
Abstract
As a prelude to immunization studies in nonhuman primates, we compared in mice the immunogenicity of a simian immunodeficiency virus (SIV)-based integrase (IN)-defective lentiviral vector (IDLV) encoding the model antigen-enhanced green fluorescence protein (eGFP) in the presence or absence of the murine granulocyte–macrophage colony-stimulating factor (mGM-CSF) expressed from an internal ribosomal entry site (IRES) sequence. BALB/c mice were immunized once intramuscularly with IDLV expressing eGFP alone or eGFP and mGM-CSF and immune responses were evaluated up to 90 days from the single intramuscular immunization. Results indicated that the mGM-CSF was unable to improve the magnitude and quality of the immune response against the eGFP transgene in the context of the SIV-based IDLV, as evaluated by enzyme-linked immunosorbent spot (ELISPOT) assays for interferon-γ (IFN-γ) and by intracellular cytokine staining for IFN-γ, interleukin-2 (IL-2), and tumor necrosis factor-alpha (TNF-α). These findings suggest that for vaccination purposes, the presence of mGM-CSF expressed after the IRES in a SIV-based IDLV system does not favor the improvement of the immunological response against the transgene of interest. Further studies should investigate whether the selection of a different cytokine gene might improve the immune response against the transgene.
Introduction
V
Concerning the use of a cytokine adjuvant, we showed that a single intramuscular inoculum in mice of IDLV expressing the HIV-Env and murine granulocyte–macrophage colony-stimulating factor (mGM-CSF) elicited long-lasting and Env-specific polyfunctional CD8+ T EM cells as well as anti-Env Abs (29,30,32). However, in these reports, we did not perform a comparative evaluation in the absence of mGM-CSF. In this context, a recent report showed that IDLV encoding cytokines, including human GM-CSF, induced differentiation of ex vivo transduced human DCs, which were able to stimulate multivalent immune responses after injection in immunodeficient NOD/Rag1 (NGR) mice (8). GM-CSF is a cytokine responsible for recruitment, activation, and maturation of antigen-presenting cells (APC) (26). Coinjection with plasmid DNA expressing GM-CSF has been reported to increase immunity against immunogens from herpes simplex virus (33), classical swine fever virus (1), foot-and-mouth disease virus (9), HIV-1 (37), and infectious bronchitis virus (43), resulting in the enhancement of antigen-specific humoral and cellular immune responses. In contrast, some studies have suggested that GM-CSF can reduce immune responses (20,40,47). In the present study, we evaluated the potential of mGM-CSF as a cytokine adjuvant coadministered with the enhanced green fluorescence protein (eGFP) model antigen from a bicistronic SIV-based IDLV vaccine to evaluate its role in the immune response.
Our results indicate that expression of mGM-CSF did not enhance cellular immune responses against eGFP and that inclusion of mGM-CSF expressed from SIV-based IDLV as a cytokine may have limited benefit as a vaccine adjuvant.
Materials and Methods
Vector construction
A schematic depiction of the transfer vectors is provided in Figure 1. The parental SIV-based self-inactivating lentiviral transfer vector expressing eGFP (pGAE-GFPW) has been described (25). To obtain plasmid pGAE-GFPmZ expressing eGFP and mGM-CSF from a bicistronic RNA, a DNA fragment containing the internal ribosomal entry site (IRES) sequence followed by the mGM-CSF coding sequence and the woodchuck hepatitis post-transcriptional regulatory element (WPRE) was obtained from plasmid pTY2-JRmZ (30) using SalI/AvaI restriction enzymes and cloned into the corresponding sites of pGAE-GFPW. To obtain plasmid pGAE-GFP-IRESLucW, expressing eGFP, and Luciferase from a bicistronic RNA, plasmid pcR2.1-IRES-Luc was restricted with SalI/BamHI, and the IRES-Luc coding sequence was inserted downstream the eGFP coding sequence in the pGAE-GFPW plasmid using the corresponding sites. The IN-defective packaging plasmid pAd-D64V, containing an aminoacid mutation in the IN catalytic triad (D64V) abolishing the IN activity, and the Envelope plasmid pMD.G, producing the pseudotyping vesicular stomatitis virus envelope glycoprotein G (VSV.G), have already been described (25).
