
Abstract
Tracheal mucosa is the primary site of replication of avian infectious bronchitis virus (IBV), which leads to both morphologic and immune modulatory changes in this organ. To increase the understanding of the mechanisms involved in these processes, we focused on the evaluation of local inflammatory and cell-mediated immune responses after challenge with the M41 strain of IBV, associating these responses with pathologic changes in the tracheal mucosa. At 24 h post-infection, inflammatory cytokines related genes were significantly upregulated, including peaks of TNFSF15 and TGFβ mRNA production, although no tracheal microscopic alterations were observed and only a slightly increase in viral load occurred. At 3 days post-infection (dpi), we observed that the highest upregulation of IL6, IL1β, and IFNγ coincided with highest scores of viral load and microscopic lesions, suggesting a role of both these cytokines and virus load on the development of tracheal lesions. Later, at 7 dpi, the most prominent increases of CD8αα mRNA and Granzyme homolog A mRNA were followed by a significant decrease of scores of tracheal lesions and viral load. In conclusion, an early upregulation of expression of proinflammatory cytokines such as IL6, IL1β, and IFNγ induced by the M41 strain of IBV may be partially implicated in the viral pathogenicity on trachea tissues of nonimmune challenged chickens, in addition to a late induction of a putative protective immune responses by this virus through upregulation of CD8αα and Granzyme homolog A genes in this organ.
Introduction
I
The tracheal lesions in chickens experimentally infected with a respiratory pathotype of IBV were related to the development of concomitant tracheal mucosa inflammatory cell infiltration, and their maximum intensity coincided with the peak of heterophil influx (14). The recruitment of leukocytes in some respiratory virus infections has been associated with early production of proinflammatory cytokines (29).
Additionally, chickens experimentally infected with attenuated IBV strain (H120) showed an increased expression of genes of proinflammatory cytokines in their tracheal tissues (15). The authors assumed that the expression of proinflammatory cytokines are involved in the activation of innate immune responses in the early phase of IBV infection, by recruiting polymorph and mononuclear leukocytes to the primary site of viral infection (15). To reinforce this hypothesis, recent studies indicated that the nonstructural protein 3 (nsp3) of coronaviruses, mainly the ADP-ribose 1′-phosphatase domain (ADRP), exerts a modulatory activity on innate immunity by inducing host cells to produce proinflammatory cytokines, such as IL6 and TNFα. Mutation in this genome region may lead to alterations in pathogenicity of virus by modulating host cytokine expression (12,30).
However, there are few studies about the course of local mechanisms involved during IBV pathogenesis. One report compared the abilities of two different strains of IBV to cause clinical signs, mortality, and microscopic lesions, and correlated them with gene expression of proinflammatory cytokines (IL1β, IL6, and lipopolysaccharide-induced TNF-α factor (LITAF) and viral load in the trachea and kidneys (18). Additionally, this report demonstrated that proinflammatory response showed diverse severity, depending on IBV strain, consequently leading to a variable degree of injury in the host (18).
It has been also recognized that several markers of cell-mediated immune (CMI) response, including cytotoxic CD8 T-cells, cytokines, perforins, and granzymes, are the major factors controlling the development of protective memory immune responses, as well as viral clearance to IBV infection (6,15,26,36). However, besides their protection role against IBV infection, CD8 T-cells could be involved in the damage to respiratory tract tissues, as reported for influenza virus infection (22).
The scarce information on the activities involved in the local pathogenesis of IBV in chickens motivate us to investigate the inflammatory and cell-mediated responses during the development and repair of IB infection, correlating them with viral load profile and with the course of pathologic changes induced at tracheal mucosa level.
Materials and Methods
Virus
The M41 strain of IBV (Brazilian Agricultural Research Corporation—Embrapa Swine and Poultry) was propagated and titrated in 10-day-old specific pathogen-free (SPF) embryonated chicken eggs. The eggs were inoculated via the allantoic sac route, as described previously (27). The virus was titrated in embryonated chicken eggs according to standard procedures, and the 50% embryo infectious doses (EID50) were determined (31).
Experimental design
All procedures have been approved by the Veterinary Sciences Animal Care Committee of Universidade Estadual Paulista, according with ethical principles of animal experimentation adopted by Brazilian College of Experimentation.
