031), as shown in Figure 3.
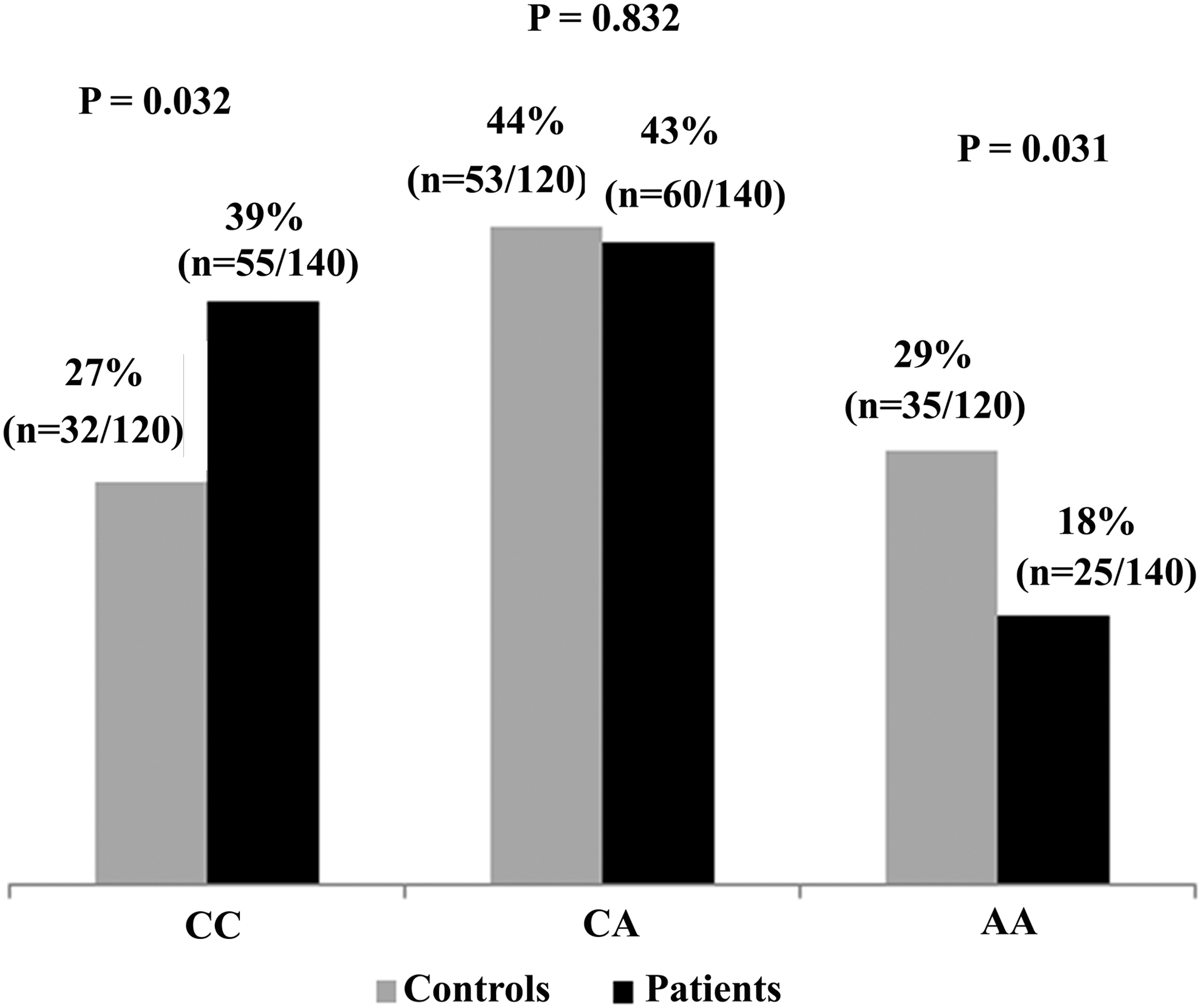
Percentage distributions and p-values of IL-18 −607 genotypes in healthy control and patients. p-Values for IL-18 −607 genotypes CC, CA, and AA were 0.032, 0.832, and 0.031, respectively.
Pearson chi square analysis of SVR and NR groups showed that there was no considerable variation between either group for the distribution of IL-18 −607CC and CA genotypes. However, the distribution of IL-18AA genotypes among the SVR group was considerably higher than the NR group (p=0.046), as shown in Figure 4.