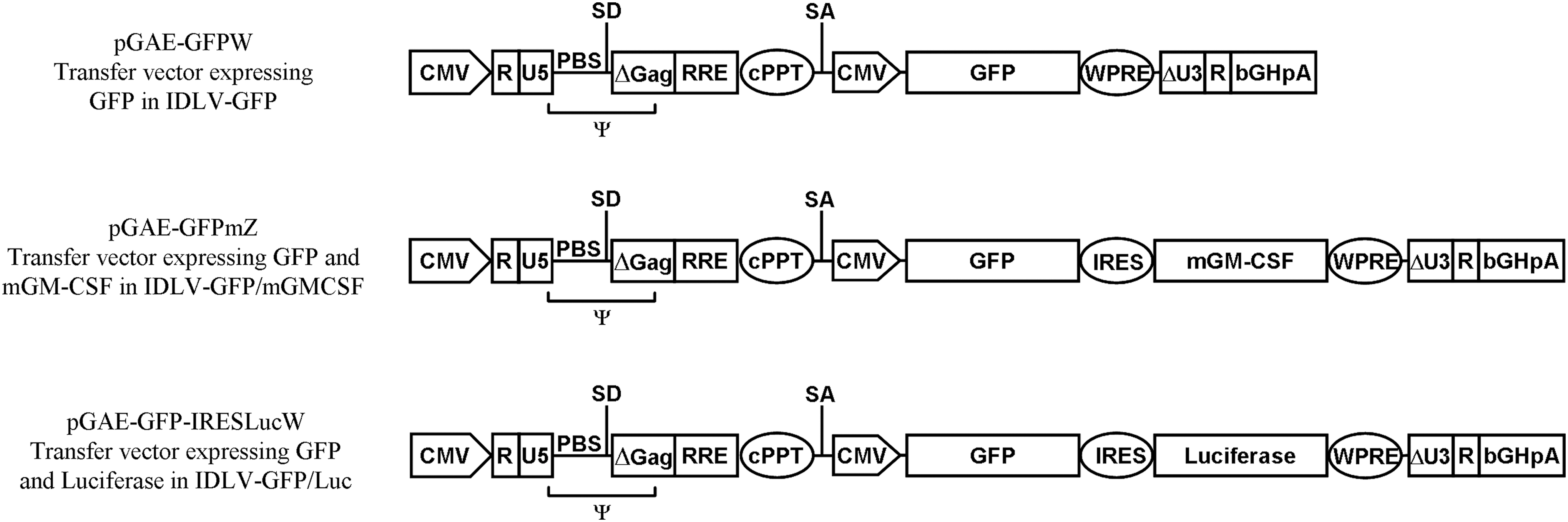
Schematic layout of transfer vectors used in the study and used for production of IDLV. The eGFP, mGM-CSF, and Luciferase coding sequences are under the control of cytomegalovirus (CMV) promoter. The packaging signal (Ψ), primer binding site (PBS), major splice donor (SD), splice acceptor (SA) sites, bovine growth hormone polyadenylation signal (bHGpA), central polypurine tract (cPPT), internal ribosomal entry site (IRES), and woodchuck hepatitis post-transcriptional regulatory element (WPRE) are indicated. The pGAE-GFPW is a monocistronic construct expressing the eGFP gene; the pGAE-GFPmZ and pGAE-GFP-IRESLucW are bicistronic constructs expressing either eGFP and mGM-CSF (pGAE-GFPmZ) or eGFP and Luciferase (pGAE-GFP-IRESLucW). IDLV, integrase-defective lentiviral vectors; eGFP, enhanced green fluorescence protein; mGM-CSF, murine granulocyte–macrophage colony-stimulating factor.
Production of recombinant vectors
The human epithelium kidney 293T-cell line was maintained in Dulbecco's Modified Eagle's medium (DMEM; Gibco, Life Technologies Europe, Monza, Italy) supplemented with 10% fetal bovine serum (FBS; Biowest, Nuaillé, France) and 100 U/mL of penicillin–streptomycin–glutamine (PSG; Gibco). For production of recombinant SIV-based IDLV (IDLV-GFP, IDLV-GFP/mGM-CSF and IDLV-GFP/Luc), cells were transfected in 100-mM-diameter Petri dishes using the calcium phosphate-based ProFection Mammalian Transfection System (Promega Corporation, Madison, WI) using a total of 12 μg of plasmid DNA for each plate in a ratio 6:4:2 (transfer vector: packaging vector: VSV.G vector), as described (25). Vectors containing supernatants were concentrated by ultracentrifugation (Beckman Coulter, Inc., Fullerton, CA) for 2 h at 27,000 rpm on a 20% sucrose gradient (Sigma Chemicals, Co., St. Louis, MO). Finally, the viral pellets were resuspended in 1× phosphate buffered saline (PBS) and stored at −80°C for further analyses. Viral titers for the eGFP-coding vectors were normalized by exogenous reverse transcriptase (RT) activity assay and titration on 293T cells for eGFP expression (4,12).
Murine GM-CSF ELISA and biological activity
The 293T cells were seeded at 5×104 per well in 6-well plates. Next day, cells were transduced with normalized amounts of IDLV (IDLV-GFP, IDLV-GFP/mGM-CSF, and IDLV-GFP/Luc). Seventy-two hours post-transduction media were recovered and production of mGM-CSF was quantified by ELISA (Immunological Sciences, Rome, Italy), while cells were evaluated for eGFP expression. The biological activity of mGM-CSF was determined using the mouse lymphoblast cell line FDC-P1 (ATCC, CRL-12103). FDC-P1 cells depend on WEHI-3 (ATCC, TIB-68)-conditioned medium for continued growth and proliferate upon mGM-CSF addition (19). WEHI-3 cells (14), kindly provided by Dr Mauro Valtieri (ISS, Rome, Italy), were maintained in Iscove's Modified Dulbecco's Medium (IMDM; Gibco) supplemented with 10% FBS (Gibco) and 100 U/mL of penicillin streptomycin (PS) (Gibco). FDC-P1 cells, a gift from Dr. Barbara Bottazzi (Istituto Clinico Humanitas, Milan, Italy), were maintained in DMEM (Gibco) supplemented with 10% FBS (Gibco), 100 U/mL of PS (Gibco), and 25% conditioned medium of WEHI-3 cells. Serial 1:2 dilutions of supernatants of 293T cells transduced with IDLV-GFP/mGM-CSF or IDLV-GFP/Luc were mixed with FDC-P1 cells (5×104 cells/well) in a total volume of 100 μL. As a reference standard, serial 1:2 dilutions of recombinant mGM-CSF (PeproTech EC Ltd., London, United Kingdom) (range 2.5–0.005 ng/mL) or DMEM alone were cultured with the cells. After 48 h of incubation at 37°C, the viability of the cells was quantified using the CellTiter 96 Aqueous Assay (Promega).
