
Abstract
Postweaning multisystemic wasting syndrome (PMWS) is regarded as an immunosuppressive disease in pigs caused by porcine circovirus type 2 (PCV2). Immune inhibitory receptors, particularly programmed death 1/programmed death-ligands (PD-1/PD-Ls) are presumably involved in the immunopathogenesis of PMWS. The aim of this investigation was to examine the relationship of immune inhibitory receptors and immunocompromised by PMWS. Nine 45-day-old conventional pigs were selected from a farm where pigs exhibited typical signs of PMWS (wasting and respiratory disorders) and tested positive for PCV2 infection by polymerase chain reaction (PCR). Six pigs were selected as controls due to their notably healthy state and absence of PCV2 infection. Heparinized blood samples were taken from each pig for pathogen detection and isolation of peripheral blood mononuclear cells (PBMCs), from which mRNA expression of immunomodulatory molecule (PD-1, PD-L1, PD-L2, PTEN, CTLA-4, LAG-3, and Foxp3) and cytokines (IL-10, IL-2, and IFN-γ) was determined. Proliferation of PBMCs was also assessed by flow cytometry utilizing cellular labeling dilutions for detection. The mRNA levels of PD-L1 (p<0.01), PD-L2 (p<0.05), and PTEN (p<0.01) were remarkably increased in the PBMCs of diseased pigs compared to healthy pigs, whereas no change was observed for PD-1, CTLA-4, LAG-3, and Foxp3 expression. Cytokine IL-10 mRNA levels were significantly elevated (p<0.01), while IL-2 and IFN-γ mRNA levels tended to be only slightly increased in the PBMCs of affected pigs compared to healthy controls. The proliferation of PBMCs was also decreased in diseased pigs. These data suggest that overexpression of PD-L1 and PD-L2 mRNA is one mechanism by which immunosupression of PMWS pigs occurs, supporting a new therapeutic strategy focused on PD-Ls for pigs suffering from PMWS.
Introduction
P
The co-stimulatory pathway consisting of Programmed death-1 (PD-1) receptor and its ligands, PD-L1 (B7-H1; CD274) and PD-L2 (B7-DC; CD273), are involved in negative immunoregulation (2,17). The activation of the T- and B-cells and monocytes induces PD-1 expression (11). PD-L1 and PD-L2 are predominantly expressed on APCs (9). Several reports have shown that the upregulated expression of PD-1 and its ligands results in T-cell exhaustion and host immunoinhibition during chronic viral diseases (2,13,24). Interestingly, PD-L1 interacts specifically with B7-1 to inhibit T-cell proliferation (4). The characteristics of T-cell impairment are loss of cytolytic functions and inhibition of proliferation and cytokine production, especially with respect to interleukin-2 (IL-2) and interferon-γ (IFN-γ) (9). In addition, phosphatase and tensin homologue deleted on chromosome ten (PTEN) activity have been linked to immunoinhibitory processes of the PD-1/PD-Ls pathway. PTEN has been shown to have an important antiproliferative role through its inhibitory effect on PI3-kinase (PI3K) and the Akt signaling pathway (22). Interestingly, activation of the PD-1/PD-Ls pathway results in an increase in PTEN activity, and blockade of the PD-1/PD-Ls pathway restores exhausted T-cell function during chronic viral infection (2,9,25). These findings suggest that the PD-1/PD-Ls pathway could emerge as an important potential therapeutic target in infectious diseases (27). In addition, other immune inhibitory receptors, such as cytotoxic T-lymphocyte antigen-4 (CTLA-4), lymphocyte activation gene-3 (LAG-3), and forkhead transcription factor 3 (Foxp3) have been implicated in immunoregulation (12,18,26,28). However, little is known about the involvement of the aforementioned immunoinhibitory receptors and ligands in the immunopathogenesis of pigs suffering from PMWS.
The goal of this study was to evaluate the mRNA expression profile of PD-1, PD-L1, PD-L2, PTEN, CTLA-4, LAG-3, Foxp3, IL-10, IL-2, and IFN-γ in peripheral blood mononuclear cells (PBMCs) of pigs naturally infected with PMWS and the proliferation of these cells to clarify further the pathogenesis of PMWS. Confirmation of the involvement of the PD-1/PD-Ls pathway in PMWS could elucidate novel therapeutic options for this disease and encourage its study in other immunomodulatory fields.
Materials and Methods
Animals
Two groups of pigs were selected from a conventional farm in the Henan province of China. The naturally occurring PMWS group infected with PCV2 included nine 45-day-old pigs with approximately 1 week of progressive weight loss and respiratory disorder. The control group tested negative for PCV2 infection and included six 45-day-old healthy pigs without any previous clinical signs of PMWS.