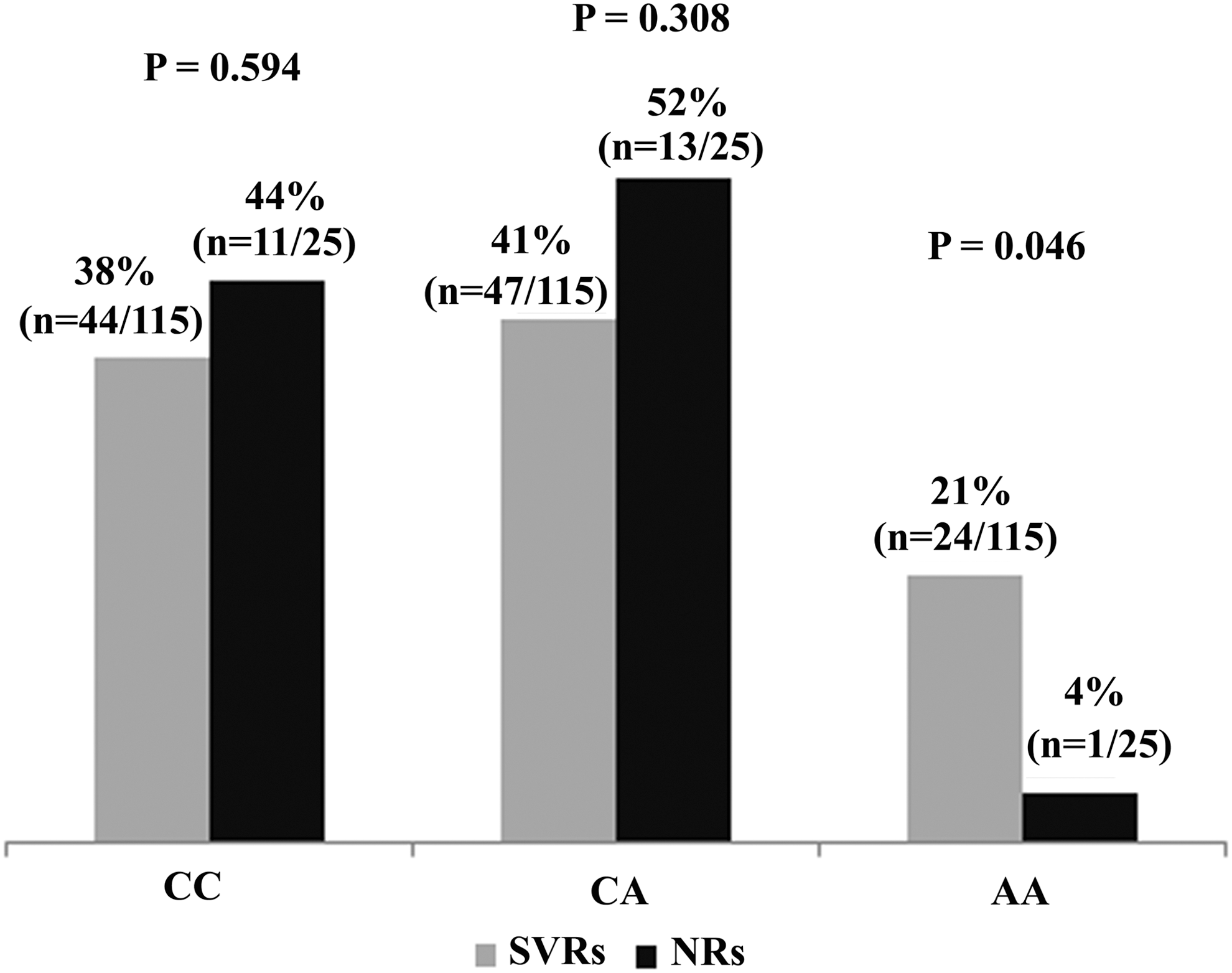
Graphic representation of IL-18 −607 genotypes distributions in SVRs and NRs. p-Values for IL-18 −607 genotypes CC, CA, and AA were 0.594, 0.308, and 0.046, respectively. SVRs, sustained virological groups; NRs, nonresponders.
Regarding IL-18 −137 genotypes, the total distributions of GG, CG, and CC genotypes were 100, 131, and 29 respectively. The prevalence of these genotypes was in agreement with the Hardy–Weinberg principle. No considerable variations in the prevalence of IL-18 −137GG, CG, or CC genotypes were noticed in any of the three groups (Table 3).