Mice immunization
Six- to 8-week-old BALB/c female mice (Harlan Italy, S. Pietro al Natisone, Italy) were used. Mice were housed and fed in separate cages, according to the experimental group, in accordance with the European Union guidelines and Italian legislation. All studies were approved by the Internal Institutional Review Committee. Mice were injected once intramuscularly with 0.2 mL of viral preparation. In the first experiment, groups of four mice were immunized with 2.7×107 RT units of IDLV-GFP or IDLV-GFP/mGM-CSF in PBS 1×. In the second experiment, groups of four mice were immunized with 3.5×106 RT units of IDLV-GFP/mGM-CSF or IDLV-GFP/Luc. Naive nonimmunized mice were kept for parallel analysis. On day 30, mice were bled orbitally under metaphane-induced anesthesia to collect whole blood in K-EDTA anticoagulant. Leukocytes were obtained after ammonium chloride potassium (ACK) lysis and used in the interferon-γ (IFN-γ) enzyme-linked immunosorbent spot (ELISPOT) assay. On day 90, mice were euthanized and spleens and draining lymph nodes were taken under sterile conditions for immunological analysis. Single-cell suspensions were obtained by mechanical disruption, passaged through cell strainers (BD Pharmingen, San Diego, CA), transferred on tubes containing the complete RPMI 1640 medium, and used in the assays.
IFN-γ ELISPOT assay
The IFN-γ ELISPOT assay was performed using reagents from Mabtech (Mabtech AB Gamla Värmdöv, Sweden), as described (25,30). A 9mer containing the H-2d restricted GFP-9mer peptide (HYLSTQSAL; UFPeptides s.r.l., Ferrara, Italy) (25) was used to stimulate GFP-specific CD8 T cells. Medium alone and unrelated H-2d restricted HIV-1 gp120 V3 loop epitope peptide (JR-9mer, IGPGRAFYT; UFPeptides) (30) were used as negative controls, while concanavalin A (5 μg/mL; Sigma) was used as positive control. Spot-forming cells (SFC) were counted with an ELISPOT reader (A.EL.VIS, Hannover, Germany) and results expressed as SFC/106 cells. Samples (specific and unrelated peptide-treated wells) were subtracted from the values obtained in medium-treated wells (background) and were scored positive when a minimum of 50 spots per 106 cells and a fold of two or higher compared to the unrelated peptide were observed.
Intracellular staining for cytokines
Splenocytes from each immunized group were pooled and cultured in the presence of GFP-specific (5 μg/mL) or JR-unrelated (5 μg/mL) peptides for 6 h in the presence of anti-mouse CD28 mAb (clone 37.51; BD Pharmingen) at 2 μg/mL. PMA (50 ng/mL; Sigma) and ionomycin (2 μg/mL; Sigma) were used as positive control. One hour after stimulation, 10 μg/mL of Brefeldin A (Sigma) was added to the cultures to inhibit cytokine secretion. Cells were membrane stained with direct fluorochrome conjugates PE-Cy5 anti-mouse CD8 (clone 53-6.7; BD, Pharmingen) or the isotype-matched mAb (BD Pharmingen), washed, fixed with 4% paraformaldehyde (Sigma), permeabilized in PBS–0.5% saponin (Sigma), and stained with APC-labeled anti-mouse IFNγ mAb (clone XMG1.2), PE-labeled anti-mouse tumor necrosis factor-alpha (TNF-α, clone MP6-XT22), and PE-labeled anti-mouse interleukin-2 (IL-2, clone JES6-5H4) or their isotype-matched controls (BD Pharmingen). Samples were washed and analyzed by a flow cytometer (FACScanto; BD Biosciences).
In vitro stimulation of effector cells and 51chromium release assay
Splenocytes from mice sacrificed at 90 days from immunizations were cultured in 24-well plates (2×106 cells/mL) in the presence of GFP-9mer peptide pulsed and irradiated syngeneic cells as stimulators. Target WEHI 164 cells were incubated with Na2 51CrO4 for 2 h at 37°C and washed thrice with a cold medium. The labeled cells were then split in two tubes; in one tube, the 9mer peptide was added at 5 μg/mL for 1 h in a serum-free medium. After washing, 1×104 target cells were added to the effectors at different effector:target ratios and incubated for 4 h at 37°C in 96-well plates. Finally, 40 μL of supernatant/well was harvested on a LumaPlate96 (Perkin Elmer Life Sciences, Boston, MA) and the emitted radioactivity was counted with a MicroBeta scintillation counter (Perkin Elmer). The spontaneous and total release was evaluated in wells containing target cells with either medium or 0.1% TritonX-100 (Sigma), respectively. The percentage of specific lysis was calculated as [(test release-spontaneous release)/(total release-spontaneous release)]×100. Spontaneous release of target cells was <10% in all assays. The intra-assay variability was determined on duplicate wells.
Statistical analyses
Statistical analyses were performed by the nonparametric Mann–Whitney U test. All p-values were two tailed and considered significant if less than 0.05. All analyses were performed using SPSS for Windows version 20.0 (IBM, Somers, NY).