Detection of viral nucleic acids by polymerase chain reaction
Plasma samples were collected from heparinized venous blood of all pigs. Lung and spleen biopsies were taken from two diseased animals. Viral genomic DNA or RNA were extracted from plasma, lung, and spleen samples using a universal genomic DNA extraction kit (Takara Bio, Inc., Japan) or TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturers' instructions. cDNA was synthesized from ∼3 μL of RNA using PrimeScript™ RT reagent Kit (Takara) according to the manufacturer's manual. Briefly, the reverse transcription reaction was performed in 20 μL total volume with 2 μL 5×PrimeScript buffer, 0.5 μL PrimeScript RT enzyme mix I, 0.5 μL oligo dT primer, 0.5 μL random 6mers, 1 μg total RNA and RNase free dH2O at 37°C for 15 min and 85°C for 5 s in a water bath. Following the reaction, cDNA was put on ice to cool for several minutes.
Several viral nucleic acids were detected by polymerase chain reaction (PCR). Briefly, primers were used as follows: 5′-CAG CAC CCT GTA ACG TTT G-3′ and 5′-AGG AGT ACC ATT CCA ACG G-3′ for PCV2 (381 bp), 5′-GCG GCA TCG CCA ACT TCT T-3′ and 5′-GTC CTC CTT GAG CGT CTT CGT C-3′ for pseudorabies virus (PRV) (263 bp), 5′-TGG TAC AAG ACG ATG CAC AC-3′ and 5′-TCC TGT GGA GAA TTC ATC TC-3′ for porcine parvovirus (PPV) (509 bp), 5′-GAA GAA TAT GGG CTC ATG-3′ and 5′-GAT GAT GGC TTG AGC TGA GT-3′ for porcine reproductive and respiratory syndrome virus (PRRSV) (747 bp), 5′-CCC TGA AGT GGA TTA GAA-3′ and 5′-TAC CCT TGT TGA TCC TAT C-3′ for classical swine fever virus (CSFV) (111 bp). The amplification was performed in 20 μL reaction volumes containing 0.4 μM each primer, 0.1 mM dNTP, 0.5 μL (1 IU) DNA polymerase (Takara), 15.7 μL reaction buffer, and 2 μL viral nucleic acid. The cycling conditions consisted of initial denaturation at 95°C for 60 s, followed by 35 cycles at 95°C for 30 s, 55°C for 30 s, 72°C for 30 s, and a final extension step at 72°C for 10 min. PCR products were analyzed by gel electrophoresis.
Isolation of PBMCs
Porcine PBMCs were isolated from 5 mL of fresh, heparinized blood samples by using gradient density centrifugation with Histopaque 1.077 (Sigma-Aldrich, St. Louis, MO) at 500 g for 30 min. Once the PBMCs were recovered from the plasma-Histopaque interphase, they were washed three times with phosphate-buffered saline (PBS) and resuspended in RPMI 1640 supplemented with 10% fetal bovine serum (FBS; Sijiqing, Hangzhou, China). The viability of PBMCs was assessed by trypan blue dye exclusion.
RNA extraction and cDNA synthesis
Total RNA was extracted from freshly isolated PBMCs using TRIzol reagent (Invitrogen) according to the manufacturer's instructions. Residual DNA was removed from the total RNA samples, and first-strand cDNA was synthesized from 1 μg of total RNA with a PrimeScript RT Reagent Kit with gDNA eraser (perfect real-time; Takara, code: RR047A) according to the manufacturer's protocol. Briefly, residual DNA was removed as follows: 10 μL total volume of 1 μg total RNA, 2 μL 5×gDNA eraser buffer, 1 μL gDNA eraser and RNase-free dH2O at 42°C for 2 min, and then put on the ice. For reverse transcription, 20 μL total volume was used with 10 μL of the reaction solution described above, 4 μL 5×PrimeScript buffer 2, 1 μL PrimeScript RT enzyme mix I, 1 μL RT primer mix, and 4 μL RNase-free dH2O. The reaction was performed at 37°C for 15 min, followed by incubation in an 85°C water bath for 5 s. The synthesized cDNA was cooled at 4°C for several minutes and then stored at −20°C until use in real-time quantitative PCR (RT-qPCR) reactions.