Two groups of 25 one-day-old SPF chicks (White Leghorn lineage) were housed in positive pressure isolators. At 21 days of age, one group was experimentally infected via intraocular and intranasal routes with 104.0 EID50/bird of IBV M41 strain. A negative control group was mock infected with SPF allantoic fluid and maintained under the same conditions. Five chickens per interval from each group were sacrificed at 8 h post-infection (hpi), 24 hpi, 3 days post-infection, 7 dpi, and 14 dpi. Tracheal samples were collected from each group. A portion was immediately frozen and kept at −70°C until processing, and the remaining portion was subjected to histopathologic and ciliostasis analysis.
Scoring of tracheal lesions
The tracheal samples were divided in three portions: proximal, medial, and distal. Each portion was prepared for ciliary kinetic analysis and histopathology, and the scores were determined as previously described (2,8,26,38).
Briefly, for ciliary kinetic analysis, the scores ranged from 0 to 4 according to the percentage of ciliary activity: 0=100% of ciliary movement, 1=75–100% of ciliary movement, 2=50–75% of ciliary movement, 3=25–50% of ciliary movement, and 4=0–25% of ciliary movement. For histopathologic analysis, the scores ranged from 0 to 3 according with the severity of the observed lesions. Absence of injury was classified as 0, while mild, moderate, and severe were classified as 1, 2, and 3 respectively. Morphologic characteristics were observed for loss of cilia and epithelial cells, degeneration of glands, inflammatory infiltrate at mucosa, inflammatory infiltrate at adventitia, and epithelial hyperplasia.
Real-time quantitative reverse transcription polymerase chain reaction
The RNA extractions from the proximal third from tracheal samples of experimentally infected chickens were performed using the Trizol Reagent (Invitrogen). The residual genomic DNA was digested by DNase treatment of RNA samples using a RNase-free DNase I kit (Fermentas). The RNA quality was analyzed on a 1% gel and quantified by ultraviolet (UV) absorbance at 260 nm (A260). The cDNAs was synthesised according to instructions provided with Reverse Transcriptase (Fermentas) and using OligodT primers (IDT), with the exception that dithiothreitol (DTT) was omitted from the reaction (26).
Real-time quantitative reverse transcription polymerase chain reaction (RT-qPCR) using the SYBR Green I marker was used for the relative and absolute quantification of mRNA and viral load respectively. The reaction mixture contained 2.0 μL of cDNA, 10.0 μL of Maxima SYBR Green qPCR Master Mix (Fermentas), and 10 pmol of each primer (Table 1) in a final volume of 20 μL. The amplification reaction included a preincubation step at 95°C for 10 min to activate the HotStart Taq DNA polymerase. This reaction was followed by 40 cycles of amplification (30 cycles for the 18S), including denaturation at 95°C for 30 sec, annealing at 49.2–65°C (see Table 1) for 30 sec, and extension at 72°C for 30 sec. Following amplification, a melting curve analysis was performed by raising the incubation temperature from 65°C to 95°C in 0.2°C increments with a hold of 1 sec at each increment. The oligonucleotides used were designed according to sequences deposited in GenBank and using Primer3 software (frodo.wi.mit.edu), with the exception of the IFNγ oligonucleotides, which were designed according to previous descriptions (see references in Table 1). The annealing temperatures were selected based on the highest sensitivity (lowest Cq value) and specificity (absence of nonspecific products) provided by the gradient of temperatures tested.
Bp, base pairs; F, forward; R, reverse; a, 5′ to 3′ sense.
The relative expression of the IL1β, IL6, IFNγ, TNFSF15, TGFβ, CD8αα, and Granzyme homolog A genes in the tracheal samples of infected chicks was quantified as the fold change relative to the noninfected group (Control) at the same time point, and the gene expression from each sample were standardized using the Cq value of the 18S rRNA for the same sample (20).
All tracheal samples were tested for viral replication using the HV+ and HV− primers (for S1 gene of IBV), and the Cq results were used to calculate the log of the number of moles using the linear equation from standard curve, optimized previously (26).
Statistical analysis
The comparisons of the mean relative changes in gene expression, and tracheal pathologic changes (viral replication, histopathology, and ciliostasis) between the experimental groups were performed using the Kruskal–Wallis test followed by Dunn's test. Correlations between the immune response parameters and the tracheal pathology parameters were estimated using the Spearman method. All analyses were conducted using GraphPad software (version 4.0), and the level of significance was set at p<0.05.
