
Abstract
Although current polyvalent vaccines can prevent development of cervical cancer, they cannot be used to treat patients who already have the disease. Adenovirus expressing calreticulin-E7 (Ad-CRT-E7) has shown promising results in the cervical cancer murine model. We also demonstrated that immunization with Lactococcus lactis encoding HPV-16 E7 (Ll-E7) anchored to its surface induces significant HPV-16 E7-specific immune response. Here, we assessed the combination of both approaches in the treatment of a cervical cancer animal model. Intranasal preimmunization of Ll-E7, followed by a single Ad-CRT/E7 application, induced ∼80% of tumor suppression in comparison with controls. Mice treated with a combination of Ll-E7 and Ad-CRT/E7 resulted in a 70% survival rate 300 days post-treatment, whereas 100% of the mice in the control groups died by 50 days. Significant CD8+ cytotoxic T-lymphocytes infiltration was detected in the tumors of mice treated with Ll-E7+Ad-CRT/E7. Tumors with regression showed a greater number of positive cells for in situ TUNEL staining than controls. Our results suggest that preimmunization with Ll-E7 enhances the Ad-CRT/E7-mediated antitumor effect. This treatment provides an enormous advantage over repeated applications of Ad-CRT/E7 by maintaining the effectiveness of the three-dose application of Ad-CRT/E7, but avoiding the high systemic toxicities associated with such repeat treatments.
C
Recently, a quadrivalent vaccine against HPV types 6, 11, 16, and 18 has been introduced to the market (6). Although this vaccine is preventative, it does not offer an effective treatment for women who already present with the disease. For these women, a therapeutic approach is essential. Several researchers have developed therapeutic vaccines targeting E6 and/or E7 oncoproteins with promising results (7 –10).
In the past two decades, a successful strategy of therapeutic vaccination against HPV-16 has been developed, which consists of the linkage of antigens with polypeptides involved in intracellular trafficking. One of these polypeptides is calreticulin (CRT), an abundant 46 kDa Ca2+-binding protein located in the endoplasmic reticulum (ER) with two main functions: to regulate the homeostasis of Ca2+ and to act like a chaperone (11,12). The protein has been shown to associate with peptides delivered into the ER by transporters associated with antigen processing (TAP-1 and TAP-2) and with MHC class I-β2 microglobulin molecules to aid in antigen presentation (13,14). The fusion of calreticulin with HPV-16 E7 can stimulate CD8+ cytotoxic T-lymphocytes, resulting in a strong antitumor immune response. This approach has been tested in several vectors such as DNA vaccine, vaccinia virus, and adenovirus with promising results (12,15 –22).
Among the microorganisms used as vectors is the nonpathogenic lactic acid bacteria Lactococcus lactis. This noninvasive Gram-positive bacterium has been studied extensively and manipulated for the production of heterologous proteins (23 –26). Its fast, low-cost, simple production makes it an ideal tool for the formulation of vaccines that can be used in the general population.
We have previously expressed the HPV-16 E7 antigen anchored to the L. lactis surface (Ll-E7) (27). Intranasal administration of Ll-E7 in C57BL/6 mice resulted in HPV-16 E7-specific immune response (27,28).
In this study, we investigated, for the first time, whether preimmunization of C57BL/6 mice with Ll-E7 enhances the Ad-CRT/E7-mediated antitumor response.
Previously, we found that three intratumoral injections of Ad-CRT-E7 caused a strong tumor suppression that resulted in 100% survival (22). To determine whether immunization with Ll-E7, followed by intratumoral injection of Ad-CRT/E7, enhances Ad-CRT-E7-mediated antitumor effect, the antitumor effect of a single dose of Ad-CRT-E7 was assessed. Female C57BL-6 mice aged 6–8 weeks were obtained from a commercial vendor (Harlan Laboratories, Indianapolis, IN). Mice were injected subcutaneously (s.c.) with 1×105 TC-1 cells in the right leg and monitored three times per week as described previously (22). When tumor size reached ∼6 mm diameter, mice were given an intratumoral (i.t.) injection of Ad-E7 (n=13), Ad-CRT-E7 (n=13), Ad-LacZ (n=10) as described previously (22), or phosphate-buffered saline (PBS; n=10) at a final concentration of 5×1010 VP in 20 μL of PBS. Tumors were measured every 7 days, and tumor volume was determined by externally measuring each in two dimensions with a caliper. Volume (V) was determined by the equation V=(L×W 2)/2, where L is length and W is width of the tumor. It was found that Ad-CRT-E7 injection reduced the tumor size by ∼60% by day 21, which was statistically significant when compared to the PBS control group (p<0.05). In contrast, tumor size reduction induced by Ad-LacZ or Ad-E7 was much slower compared with Ad-CRT-E7 (Fig. 1A).
