
Abstract
Subacute sclerosing panencephalitis (SSPE) is caused by a persistent measles virus infection. Regulatory mechanisms can be responsible for a failure of immunosurveillance in children with SSPE. In this study, peripheral blood cells of 71 patients with SSPE and 57 children with other diseases were compared phenotypically. The proportions of CD4+, CD8+ T, and NK cells were homogenous, whereas total CD3+ T and Treg (CD4+CD25+CD152+) cells were decreased in patients with SSPE. The proportion of CD8+ T cells expressing the inhibitory NKG2A+ receptor was also decreased (1.7%±1.7% vs. 2.6%±1.9%, p=0.007) in patients with SSPE, whereas the proportion of NK cells expressing activating NKG2C was increased compared with the control group (30.0%±17.3% vs. 22.2%±17.0%, p=0.039). The decrease in the number of cells with regulatory phenotype, the lower presence of the inhibitory NK receptors on CD8+ cells, and higher activating NK receptors on NK cells in SSPE indicate an upregulation of these cell types that favors their response. This state of active immune response may be caused by chronic stimulation of viral antigens leading to altered regulatory pathways.
Introduction
S
On the other hand, the effect of inhibitory/activating NK receptors (NKR) on effector cells may also be operating in virus-specific CD8+ T cells or NK cells, which can be controlled by variable expression of their ligands during viral infection. NKR expressed on antiviral cells can control cytotoxic and cytokine-producing activities of T and NK cells (18). Among NKR, binding of heterodimers of CD94 to its ligand conveys an inhibitory signal when with NKG2A, whereas the signal through CD94/NKG2C has a costimulatory effect (2). CD94/NKG2A and CD94/NKG2C are normally expressed on most NK cells and on a small fraction of CD8+ T lymphocytes (20,37). Inhibitory NKR can differentially regulate the activities of CD8+ T cells, which protect against viruses that are completely cleared compared with those that establish persistent infection (21).
For the identification of the immune mechanisms required for the reactivation of the persistent MV, we analyzed probable Tregs and the expression of regulatory molecules on T and NK cells in SSPE patients.
Materials and Methods
Patients and controls
Seventy-one patients with SSPE (45 boys and 26 girls, mean age: 10±6 years) were recruited for the study between the years 2002 and 2013. All patients had increased anti-MV antibody titers and oligoclonal IgG bands in CSF, with typical electroencephalographic and clinical features fulfilling the criteria for SSPE (12,13,25). According to the clinical staging, 10 patients had early findings (1 patient with 1B and 9 patients with 2A) and 32 patients had more severe disease at 2B (n=12), 2C (n=14), 2D (n=3), 2 (n=3), and 4 (n=1) stages. Fifty patients had natural measles between the ages of 3 months and 9 years. The measles history of the remaining 17 patients was unknown. Thirty-seven of the patients had a history of measles vaccination. Informed consent was obtained from the parents, and the Ethics Committee approval was taken locally.
Fifty-seven children (23 girls and 34 boys, mean age: 9±6 years) were used as controls (CON). Twenty-eight (mean age: 9±4 years) had inflammatory diseases (inflammatory controls [ICON]) such as upper respiratory tract infections, asthma, and otitis media, and 29 (mean age: 10±7 years) had noninflammatory diseases like epilepsy and Lafora (non-inflammatory controls [NICON]) or no specified symptoms. Due to the ethical considerations of taking blood from healthy children, we could not include healthy donors.
Phenotypic staining and reagents
Erythrocyte-lysed peripheral blood cells of donors were surface stained with fluorochrome-conjugated mouse anti-human CD3-APC, CD8-APC, CD4-APC, CD25-FITC (all IgG1; DakoCytomation, Glostrup, Denmark), CD56-FITC (IgG2a; Serotec, Oxford, United Kingdom), CD152-PE (anti-CTLA-4, IgG2a), CD94-FITC (IgG1), CD28-PE (IgG1; BD Biosciences Pharmingen, Erembodegem, Belgium), NKG2A-PE (IgG2a), NKG2C-PE (IgG1; R&D Systems, Minneapolis, MN), CD45-FITC/CD14-PE (IgG1/IgG2a; Diaclone, Stanford, CT) antibodies, and isotype control antibodies (BD Biosciences Pharmingen, DakoCytomation, Glostrup, Denmark). Cell subset phenotypes were analyzed by flow cytometry (FACSCalibur; Becton-Dickinson) and compared in both lymphocyte (CD45+CD14−) and respective cell-type gates (CD8+, CD4+, CD56+) between groups (Supplementary Fig. S1; Supplementary Data are available online at
Statistical analyses
Statistical analyses were performed by the nonparametric Mann–Whitney U test for comparisons. Results are presented as mean values. As no significant difference was detected between the ICON and NICON groups, we evaluated all control donors as a single CON group.