Results
Expression and biological activity of mouse GM-CSF
Transduction of 293T cells with IDLV-GFP/mGM-CSF confirmed that mGM-CSF was produced in the supernatants of transduced cells 3 days after infection (average: 5,424.6 pg/mL, range: 4,872.5 to 5,872.5 pg/mL). To verify the biological activity of recovered mGM-CSF, a biological assay on FDC-P1 cells was performed. FDC-P1 cells were incubated with serial dilutions (range: 1:2 to 1:32) of the supernatants from 293T cells transduced with IDLV-GFP/mGM-CSF and IDLV-GFP/Luc, as described in Materials and Methods. Results showed that IDLV-GFP/mGM-CSF-produced mGM-CSF stimulated proliferation of FDC-P1 cells, whose growth is dependent on mGM-CSF, at all the tested dilutions (Fig. 2). Importantly, the activity of mGM-CSF present in the supernatants was similar to the activity of equal concentration of recombinant mGM-CSF (data not shown). Conversely, supernatants from 293T cells transduced with IDLV-GFP/Luc did not support proliferation of FDC-P1 cells.
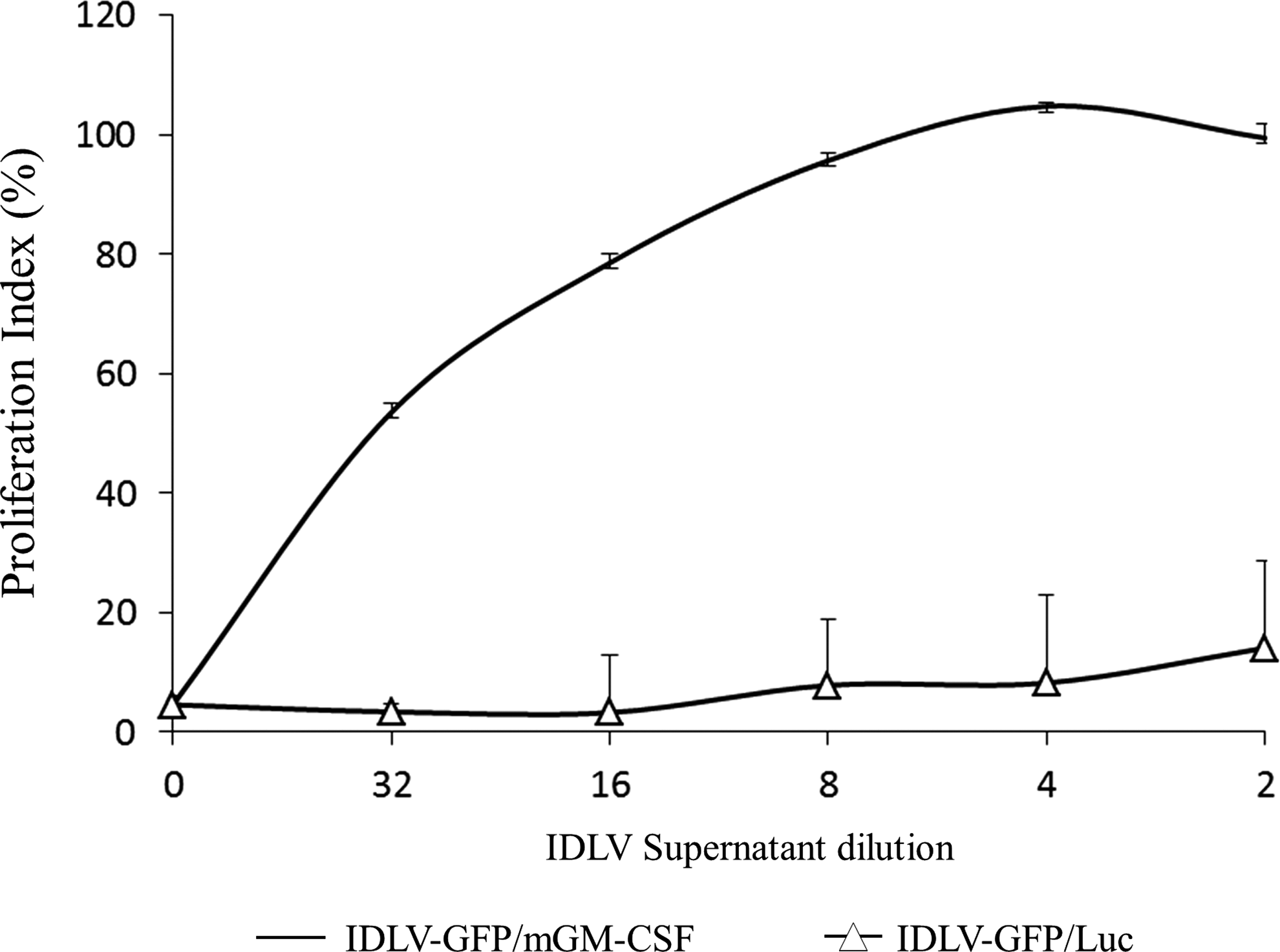
Biological activity of mGM-CSF produced from IDLV-GFP/mGM-CSF. The FDC-P1 cells were incubated with serial dilutions of supernatants from 293T cells transduced with IDLV-GFP/mGM-CSF (—) or with IDLV-GFP/Luc as vector control (−Δ−). Proliferation of cells was measured by the CellTiter 96 Aqueous Assay (Promega) and determined at 490 nm. The 1:2 serial dilutions of supernatants were in a range of 1:2 to 1:32 (corresponding to 2.7 to 0.17 ng/mL input mGM-CSF, as calculated by ELISA). Results are presented as the mean from two independent experiments performed in quadruplicates. Error bars represent the SD of the average.