Results
Course of viremia and pathologic alterations of the trachea induced by IBV challenge
No pathologic alterations of trachea (ciliostasis and histopathology) were observed in the uninfected group for all post-infection intervals tested (Figs. 1 and 2A). In addition, no positive sample for S1 gene was detected by RT-qPCR in the samples from control group.
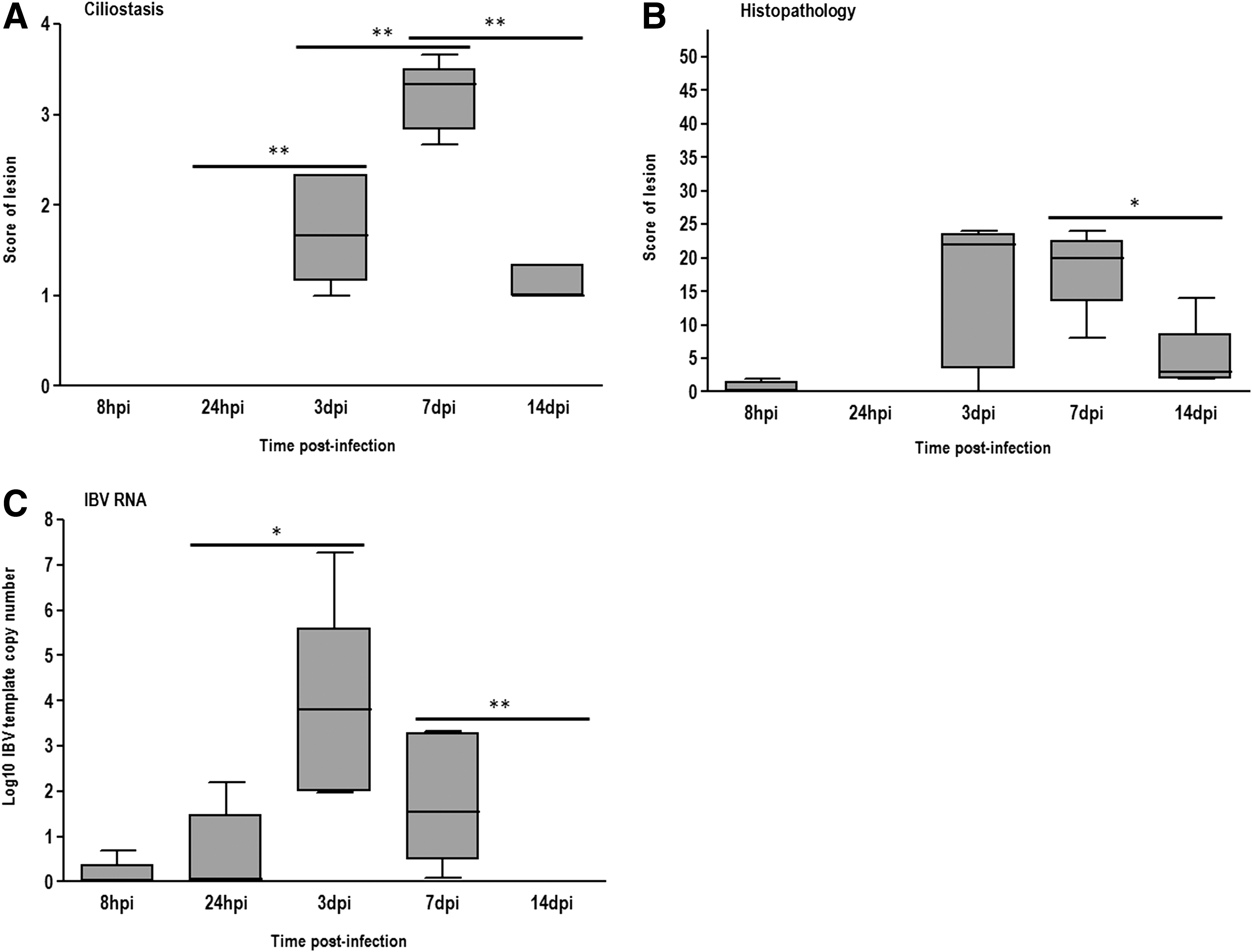
Pathologic alterations of the trachea induced by infectious bronchitis virus (IBV) challenge in samples collected at 8 h post-infection (hpi), 24 hpi, 3 days post-infection (dpi), 7 dpi, and 14 dpi from chickens experimentally challenged at 21 days of age with the M41 strain of IBV. **p≤0.005; *p≤0.01.