Results
Distribution of T and NK cells in the blood
To determine the proportions of CD3+, CD4+, CD8+, and CD8−CD56+ cells in patients with SSPE and CON, lymphocyte gate (CD45+CD14−) was applied. The proportion of CD3+ T cells was slightly decreased in patients with SSPE compared with age-matched CON (60.9%±12.8% vs. 66.1%±11.4%, p=0.028). The lower percentage of CD3+ cells in patients with SSPE could not have been caused by an altered ratio of CD4+ or CD8+ cells because no statistically significant difference was observed in CD4+ and CD8+ T cells between the groups (33.9%±11.4% vs. 33.4%±10.8% and 27.5%±8.7% vs. 26.2%±8.6%, respectively). When the distribution of CD8−CD56+ NK cells was compared, the proportions were similar in both groups (6.0%±5.1% vs. 6.1%±4.7%, Fig. 1).
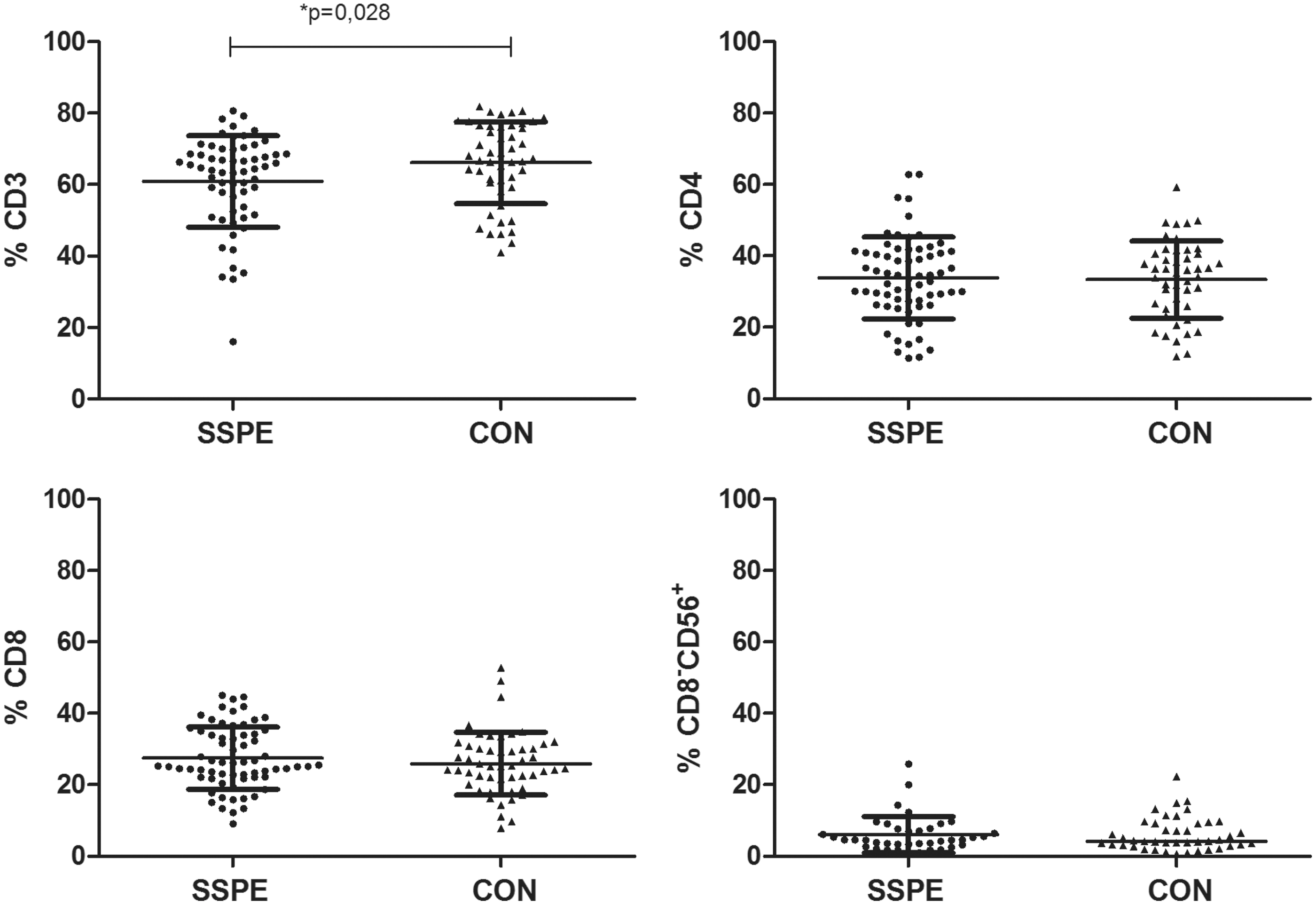
The proportions of CD3+, CD4+, CD8+ T cells and CD8−CD56+ NK cells in subacute sclerosing panencephalitis (SSPE) and CON donors. CD3+ T cells are slightly decreased in SSPE patients compared to CON. Results are presented with mean (horizontal lines)±SD values. Black circles, SSPE donors; black triangles, CON donors.