GFP-specific T-cell response in IDLV-GFP/mGM-CSF vaccinated mice is lower than in IDLV-GFP immunized animals
In the first set of experiments, mice received a single injection of IDLV-GFP or IDLV-GFP/mGM-CSF. Naive mice were used as controls. At 30 and 90 days after the inoculum, mice were sacrificed and splenocytes were evaluated for immunological responses by IFN-γ ELISPOT assay, chromium release assay (cytotoxic T lymphocyte [CTL] activity), and the presence of polyfunctional GFP-specific CD8+ T-cell responses in splenocytes by intracellular cytokine staining (ICS) for IFN-γ, IL-2, and TNF-α, following stimulation with the specific GFP-9mer and the unrelated JR-9mer peptides. Surprisingly, by all the performed assays, in mice immunized with IDLV-GFP, the GFP-specific response was significantly higher than that recovered in mice immunized with IDLV-GFP/mGM-CSF (Supplementary Fig. S1; Supplementary Data are available online at
To understand the unexpected results obtained after immunization in mice, we wondered whether the presence of IRES in the IDLV-GFP/mGM-CSF vector was detrimental to the expression of the transgene, as described in other settings (3,5,10). 293T cells were transduced with normalized amounts (measured as RT activity) of IDLV-GFP, IDLV-GFP/mGM-CSF, or IDLV-GFP/Luc viruses. At 3 days post-transduction, cells were analyzed for eGFP expression by flow cytometry analysis. Results showed that the bicistronic IDLV showed 80% reduction of eGFP expression compared to the IDLV expressing eGFP alone (Fig. 3). This prompted us to reconsider our previous immunization schedule by performing new immunization experiments aimed at comparing T-cell responses after immunization with IDLV-GFP/mGM-CSF and IDLV-GFP/Luc to verify the mGM-CSF activity in the context of similar eGFP transgene expression.
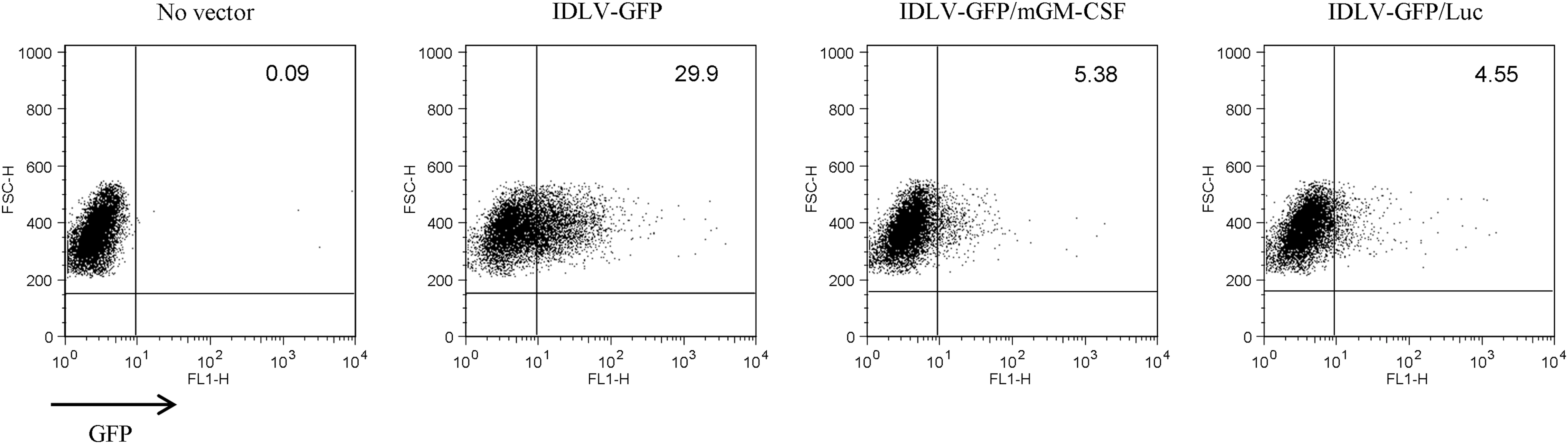
Transduction of 293T cells with IDLV expressing eGFP. 293T cells were transduced with normalized amounts of IDLV-GFP, IDLV-GFP/mGM-CSF, or IDLV-GFP/Luc (0.3×105 RT counts/1×105 cells) or left untreated (no vector). At 3 days post-transduction, cells were analyzed by FACS to evaluate the percentage of eGFP-expressing cells. A representative experiment out of three performed is shown. FACS, flow cytometry; RT, reverse transcriptase.
mGM-CSF delivered by IDLV does not increase the transgene-specific immune response
In this second set of experiments, mice received a single injection of 3.5×106 RT units per mouse of IDLV-GFP/mGM-CSF or IDLV-GFP/Luc. Naive mice were used as controls. IFN-γ ELISPOT assay was performed at 30 days after the single immunization on blood cells stimulated with the H-2d-specific GFP-9mer peptide or with the H-2d-matched JR-9mer unrelated peptide. Results showed that a similar GFP-specific response was present on cells derived from both groups of immunized mice (Fig. 4a). In particular, cells from IDLV-GFP/mGM-CSF displayed an average of 846 SFC/106 cells (range 545 to 1,070), while IDLV-GFP/Luc displayed an average of 510 SFC/106 cells (range 85 to 985). No statistical differences were seen between the two vaccinated groups (p=0.248). Specific responses were absent in naive mice or in cells pulsed with the H-2d-matched unrelated JR-9mer peptide.