OPN −442 polymorphism
The collective prevalence of the OPN −442 polymorphism in 260 subjects was 53, 118, and 89 for CC, CT, and TT genotypes respectively (Table 3). The distributions of all OPN −442 genotypes were in accordance with the Hardy–Weinberg equilibrium. We found no considerable variations in occurrence of all OPN −442 genotypes among healthy controls and patients. When the patients were further categorized into SVR and NR groups, no significant variations were found in the prevalence of OPN −442CC and CT genotypes, as shown in Figure 5. However, the distribution of the OPN −442TT genotype was considerably higher in the SVR group than the NR group, signifying its positive correlation with treatment response to combinational therapy (p=0.034). Moreover, we found that all IL-18 −607 genotypes were inherited and independent of OPN −442 genotypes. There was no coinheritance of any genotype of IL-18 −607 with any OPN −442 genotype.

Graphic representation of OPN −442 genotypes distributions in SVRs and NRs. p-Values for OPN −442 genotypes CC, CT, and TT were 0.441, 0.155, and 0.034, respectively.
TGFβ gene polymorphism
The distributions of TGFβ codon 10 polymorphisms in 260 individuals were 80, 140, and 40 for TT, CT, and TT genotypes respectively, while the distributions of TGFβ codon 25 polymorphism were 124, 101, and 35 for genotypes GG, GC, and CC respectively (Table 3). Genotype distributions of both polymorphisms in all three groups (control, SVR, and NR) were in agreement with the Hardy–Weinberg equilibrium and showed no significant differences. Pearson chi square analysis of the SVR and NR groups showed no significant correlation of any genotype of TGFβ codon 10 or TGFβ codon 25 polymorphisms with the treatment response to HCV infection (Table 3).
GALNT8 gene polymorphism
The distributions of GALNT8 genotypes in 260 individuals showed that their collective prevalence was 63, 133, and 64 for genotypes CC, CG, and GG respectively. The distribution of this polymorphism in all groups followed the Hardy–Weinberg equilibrium, with no important variations in the prevalence of GALNT8 polymorphism in any of the three groups (control, SVR, and NR). Pearson chi square analysis of the SVR and NR groups showed no significant correlation of any GALNT8 genotype with response to interferon plus ribavirin treatment of HCV infection (Table 3).
Discussion
Our recent efforts were focused on those host factors that may significantly predict treatment response of chronic HCV-infected subjects in the Pakistani population. We considered some of the most important SNPs of IL-18, OPN, TGFβ, and GALNT8 regarding natural clearance and treatment response of HCV-infected subjects from Sind and Khyber Pakhtun khwa. We found no considerable discrepancies in the prevalence of host SNPs. The rate of treatment response in chronic HCV-infected subjects from the two studied provinces was also comparable. Therefore, we did not categorize patients on the basis of province. Our results showed that, overall, there was a high SVR rate for interferon-based treatment of HCV infection in our population (82%). Published data from this region have also shown high SVR rates for interferon-based therapy of HCV infection (4).
SNPs of host genes contribute significantly to interferon-based therapy of HCV infection. Genome-wide association studies (GWAS) suggest a significant correlation of IL-28B SNPs (rs12979860, rs12980275, and rs8099917) in spontaneous clearance and interferon plus ribavirin treatment of HCV infection (14,39). The function of IL-28B SNPs in HCV genotype 2 and 3 is still controversial. However, the function of IL-28B SNPs in HCV genotype 1 is well established. Therefore, genetic testing of IL-28B is approved only for HCV genotype 1 by the European Association for the Study of the Liver (EASL) and the American Association for the Study of Liver Diseases (AASLD) (5).
IL-18 with six exons and five introns is positioned on chromosome 11q22.2-q22.3 (27). Promoter polymorphism of IL-18 −607 and −137 may potentially have an effect on IFN-γ (50). These promoter polymorphisms have been observed as susceptible loci for various diseases such as atopic eczema (36), type 1 diabetes (26,21), and rheumatoid arthritis (17,43). Our current findings suggest that IL-18 promoter variant (-607) is also associated with treatment outcome HCV infection in the Pakistani population. Chronically infected HCV subjects possessing AA genotype for IL-18 −607 have a higher SVR rate than other subjects possessing CC or CA genotypes for interferon plus ribavirin treatment (p=0.046). A recent report from Egypt has demonstrated that the occurrence of IL-18 −607AA genotype was considerably higher in healthy controls than patients, pointing to the significant contribution of this genotype in the spontaneous clearance of HCV infection (13). A recent study from India has also demonstrated that IL-18 −607AA genotype has a protective role in HCV infection, as it is linked with a reduction in disease severity (30). In our study, we not only found a significant variation in the prevalence of the IL-18 −607AA genotype in healthy controls and patients (p=0.031), but we also found that the distribution of this genotype was very much higher in the SVR group than the NR group (p=0.046). Hence, it is can be concluded that individuals possessing the IL-18 −607AA genotype have a significantly higher SVR rate than individuals possessing the IL-18 −607CA or CC genotypes. Regarding IL-18 −137 gene polymorphism, we did not find any association with natural clearance and treatment outcome of HCV infection (Table 3 and Supplementary Figures).