Regulatory T cells in SSPE patients
As possible regulatory cells, we analyzed CD4+CD25high, CD4+CD152+ cells, and the suppressor phenotype CD8+CD28− cells. The proportion of most probable Tregs, CD4+CD25high cells, was lower in patients with SSPE than CON (1.0%±4.1% vs. 1.6%±2.8%, p=0.043, Fig. 2A). When Tregs were identified as CD152+ on CD4+CD25+ cells, the proportion of these cells in lymphocytes was lower in the SSPE group compared with CON, with a tendency to differ (2.0%±2.7% vs. 4.0%±3.7%, p=0.051, Fig. 2B). CD4+CD152+ cells were also downregulated in patients with SSPE compared with CON (0.6%±0.7% vs. 1.1%±0.8%, p=0.003, Fig. 2C). The distribution of CD8+CD28− cells was not different between groups (data not shown).
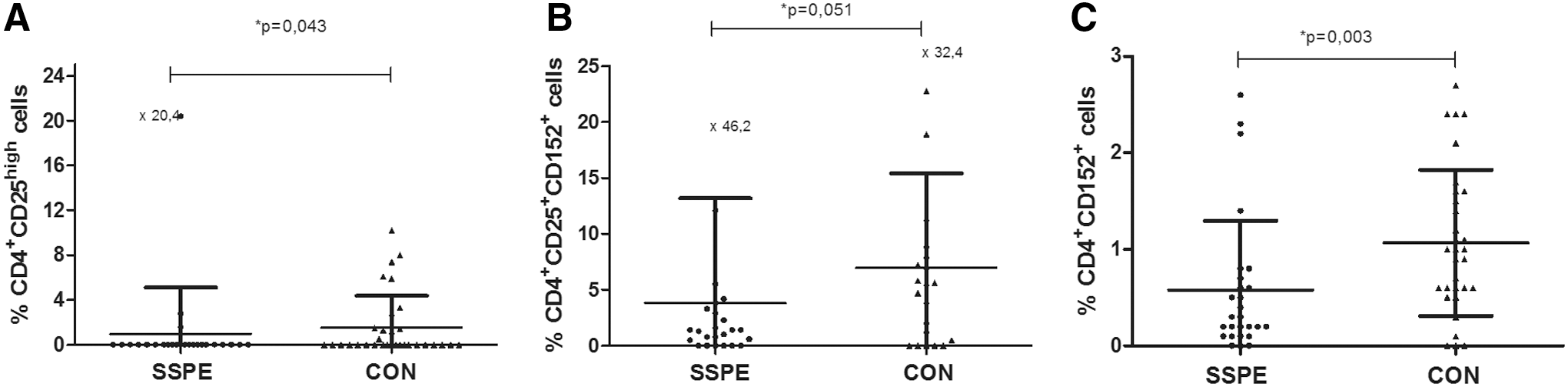
Proportions of regulatory T cells (Tregs) in SSPE patients and CON. The distribution in groups and mean (horizontal lines)±SD values are presented.