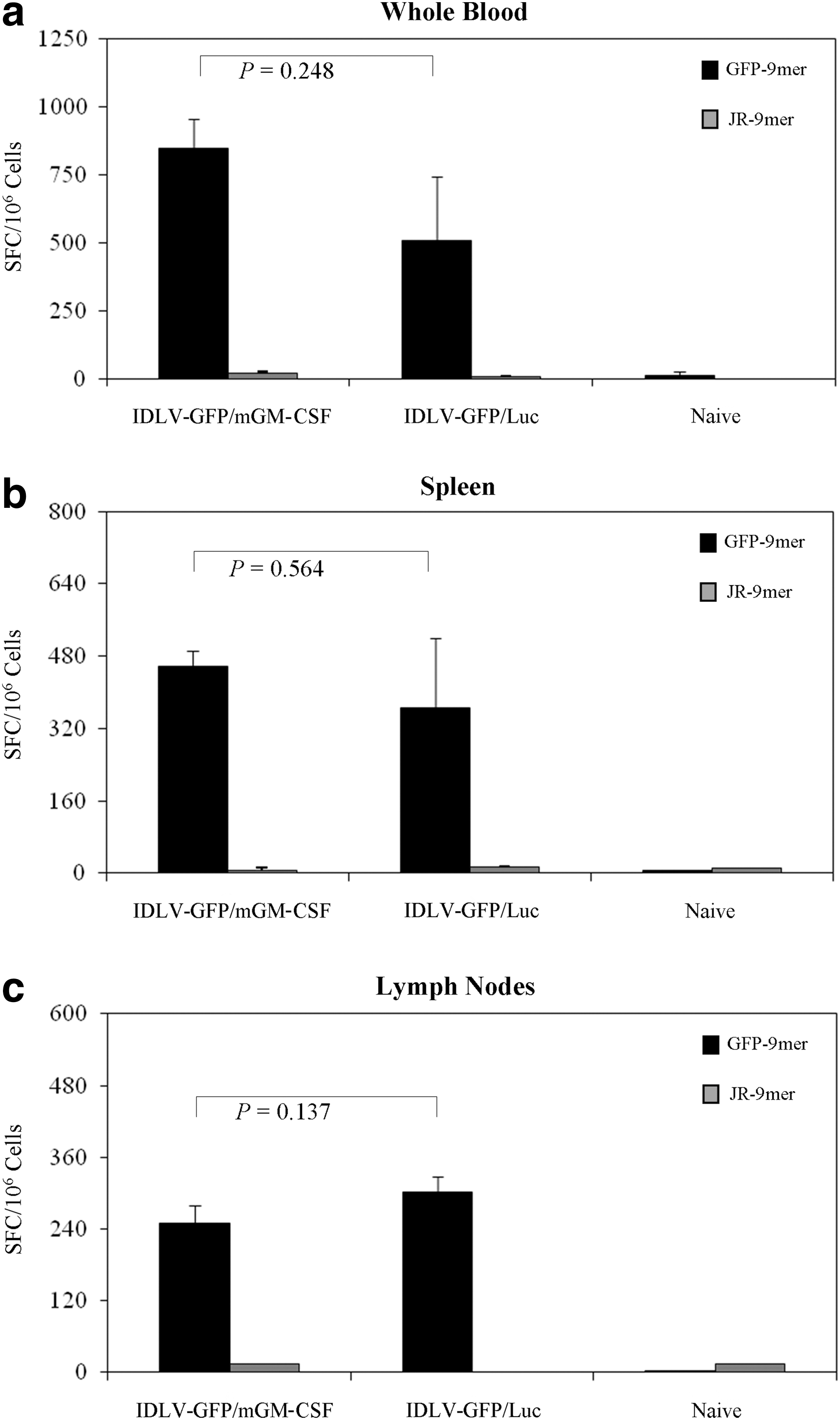
IFN-γ responses in mice immunized with IDLV expressing eGFP in combination with mGM-CSF or Luciferase. IFN-γ ELISPOT assay was performed on
At 90 days after immunization, analysis of splenocytes did not reveal significant differences in the number of responding cells in the vaccinated mice. In mice immunized with IDLV-GFP/mGM-CSF, SFC/106 ranged from 385 to 545 (average value of 458.1), while in mice immunized with IDLV-GFP/Luc, SFC/106 ranged from 160 to 747.5 (average value of 366.3) (p=0.564) (Fig. 4b). The presence of GFP-specific IFN-γ -producing T cells was also detected in the draining lymph nodes (Fig. 4c). In particular, the average values observed were 250 SFC/106 cells in mice immunized with IDLV-GFP/mGM-CSF and 302.5 SFC/106 cells in mice immunized with IDLV-GFP/Luc (p=0.317). The use of the H-2d-matched unrelated JR-9mer peptide did not show any response.