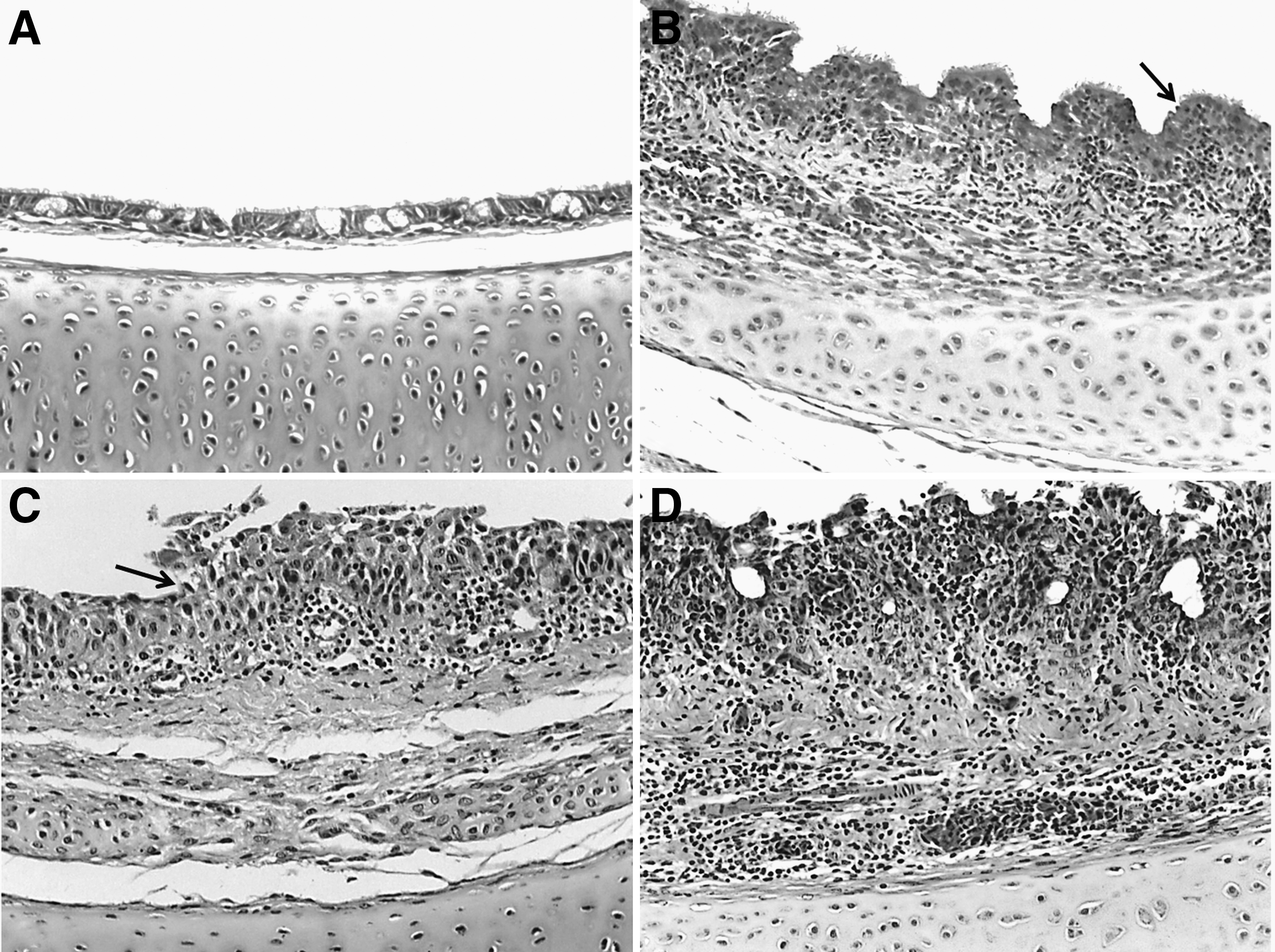
Microscopic lesions (200×) observed in tracheal samples collected from chickens experimentally infected with the M41 IBV strain or mock infected, at different time points after challenge.