Regulatory receptor expression on NK cells
To investigate the expression of heterodimeric inhibitory NKG2A or activating NKG2C receptors in patients with SSPE, we compared the presence of these molecules on NK cells. The presence of inhibitory NKG2A molecules on CD8−CD56+ NK cells as well as CD56+NKG2A+ double-positive cells among the lymphocytes was not different in the SSPE and CON groups (27.5%±21% vs. 29.6%±20% and 2.4%±2% vs. 3.5%±3%, respectively, Fig. 3A). Although the CD56+NKG2C+ double-positive cells were relatively low in both groups (2.3%±1.7% vs. 2.4%±2.3%), the expression of the activating NKG2C on CD56+CD8− cells was higher in patients with SSPE compared with CON (30.0%±17% vs. 22.2%±17%; p=0.039, Fig. 3B).
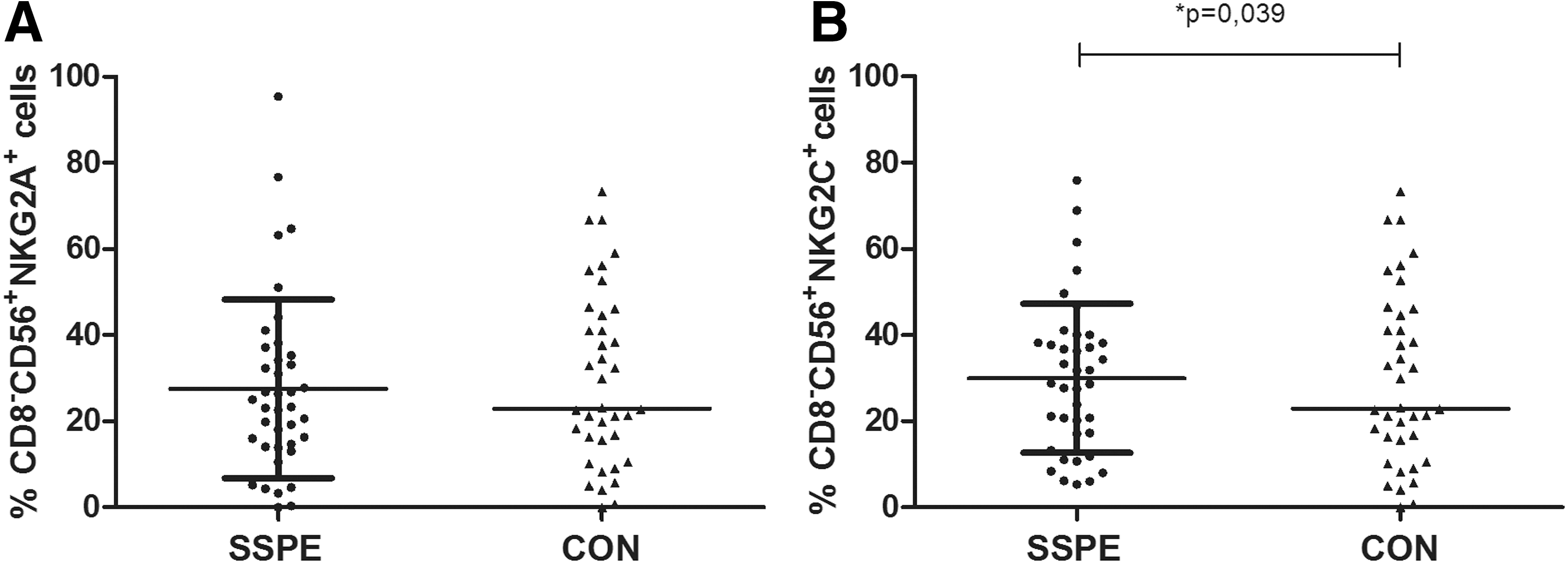
Analysis of NKG2A and NKG2C expression on CD8−-CD56+ NK cells in the SSPE and CON groups. Mean (horizontal lines)±SD values are presented.
Regulatory receptor expression on CD8+ T cells
Next, we analyzed the expression of the same NKR on CD8+ T cells. Although the proportions of CD8+ T cells were not different between the groups, the percentage of CD8+CD94+ cells in lymphocytes was lower in patients with SSPE than in CON (4.3%±2% vs. 5.9%±3%; p=0.056). This cell group was further evaluated for the receptors with activating or inhibitory heterodimers. The proportion of CD8+ cells expressing the inhibitory NKG2A was lower in patients with SSPE than in the CON group (1.7%±1.7% vs. 2.6%±1.9%; p=0.007, Fig. 4A), whereas CD8+ cells expressing the activating NKG2C were not different between groups (3.1%±3.0% vs. 3.2%±3.0%, Fig. 4B).