To analyze the effective ability of GFP-specific IFN-γ-producing CD8+ T cells in killing target cells, a chromium release assay was performed using splenocytes stimulated with irradiated naive splenocytes pulsed with the GFP-9mer peptide in both groups of mice. A specific lysis of target cells was present at all effector-to-target ratios analyzed in both groups of mice, reaching values of specific lysis 20.33% (IDLV-GFP/mGM-CSF) and 16.50% (IDLV-GFP/Luc) at effector-to-target ratios 80:1 (Fig. 5a). However, differences between the two groups were not statistically significant at any tested effector-to-target ratios (p=0.386).
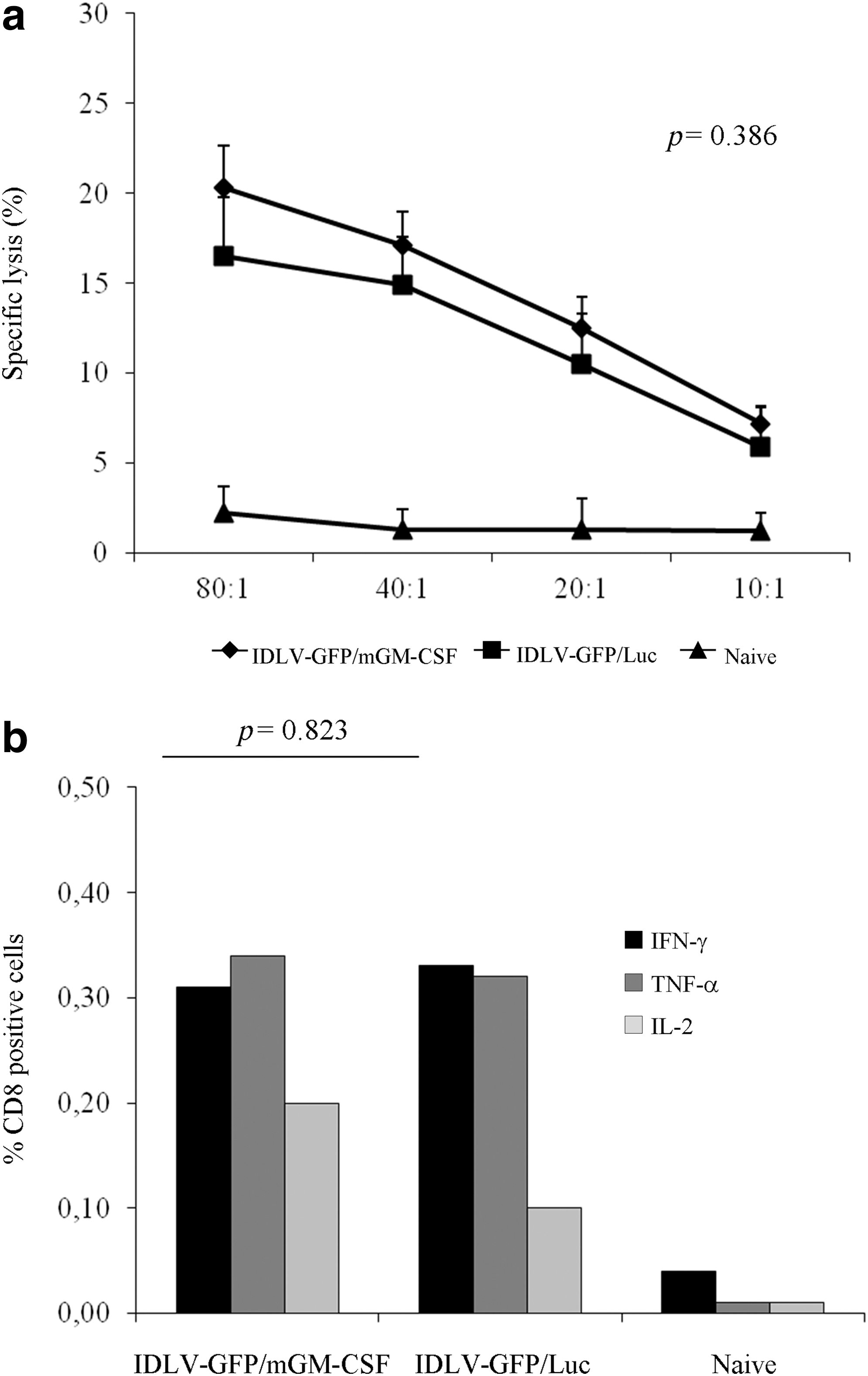
Cytotoxic activity and polyfunctional antigen-specific CD8+ T-cell response in mice immunized with IDLV expressing eGFP in combination with mGM-CSF or Luciferase.
To further characterize the CD8+ T-cell response, we evaluated polyfunctional GFP-specific CD8+ T-cell responses in splenocytes by ICS for IFN-γ, IL-2, and TNF-α, following stimulation with the specific GFP-9mer and the unrelated JR-9mer peptides. All inoculated mice showed CD8+ GFP-specific T-cell responses to all cytokines analyzed (Fig. 5b), although at about 1-log lower levels than those found in mice immunized with a higher dose of the IDLV-GFP/mGM-CSF (Supplementary Fig S1). No statistical differences were observed between the groups of immunized mice (p=0.823). Splenocytes from naive mice did not show any GFP-specific significant production of cytokines.
These results indicate that both vaccinations qualitatively and quantitatively induced similar CD8+ T-cell responses, and that the presence of mGM-CSF does not increase the immune response against the eGFP transgene.