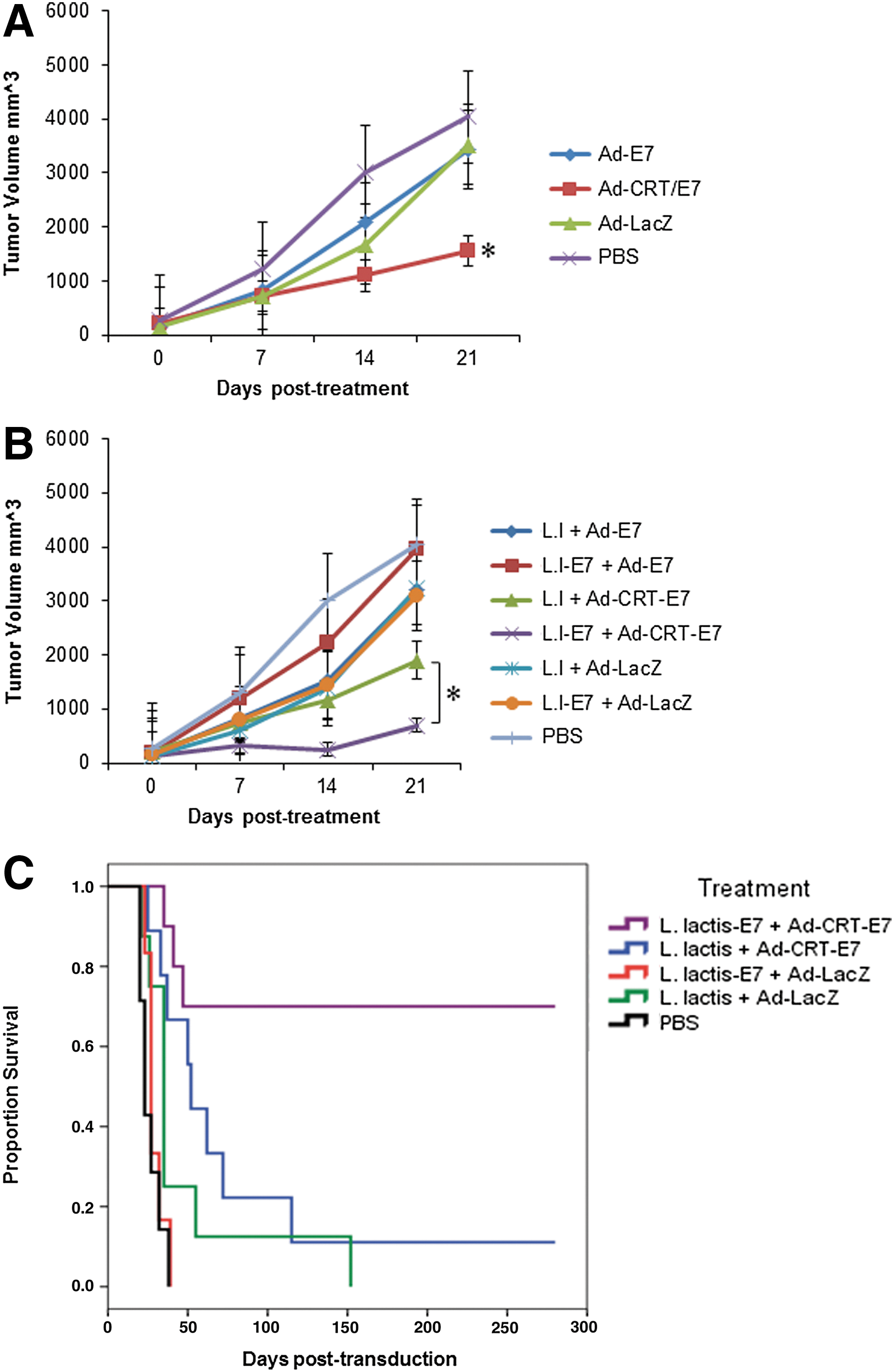
Antitumor effect of Ad-CRT-E7 alone or in combination with Ll-E7 preimmunization. (
We further investigated whether the Ad-CRT-E7 antitumor effect could be enhanced by preimmunization with Ll-E7. To this purpose, mice were immunized intranasally with L. lactis (Ll) NZ9000 or Ll-E7 as described previously (27). Ll was grown in M17 medium (Becton, Dickinson and Company, Franklin Lakes, NJ) supplemented with 1% glucose (Sigma-Aldrich, St. Louis, MO) at 30°C without agitation. The Ll-E7 strain was grown in the GM17 medium plus chloramphenicol (CM; 10 μg/mL) and Nisin (10 ng/mL; Sigma-Aldrich) to start the production of the recombinant protein. A 10 μL concentrated dose of either Ll or Ll-E7 at a concentration of 1×109 colony formation unit (CFU) was administered on days 0, 14, and 28. One week after the last dose, mice were challenged s.c. with 1×105 TC-1 cells in 50 μL into the right leg. Mice were monitored three times per week. When the tumors reached 6 mm in diameter, the mice were injected i.t. with Ad-CRT-E7 (n=13), Ad-LacZ (n=7), or PBS (n=7) at a final concentration of 5×1010 VP in 20 μL of PBS. The tumor size was determined as described above. Survival was monitored until the day the mouse died or it was euthanized. If there were no tumors, the mice were monitored for 300 days. Tumor growth rate was constant, as was expected in the group of mice treated with PBS. There was a similar tumor growth rate observed in the groups treated with Ll+Ad-LacZ or Ll-E7+Ad-LacZ. The Ll+Ad-CRT-E7 treatment decreased tumor size by ∼47% in comparison with Ll+Ad-E7. The treatment with Ll-E7+Ad-CRT-E7 decreased tumor size by ∼80% compared with Ll-E7+Ad-E7. Most