The IBV-challenged group had no significant changes of ciliostasis and microscopic lesions in their tracheal samples at 8 hpi and 24 hpi. However, the copy numbers of S1 gene showed a slight increase (0.56 and 1.62 median Log10 of IBV copies for 8 hpi and 24 hpi respectively). At 3 dpi, all the pathologic scores were markedly increased, and they were characterized by the detection of the most prominent changes in histopathology and viral replication (4.96 median Log10 IBV copies), though ciliostasis peaked at 7 dpi (Figs. 1 and 2B–D). The severity of lesions decreased significantly (p≤0.05) at 14 dpi and nearly reached the basal levels. A similar trend was found for viral load (0.09 median Log10 IBV copies; Figs. 1 and 2). Additionally, significant positive correlations were observed between viral load and the scores of tracheal histopathology, as well as with ciliostasis scores (Table 2).
GraA, granzyme homolog A; C, ciliostasis scores; H, histopathology scores; VL, viral load—absolute quantification of S1 gene of IBV; NS, no significant correlation; *** p<0.001; ** p<0.005; * p<0.05.
Inflammatory related innate immune responses
All proinflammatory transcripts evaluated in this study were significantly upregulated in the tracheal samples at 24 hpi. IL1β reached higher levels of expression at 3 dpi, showing approximately fourfold higher levels than those recorded for birds from the negative control group (Fig. 3B). The expression of IL6 showed a similar trend of increase, except that higher levels of expression were observed, for example an approximately 16-fold increase compared to the negative control samples (Fig. 2A). TNFSF15 and anti-inflammatory cytokine (TGFβ) presented a slightly significant increase in their expression and peaked at 24 hpi (Fig. 3C and D). The levels of expression of IL1β and IL6, after peaking at 3 dpi, declined up to 14 dpi, while the levels of expression of TNFSF15 and TGFβ, after peaking at 24 hpi, declined in the two subsequent time points (3 and 7 dpi), but increased slightly at 14 dpi.
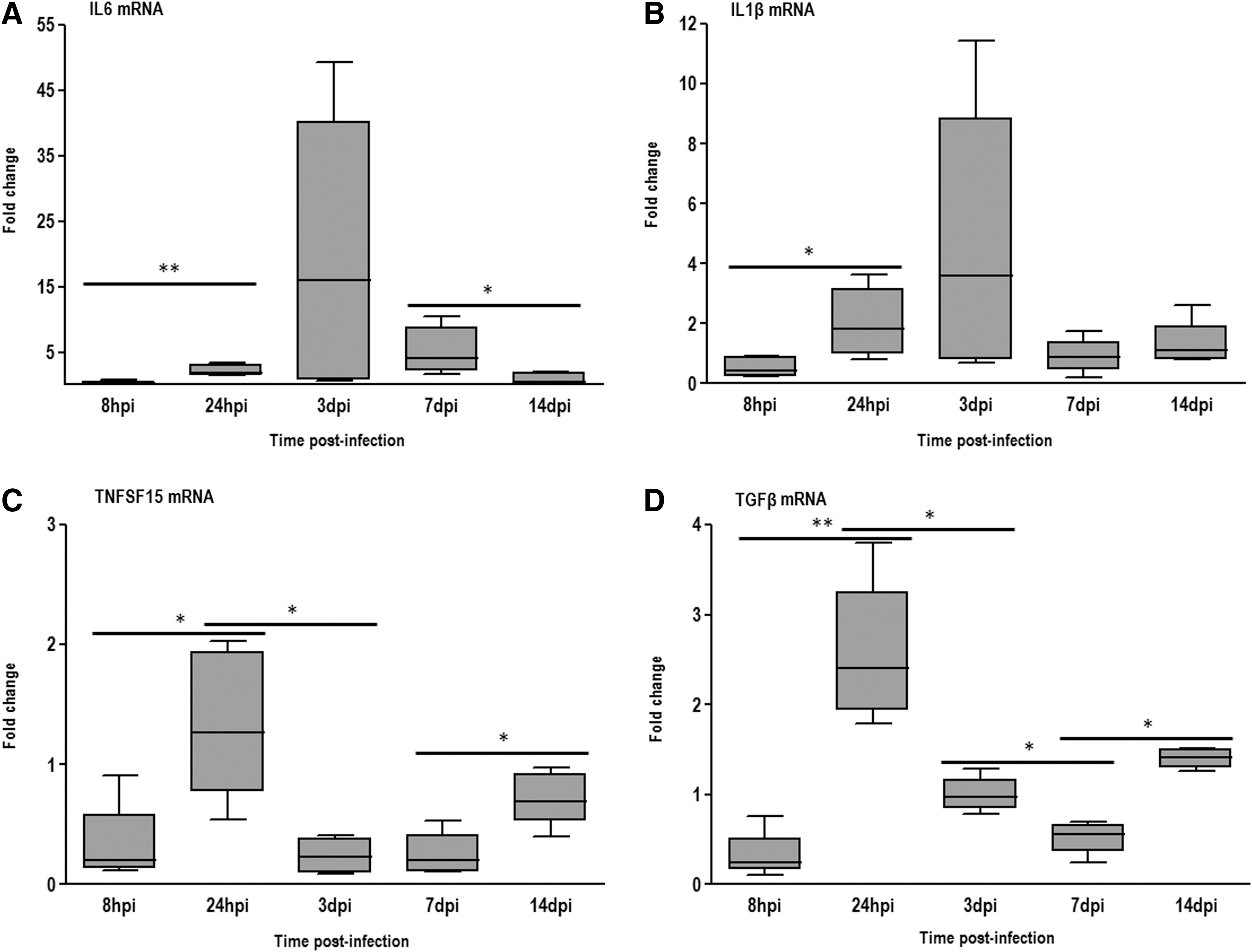
Fold changes mRNA expression of inflammatory related genes from innate response