Discussion
Immunization with IDLV results in the induction of a prolonged antigen-specific T-cell immunity against the delivered antigen with low interfering vector-specific responses (15). However, we reasoned that the inclusion of a cytokine adjuvant might further improve the immune response against the transgene. We focused our attention on GM-CSF, since plasmid delivery of GM-CSF was shown to recruit and activate DCs and macrophages at the site of inoculum (13,23), and IDLV-transduced DCs are able to trigger the effective induction and expansion of antigen-specific human CD8+ T cells in vitro (27) and in vivo (16).
Our results indicate that inclusion of mGM-CSF in the context of the IDLV-GFP does not improve T-cell immune response against the eGFP model antigen, as measured by GFP-specific IFN-γ ELISPOT, polyfunctional CD8+ T cells, and CD8+ T cell cytotoxic activity. This was unexpected, since coinjection with plasmid DNA expressing GM-CSF has been reported to increase the immune response against the transgene in several settings (37,48). However, other reports have shown the opposite (20,40,47,46), suggesting that the full potential of GM-CSF as a vaccine adjuvant remains to be established.
For delivering mGM-CSF from IDLV, we used a bicistronic vector expressing, from the CMV promoter, cap-dependent eGFP as immunogen and mGM-CSF from an IRES sequence, which is remarkable for its ability to initiate high levels of cap-independent protein synthesis in mammalian cells and cell-free extracts. This was considered advantageous, since simultaneous delivery of immunogen and adjuvant from the same transcript in the context of IDLV might be convenient for vaccination purposes. IDLV-produced mGM-CSF was biologically active, as confirmed in a proliferation assay using the FDC-P1 mouse lymphoblast cell line, which is growth-dependent on mGM-CSF (19). Conversely, we found that a lower level of eGFP protein was produced in both the IDLV containing the IRES sequence, IDLV-GFP/mGM-CSF, and IDLV-GFP/Luc, compared to IDLV-GFP, which does not contain an IRES sequence. This is in line with reports showing either that there is a direct competition between a functional IRES (in this case producing mGM-CSF) and the cap-dependent translation mode (in this case producing eGFP) or that the introduction of the eGFP sequence in the first cistron leads to decreased transduction efficiencies (3,5,10). This might explain why the GFP-specific T-cell response in IDLV-GFP/mGM-CSF vaccinated mice was lower than in IDLV-GFP immunized mice (Supplementary Fig. S1), while similar to that found in IDLV-GFP/Luc vaccinated mice (Figs. 4 and 5). Importantly, comparison of the immune response against eGFP in the IDLV-GFP/mGM-CSF and IDLV-GFP/Luc vaccinated mice clearly indicated that mGM-CSF does not improve the T-cell immune response against the eGFP immunogen in the context of a SIV-based IDLV. These results are consistent with previous studies using GM-CSF with a different delivery strategy and in clinical trials on oncologic patients, significantly better CD8+ and CD4+ responses were obtained in the absence of GM-CSF (20,34,39,40,41,46,47). Recent studies reported conflicting data on the role of GM-CSF as an immune adjuvant or therapeutic agent, and evaluation of T-cell response to a VSV-based vector expressing SIV gag showed that expression of GM-CSF may have limited vector replication excessively in the macaque model, but not in the murine model (40,38). A possible mechanism by which GM-CSF may interfere with immune function is through milk fat globule EGF-8 (MFG-E8) (17,18). GM-CSF induces expression of MFG-E8 by DCs and macrophages, and MFG-E8 can maintain regulatory T cells and downregulate Th1 responses (17,18). The failure of plasmid GM-CSF to augment plasmid DNA vaccine-elicited immune responses may be attributable to local increases in the production of certain cytokines like IFNs. In fact, IFNs may decrease vaccine immunogenicity by suppressing antigen expression (21,36).
In summary, our study shows that while immunization with SIV-based IDLV expressing eGFP was immunogenic in mice, even at low dose, addition of mGM-CSF does not enhance immune responses. In fact, CD8+ T-cell responses were lower when using the bicistronic IDLV and long-term memory CD8+ T-cell responses were not enhanced. Although this might have been ascribed to the lower amount of eGFP produced from the IDVL-GFP/mGM-CSF vector, CD8+ T-cell responses to eGFP in the IDLV-GFP/mGM-CSF immunized mice were nearly equivalent to those present in mice immunized with IDLV-GFP/Luc. Additional studies are needed to determine if mGM-CSF may alter the long-term quality of vaccine-induced T cells and whether the inclusion of mGM-CSF in the vaccine preparation may have an effect on clinical outcome, for example, after challenge in a disease model.
Footnotes
Acknowledgments
The authors wish to thank Prof. François-Loïc Cosset for providing the original pAd-SIV3+ packaging construct, Ferdinando Costa and Patrizia Cocco for technical support, Daniele Macchia for animal care, Marina Franco for editorial assistance, and Stefania Donnini for secretarial assistance. The research leading to these results has received funding from the European Union's Seventh Framework Programme [FP7/2007-2013] under Grant Agreement No: 280873 ADITEC, the Italian AIDS National Program (RF-ISS-2009-1300961 to A.C. and RF-ISS-2009-1302984 to D.R.M.N.), and the Italian Association for Cancer Research (AIRC-ISS grant n. IG 10115 to A.C.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.