importantly, Ll-E7+Ad-CRT-E7 treatment reduced tumor size by ∼60% in comparison with Ll+Ad-CRT-E7. This tumor suppression was statistically significant (p<0.05; Fig. 1B). None of the mice treated with Ll-E7+Ad-LacZ, Ll+Ad-LacZ, or PBS survived past 150 days after the adenovirus injections. The percentage survival for the group of mice treated with L. lactis control+Ad-CRT-E7 was <20% (Fig. 1C). In contrast, a 70% survival was measured in the group treated with Ll-E7+Ad-CRT-E7. These mice survived for at least an additional 250 days after the 300 days of initial survival monitoring; no signs of tumor were found at the time the experiment was stopped (Fig. 1C). These data suggest that preimmunization with Ll-E7 enhances the Ad-CRT-E7 antitumor effect. Animal experiments were done in accordance with institutional guidelines and were approved by the Universidad Autónoma de Nuevo León Institutional Animal Care and Use Committee.
In order to determine whether CD8+ cytotoxic T-lymphocytes (CTL) had played a role in the tumor suppression found in our results, mice receiving Ll-E7+Ad-LacZ (without tumor regression) or Ll-E7+Ad-CRT/E7 (with tumor regression) were euthanized and subjected to histopathologic analysis. The tumors were removed and immersed in 4% paraformaldehyde and embedded in paraffin. For the morphological studies, sections 6 μm thick were stained with hematoxylin and eosin. Tumors without regression (Ll-E7+Ad-LacZ) showed a uniformity of cells transformed and a remarkable vascularity. A representative microphotograph is shown in Figure 2A. In contrast, tumors with regression (Ll-E7+Ad-CRT-E7) were deficient in vascularity. A representative microphotograph is shown in Figure 2B.