OPN polymorphism at nt −442 is found to be closely associated with hepatitis activity in HCV subjects (32). Our findings suggest that HCV-infected subjects with the OPN −442TT genotype demonstrate a high SVR rate to interferon and ribavirin treatment. The incidence of the OPN −442TT genotype was significantly higher in the SVR group than the NR group (p=0.034), signifying its positive correlation with combinational therapy for HCV infection. We did not find a statistically significant correlation of any other OPN −442 genotypes in the natural clearance or treatment response of HCV infection (Table 3). A recent study has established that promoter polymorphism of OPN may be a useful marker for the treatment outcome of HCV infection (33).
Our data suggest that the occurrence of the IL-18 −607AA genotype in SVR subjects of HCV genotype 3a was two times greater than NR subjects of HCV genotype 3a (7.82% vs. 4%). On the other hand, the distribution of the OPN −442TT genotype was even higher (six times) in SVR subjects of HCV genotype 3a than NR subjects of HCV genotype 3a (25.2% and 4%). Our results reveal that the OPN treatment favorable genotype −442 TT was more frequent in HCV genotype 3a SVR subjects than the IL-18 treatment favorable −607AA genotype.
In the case of chronic HCV patients, it is demonstrated that viral core protein upregulates TGFβ transcription (46), which in turn leads to exacerbating liver fibrosis progression (25). TGFβ polymorphisms at codon 10 and codon 25 have been linked with faster progression of liver fibrosis (15,38). Functional polymorphism at codon 10 of TGFβ is likely to interrupt the export capability of newly synthesized proteins (18). It has been established that genetic variant at codon 25 of TGFβ is linked with the development of graft fibrosis after liver transplantation (12). In the current study, we searched for the correlation of TGFβ polymorphism at codon 10 and codon 25 with HCV pathogenesis and treatment response. Our findings demonstrated that there were no significant variations in the distributions of TGFβ functional variants at codon 10 or 25 in any of the three groups (control, SVR, and NR as shown in Supplementary Figures). Thus, there was no significant correlation of any TGFβ genotype with spontaneous clearance and treatment response of HCV infection in our population (Table 3).
A recent report by Nakano et al. demonstrated an association of GALNT8 polymorphism rs2286580 in intron 6 of polypeptide with interferon monotherapy of HCV infection in the Japanese population (34). However, the results were not replicated in patients of the same population receiving pegylated interferon and ribavirin treatment for HCV infection. The correlation peaked for the GALNT8 variant rs10849138 for combinational therapy of HCV infection. We studied the correlation of rs10849138 for combinational therapy of HCV infection. Our results showed no significant correlation of any genotype of rs10849138 with spontaneous clearance and response to interferon plus ribavirin treatment of HCV infection in our population (Table 3 and Supplementary Figures S1–S12).
The main limitation of our study was the small sample size, particularly when the data were evaluated on the basis of HCV genotypes or provinces. However, the sample size for HCV genotype 3a was sufficient to highlight its association with IL-18 −607AA and OPN −442TT genotypes.
Conclusion
In conclusion, interferon-based therapy of chronic HCV infection is associated with serious side effects and cost effects besides its long treatment duration. This study may help health policy makers to suggest alternative treatment options for those chronic HCV patients where the chance of failure of interferon and ribavirin treatment of HCV infection is greater. Our study shows that IL-18 AA genotype at −607 and OPN TT genotype at −442 may be used as positive predictive markers for combinational therapy of HCV infection in the Pakistani population.
Footnotes
Acknowledgments
Financial support for this study was provided by the National University of Sciences and Technology (NUST), H-12, Islamabad, Pakistan, and Higher Education Commission of Pakistan.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.