The increased IL6 mRNA levels were positively correlated with viral load in the trachea and ciliostasis scores, while no correlation was observed for IL1β expression levels with pathologic alterations in the trachea. Quantification of TNFSF15 transcripts was negatively correlated with ciliostasis and histopathologic scores. No significant correlation was observed between TGFβ transcripts and pathologic changes in the trachea (Table 2).
CMI response
Significant increases were observed for the expression of Granzyme homolog A and IFNγ at 24 hpi. The highest levels of IFNγ transcripts were observed at 3 dpi, while CD8αα and Granzyme homolog A peaked at 7 dpi (Fig. 4).
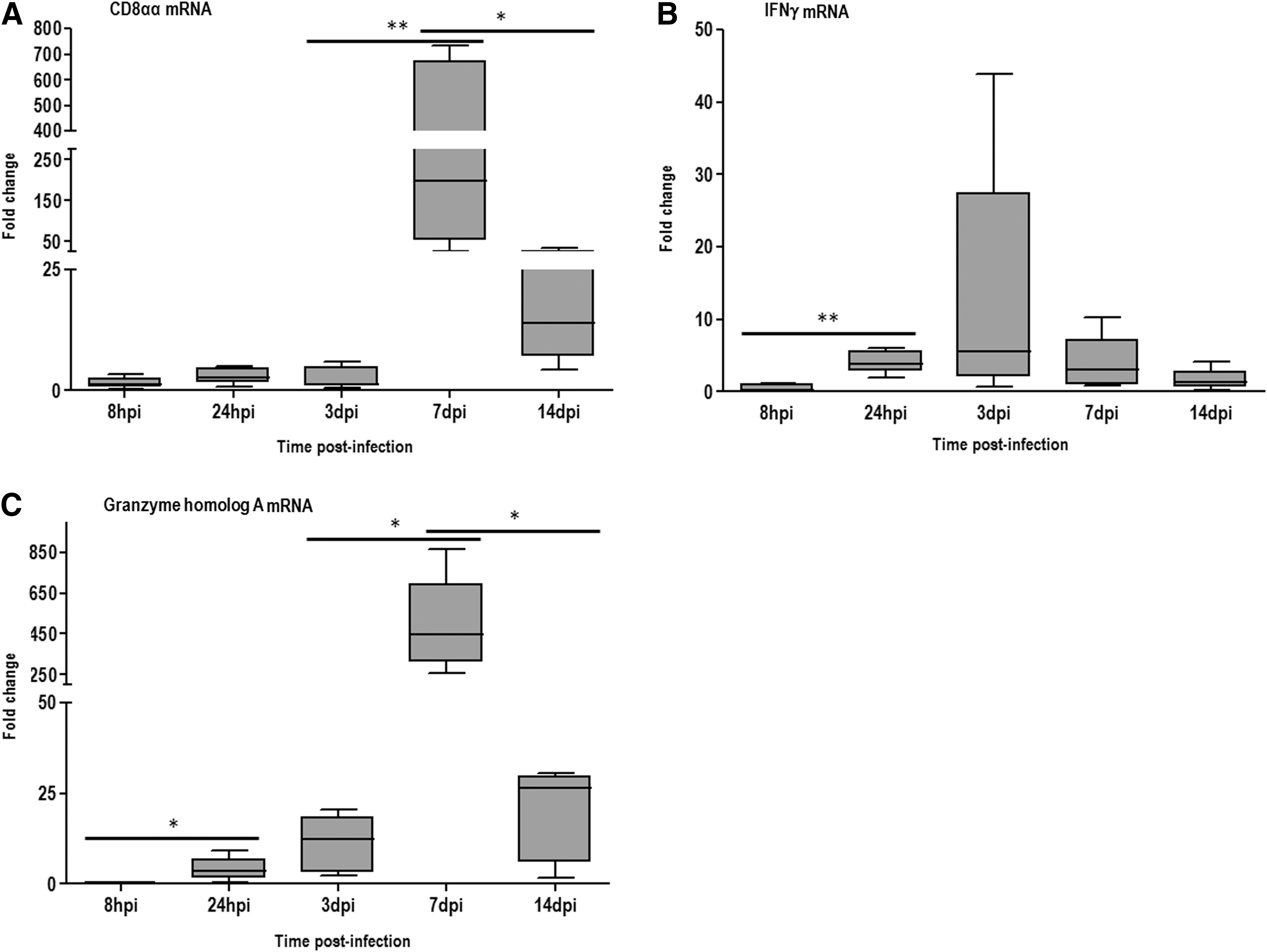
Fold changes mRNA expression of cell-mediated related genes
Significantly positive correlations were found for the expression of CD8αα and Granzyme homolog A and pathologic changes in the trachea (ciliostasis and histopathology) were found, including a correlation coefficient of 0.8051 for Granzyme homolog A expression and ciliostasis scores (Table 2).
Discussion
IBV causes an acute and highly contagious disease in chickens. Tracheal mucosa is the primary site of viral replication. However, the local inflammatory responses and the immune mechanisms involved in the development and repair of tracheal lesions occasioned by IBV, particularly regarding to the kinetics, magnitude, and duration, are still lacking.
At the early phase of IBV infection, from 8 hpi to 24 hpi, no relevant pathologic changes in the trachea coincided with significant increases of all transcripts of pro- and anti-inflammatory responses (IL6, IL1β, TNFSF15, and TGFβ) or of Granzyme homolog A and IFNγ. These increases in the expression of pro- and anti-inflammatory cytokines could be derived from the activation of macrophages and epithelial cells present in the trachea, while increased expression of Granzyme homolog A and IFNγ was probably produced by NK cells and not by CD8 T-cells, at this time point, as described in the respiratory tract for influenza virus, IBV, and SARS coronavirus infections (25,33,36).