Tumor histopathologic analysis, immunofluorescence, TUNEL assay and IFN-γ quantification. The tumor sections from mice treated with Ll-E7+Ad-LacZ (
The presence of CD8+ CTL in the tumors was detected by incubation with a specific anti-mouse CD8-FITC (Milteny Biotech, San Diego, CA). Briefly, deparaffinized tumor sections were blocked with 1% horse serum in PBS for 1 h and washed with PBS. The antibody dilution (1:200) was added to the sections and incubated for 2 h at room temperature. They were then washed and mounted with vectashield (Vector) and analyzed in a fluorescence confocal microscope (Leica TCS SP5). The immunofluorescence analysis revealed negative CD8+ T-lymphocyte-staining in tumors without regression (Fig. 2C), whereas a greater amount of positive CD8+ T-lymphocyte-staining was seen in tumors with regression (Fig. 2D). This result suggests that Ll-E7+Ad-CRT-E7-mediated tumor suppression was, at least in part, due to the antitumor activity of CTL.
The CD8+ CTL have both perforin-based and Fas ligand (FasL)-based lytic activities that result in the activation of caspase and apoptosis (29). Apoptosis is associated with internucleosomal degradation of genomic DNA, which can be assessed by in situ terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay (30). Briefly, paraffin-embedded tissue sections were processed for the TUNEL assay, which was done according to the manufacturer's instructions (Trevigen, Gaithersburg, MD). The tumor sections of mice treated with Ll-E7+Ad-LacZ (without tumor regression) did not show significant evidence of internucleosomal DNA fragmentation (Fig. 2E). In contrast, the tumor sections of mice treated with Ll-E7+Ad-CRT-E7 (with tumor regression) showed a greater number of positive cells for in situ TUNEL staining (Fig. 2F). These data indicate that Ll-E7+Ad-CRT-E7-mediated tumor suppression was, at least in part, caused by apoptosis.
In addition to infiltration of CTLs into the tumor, IFN-γ production was analyzed, which is a signature cytokine for Th1 responses and an effector function of CD8+ T-cells that are critical to tumor eradication (22). Splenocytes (1×107) from different treatments were co-cultivated with mitomycin-C-treated TC-1 cells (1×106) in 6-well plates for 3 days. The supernatants of the culture were collected, and levels of IFN-γ were determined using an IFN-γ enzyme-linked immunosorbent assay (ELISA) kit (R&D systems, Minneapolis, MN), according to the manufacturer's protocol. The concentration of IFN-γ in the group without tumor regression treated with Ll+Ad-LacZ or Ll-E7+Ad-LacZ increased slightly in comparison to mitomycin-C-treated TC-1 cells alone. In contrast, IFN-γ concentration was significantly greater in the Ll-E7+Ad-CRT-E7 treatment, which showed tumor regression (p<0.05; Fig. 2G). Taken together, these results show that the tumor suppression was mediated, at least in part, by the cellular immune response.
The results of the in vitro assays and tumor growth in mice from treatment groups were analyzed by the unpaired Student's t-test using a one-way ordinary parametric analysis of variance. Statistical significance was set at p<0.05.
Here, we report, for the first time, that preimmunization with L. lactis expressing E7 anchored to its surface in combination with single dose of Ad-CRT-E7 has similar antitumor immune response to three doses of Ad-CRT-E7. We suggest that this protocol of immunization has great potential to be translated to clinical settings given its enormous advantage over repeated applications of adenovirus. In this way, adenoviral-mediated hepatic and other potential toxicity could be greatly reduced. At this point, we do not know the mechanism by which preimmunization with Ll-E7 enhances Ad-CRT-E7-mediated antitumor activity. However, previously, we found that nasal immunization with Ll-E7 caused the production of E7-specific antibodies and IFN-γ (27). Therefore, it is possible that an antibody-dependent cell-mediated cytotoxicity and/or IFN-γ-mediated immunoregulatory and antitumor activities could play a role.
Footnotes
Acknowledgments
This work was supported by award numbers National Council of Science and Technology (CONACYT 47478-M) and (IMSS NO. 2005-1/I/196). BRRG received a PhD scholarship from CONACYT of Mexico. The authors thank Margaret Abby for editing.
Author Disclosure Statement
No competing financial interests exist.