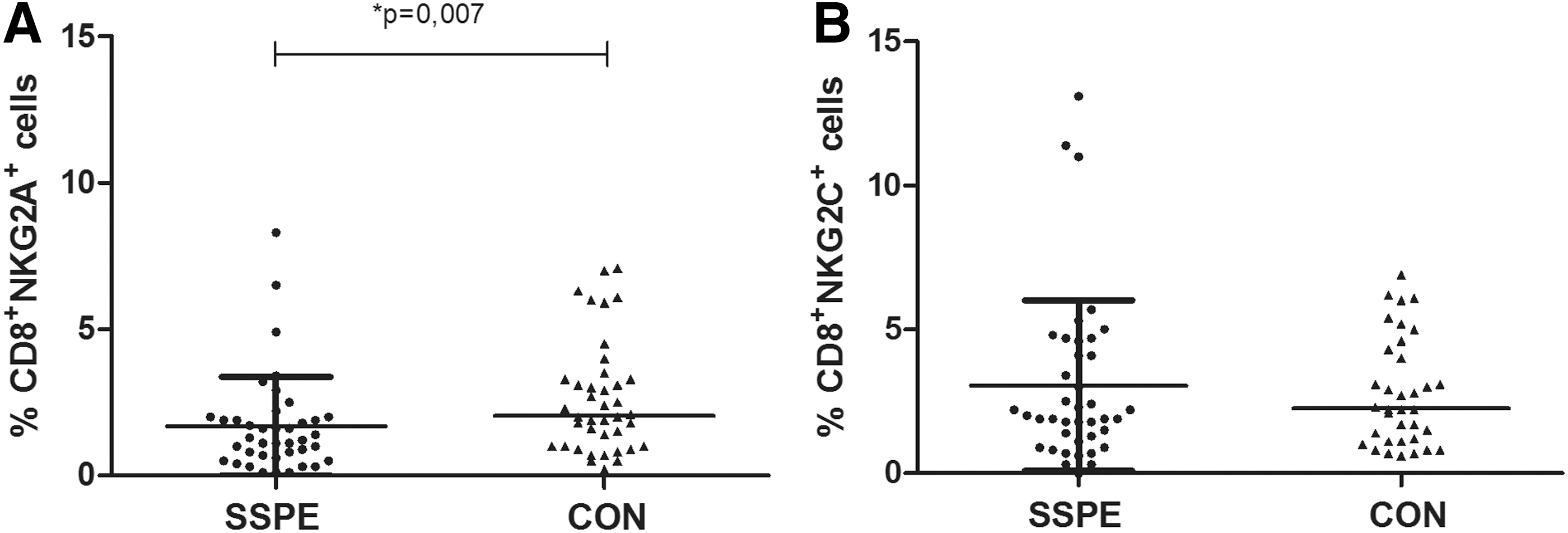
The expression of NKG2A and NKG2C on CD8+ cytotoxic T cells. Mean (horizontal lines)±SD values are presented.
A positive correlation between CD8+CD94+ cells and CD8+NKG2A+ cells was observed only in the CON (r=0.46, p=0.006) and not present in SSPE, which implicated a dysregulation of these surface molecules in SSPE. CD8+CD94+ and CD8+NKG2C+ cells were correlated in both the SSPE and CON groups (r=0.59, p=0.001 and r=0.41, p=0.02, respectively).
Discussion
To characterize the general state of the immune system during the persistent MV infection, we analyzed the regulatory cells and molecules with regulatory function on the blood cells in SSPE. By comparing the patients with SSPE with controls, we noted a decrease in the number of cells with regulatory phenotype and an increase of activating receptors on NK cells but not on CD8+ cells, which indicated alterations in both cell types.
Previous studies in SSPE revealed lower proportions of CD3+ T cells (27), but increased percentage of CD8+ T and NK cells (1,31). In this relatively large cohort of patients with SSPE, the decrease of CD3+ T cells was replicated, whereas the total NK and CD8 cells were not increased. The decrease in the number of CD3+ cells may be caused by MV-infected monocyte-induced apoptosis of these cells (33).
Measles is a highly contagious childhood disease that is associated with a strong virus-specific immune response resulting with the establishment of lifelong immunity. Measles infection is also followed by an acute and profound immunosuppression leading to an increased susceptibility to secondary infections (14). The immunosuppression observed after the acute infection may also be a possible link to viral persistence that causes late disease forms such as SSPE. However, our findings indicate that no suppression by Tregs is evident during the clinical chronic phase of established disease.