Gene expression of TNFS15, a functional homolog of TNFα (28,35) involved in chicken inflammatory responses, increased mainly at the early phase of IBV infection, and demonstrated a significant negative correlation with ciliostasis and histopathologic score, for example the higher the levels of TNFSF15, the lower were the numbers of lesions observed. These results indicate that this proinflammatory cytokine could be involved in inflammatory responses that contribute to the reduction of both viral infection and tracheal pathologic alterations, though further investigations must be conducted to verify this hypothesis. In this context, although the “cytokine storm” after a number of viral infections is commonly associated with morbidity and contribution to the severity of pathologic changes in the host, a recent study (5) indicated a role of local immune responses acting to limit the SARS-CoV replication in experimentally infected primates. Besides, inhibition of TNFα response in mice inoculated with a high pathogenic strain of H5N1 avian influenza virus did not reduce the morbidity and mortality commonly caused by this viral infection (32).
During the acute phase of IBV infection, particularly at 3 dpi, the highest values of histopathology scores and viral load were found, and they were associated with peaks of the expression of inflammatory cytokines, such as IL6 and IL1β, and higher IFNγ mRNA expression values were also observed. This suggests a role of both these cytokines and virus load on the development of tracheal lesions. Similar findings were reported after experimental infection with two other genotypes and pathotypes of IBV in both trachea and kidney samples (18). In that study, the highest production of local inflammatory cytokines (IL6, IL1β, and LIFAT) mRNA has been demonstrated to be associated with peaks of viral load in these organs. In our study, although no significant correlations were observed for IL1β against pathologic alterations and/or viral load, the profile of transcription of this cytokine was close similar to IL6 proinflammatory cytokine tested in this experiment.