Along with CD25 and FOXP3, the CD4+ regulatory T-cell population requires CTLA-4 as a costimulatory molecule for stronger functional activation (4,30). Due to the limited volumes of blood available from these rare disease patients, surface staining only is preferred for the identification of Tregs cells and no functional evaluation was possible. The increase of CD4+CD25+ Tregs has been demonstrated in adults as well as in animal models in the acute phase of an MV infection (28,36). Despite the increased frequency of FOXP3+CD25+CD4+ Tregs both in the periphery and in the brain, specific antiviral responses could be demonstrated, whereas the capability of T lymphocytes to respond to virus-unrelated antigens was strongly suppressed. This finding implicated that MV affects regulatory T-cell homeostasis and that MV-induced Tregs could have favored the development of persistence and SSPE (29). However, in a persistent MV infection model, depletion of CD4+CD25+FOXP3+ Tregs increased virus-specific CD8+ effector T cells in the brain and caused a reduction of the persistent infection (26). Viral persistence was also shown to be relatively independent of CD25+CD4+ Tregs in another recent model of acute viral infection (8). The present results do not provide evidence for the effect of CD4+CD25high Tregs in the persistent brain disease stage. Lower numbers of CD4+CD25high T cells in the patients with SSPE may be related to the late time point of the disease and a compensatory mechanism induced by viral persistence for a more efficient viral clearance.
Expression of NKR on NK and CD8+ T cells, which may be involved in the cellular responses in SSPE, was also investigated in this study. NK cells may respond to persistent virus by expressing the activating receptor NKG2C, proliferate, and maintain in numbers for a prolonged period of time dependent on virus-induced ligands (17). Human cytomegalovirus and human immunodeficiency virus (HIV) infections are associated with increased percentages of a particular NK cell subset expressing the activating receptor CD94/NKG2C (23,32). In SSPE, the relative increase of activating NKG2C on NK cells in the periphery may also represent the continuous but inefficient activity during a sustained viral infection.
CD8+ T lymphocytes mediate immunosurveillance against persistent virus infections. NKR-expressing CD94/NKG2+CD8+ T cells are oligoclonal populations characterized by an effector memory phenotype, suggesting that they could be the result of antigen-specific expansion in vivo (5). However, pathogen-specific CD8+ T cells may also initiate expression of the inhibitory CD94/NKG2A upon activation and this expression maintains long term in persistent infections by repeated TCR stimulation (6,19). The percentage of CD8+ T lymphocytes expressing the inhibitory heterodimer was shown to be significantly decreased in HIV-I-infected patients in comparison with noninfected controls, with no significant difference in activating CD94/NKG2C+ on CD8+ (37). A similar decrease of NKG2A on CD8+ with the accompanying increase of NKG2C on NK cells was recently demonstrated in an HIV infection with a higher viral load (32). Our results revealed close similarities to the HIV infection. No evidence for a mechanism of immunosuppression by either NK or CD8+ T cells is supported in SSPE by the present findings.
An increase of activating receptor on NK and the decrease of inhibiting receptor on cytotoxic T cells in SSPE favor an immune status where the antiviral response is probably facilitated through both cell groups, although it has not been sufficient to clear the virus efficiently. Our data cannot determine whether this is a result of high viral load or of sequestration of the specific cells into the CNS. Our results implicate that the downregulation of the inhibitory NKR NKG2A on CD8+ cells in SSPE may be caused by altered regulation of CD8+ T cells. Inhibitory NKG2A receptors may induce MV-specific CD8+ T-cell population to become cytolitically anergic, which may have contributed to virus persistence. This finding is also in accordance with our previous data on inefficient T-cell responses in patients with SSPE. These children may produce only an exhausted immune response, which causes viral persistence due to inefficient removal of the virus (35).
The findings of the present study may only reflect the cell distribution in the peripheral blood, and the possibility of an accumulation of the regulatory cell groups in the CNS cannot be excluded by the data in SSPE (22). A better understanding of the balance between measles-induced effector and regulatory mechanisms, both in the periphery and in the brain, may be of critical importance in the design of novel approaches for the prevention and treatment of measles pathology. Further investigations of T-cell functions of SSPE patients will elucidate the pathogenic mechanisms in this disease.
Footnotes
Acknowledgments
The authors thank the physicians who follow patients with SSPE, for referring their patients. Also, the critical reading by David Chapman is appreciated. The study is supported by grants from the Research Fund of Istanbul University (Project #: 192).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.