In our experiment, the level of IL6 transcripts increased sharply at 3 dpi and showed positive correlations with ciliostasis scores and viral load. Although no significant correlation was observed between IL6 mRNA and microscopic lesions, similar profiles of increasing and decreasing of these parameters were observed for both methods, except at the 7 dpi interval when the transcripts of IL6 mRNA returned to expression levels near to the basal values while the scores of tissue damage remained high. A similar marked increase in IL6 expression was previously observed in kidney tissues from birds experimentally infected with nephropathogenic IBV strain (7). In another study, IL6 transcripts were also directly related to renal lesions in chickens challenged with a nephropathogenic strain of IBV, mainly in IBV more susceptible S-line of chickens, compared to a more resistant HWL line of chickens (3). IL6 was also previously associated with the amount of viral RNA expression and extension of clinical illness in birds infected with NDV (11).
Moreover, in a study of protection induced by vaccine against Marek's disease, an upregulation of IL6 expression was detected after challenge in nonvaccinated group compared with the vaccinated group (1). Similar overexpression of cytokine genes was observed in chickens experimentally infected with the strain H9N2 of influenza virus, and as viral titers increased in the trachea and lung, and TNF and IFNγ genes showed a marked increase in their expression (23). TNFSF15 has been also shown to be associated with severity of inflammation caused by primary infection with Eimeria (28). More closely related to IBV infection, patients affected by severe acute respiratory syndrome (SARS) coronavirus infection exhibited significant upregulation in the expression of IL6 and IFNγ, and the levels of expression of these cytokines were highly correlated with severity of acute infection (25). IL6 is a multifunctional cytokine and can cause tissue pathology by recruiting inflammatory cells and promoting monocyte differentiation via various signaling pathways. Although we do not have direct evidence to show that IL6 damages tracheal epithelial cells, studies have shown that upregulation of IL6 plays an important role in mediating endothelial cell dysfunction through reactive oxygen species production (32,37). Overall, we believe that both viral load and inflammatory cytokines such as IL6 contribute to the local pathologic changes induced to tracheal tissues by IBV infection.
The most prominent scores of ciliostasis were found at 7 dpi, although this finding may be at least in part related to the absence of ciliary epithelia at this time point, when transcripts of CMI responses presented the highest expression (means of 200-fold for the expression of CD8αα and 450-fold for the expression of Granzyme homolog A). This upregulation in the transcription of CMI genes was followed by a significant decrease of scores of tracheal lesions and viral load, suggesting this response seems to be more related to viral clearance and reduction of lesions than to the development of tracheal lesions. Thus, the high correlations found between tracheal pathologic changes and the increased expression levels of CD8αα and Granzyme homolog A seem to point to a possible role of CMI responses in the resolution of tracheal pathologic alterations, rather than to support an immunopathologic activity for CMI responses leading to tracheal lesions. This inference was consistent with previous findings on the role of CTLs in IBV clearance (6,19,36). Moreover, early increases in the expression of CMI genes, such as IFNγ, CD8αα, and Granzyme homolog A, were related to protective memory immune response provided by vaccination with attenuated IBV strain, in a dose dependent manner (26).
In the recovery phase of IBV infection (at 14 dpi), the mucosal epithelia of trachea and ciliary movement returned to the normality, and the scores of ciliostasis and histopathology were indistinct from those observed in the birds from the noninfected group. Additionally, <1 copy of IBV (0.09 Log10 mean copy number of S1 gene) was detected by RT-qPCR at this time point. The levels of expression of IL6, IL1β, and IFNγ resumed to basal values at this interval, while others cytokine and CMI genes (TNFSF15, TGFβ, CD8αα, and Granzyme homolog A) decreased from 7 dpi, though they remained higher than their basal levels.
In summary, our results indicates that an early upregulation of expression of proinflammatory cytokines is induced by M41 strain of IBV, such as IL6 and IL1β, which seem to be partially implicated in the tracheal pathology caused by this virus in nonimmune challenged chickens. In addition, a late induction of putative protective immune responses was elicited by this virus through upregulation of CD8αα and Granzyme homolog A genes.
Footnotes
Acknowledgments
We would like to thank FAPESP (07/55070-2) and CNPq (476120/2007-1) for the financial support, Dr. Liana Brentano from Swine and Poultry, Zoetis, and M.S.D. for providing the SPF eggs and SPF chicks.
Author Disclosure Statement
No competing financial interests exist.