Expression analysis of porcine PD-1, PD-L1, PD-L2, PTEN, and cytokine mRNA by RT qPCR
Expression of mRNAs was evaluated by RT qPCR procedures as previously described (Yue F, 2014, unpublished data). Briefly, primers were used as follows: 5′-GCT TGT GGC TGT CAG ACT CC-3′ and 5′-CGA CCT TGT GCC TCT CCT-3′ for PD-1, 5′-AAT GGC GAG GAA GAC CTG AA-3′ and 5′-CAG CAG TAA ACC CCT GCA TCT-3′ for PD-L1, 5′-GCC AAC ACC AGC TAT ACC AAG AC-3′ and 5′-GGG ATG AAA ACA CGG AGC A-3′ for PD-L2, 5′-CCA TTG CCT GTG TGT GGT G-3′ and 5′-GGT TTC CTC TGG TCC TGG TAT G-3′ for PTEN, 5′-TCT TCA TCC CTG TCT TCT CCA AA-3′ and 5′-GCA GAC CCA TAC TCA CAC ACA AA-3′ for CTLA-4, 5′-CTC CTC CTG CTC CTT TTG GTT-3′ and 5′-CAG CTC CCC AGT CTT GCT CT-3′ for LAG-3, 5′-CCG TGG ACG AGT TCG AGT TT-3′ and 5′-CAT CCT TCT TTC CTT GGC TTT G-3′ for Foxp3, 5′-CAT TGC ACT AAC CCT TGC ACT C-3′ and 5′-GGC TCC AGT TGT TTC TTT GTG TT-3′ for IL-2, 5′-CCT GAC TGC CTC CCA CTT TC-3′ and 5′-GGG CTC CCT AGT TTC TCT TCC T-3′ for IL-10, 5′-GAG GTT CCT AAA TGG TAG CTC TGG-3′ and 5′-TCT GAC TTC TCT TCC GCT TTC TT-3′ for IFN-γ, 5′-GGA CTT CGA GCA GGA GAT GG-3′ and 5′-AGG AAG GAG GGC TGG AAG AG-3′ for β-actin. RT qPCR was performed on the Applied Biosystems 7500 Real Time PCR system (Applied Biosystems, Waltham, MA) using SYBR® Select Master Mix (Applied Biosystems) according to the manufacturer's manual. Briefly, a total reaction volume of 20 μL was utilized containing 10 μL SYBR Select Master Mix, 0.4 μL each primer (0.4 μmol/L), 2 μL cDNA, and 7.6 μL RNase-free dH2O. The cycling conditions consisted of initial denaturation at 95°C for 30 s, followed by 40 cycles at 95°C for 5 s and 60°C for 34 s. A final melting curve analysis was performed utilizing conditions of 95°C for 15 s, 60°C for 60 s, followed by 95°C for 15 s. All samples were performed in duplicate using β-actin as an internal reference gene. The relative expression of the objective genes compared to β-actin was determined as previously described (19).
Proliferation of PBMCs
PBMCs were labeled with 10 μmol/L carboxyfluorescein diacetate succinimidyl ester (CFSE) (Sigma-Aldrich) for 10 min at 37°C. CFSE-labeled PBMCs (5×104 cells/well) in 96-well plates were incubated in triplicate at 37°C in 5% CO2 for 5 days with stimulation by the addition of 10 μg/mL of concanavalin A (con A; Sigma-Aldrich) to RPMI-1640 culture medium (Sigma-Aldrich) containing 10% FBS, 4 mmol/L L-glutamine, 100 IU/mL penicillin, and 100 μg/mL streptomycin. The cells were harvested after 5 days in culture, and then washed twice with PBS. Proliferation of PBMCs was evaluated via dilution of CFSE on an Applied Biosystems Attune Acoustic Focusing Cytometer (Applied Biosystems). Data from 1×104 cells were analyzed using software version 1.2.5.
Statistical analysis
Data were analyzed by using the paired samples t-test and are presented as mean±standard deviation (SD) of the number of samples using IBM SPSS Statistics for Windows v19.0 (IBM Corp., Armonk, NY). p-Values of <0.05 were considered statistically significant.
Results
PCV2 infection in PMWS-affected pigs
Viral nucleic acids were evaluated by PCR in plasma samples from PMWS-affected and control groups. As shown in Figure 1, specific bands of 381 bp product for PCV2 were found to be amplified in the plasma (Lane 1, Fig. 1A), lung (Lane 1, Fig. 1B), and spleen (Lane 1, Fig. 1C) of PMWS-affected pigs, whereas no bands for PPV, PRV, PRRSV, and CSFV were observed in any of the samples. These data confirm our physiological observations of the PMWS-affected and uninfected groups.
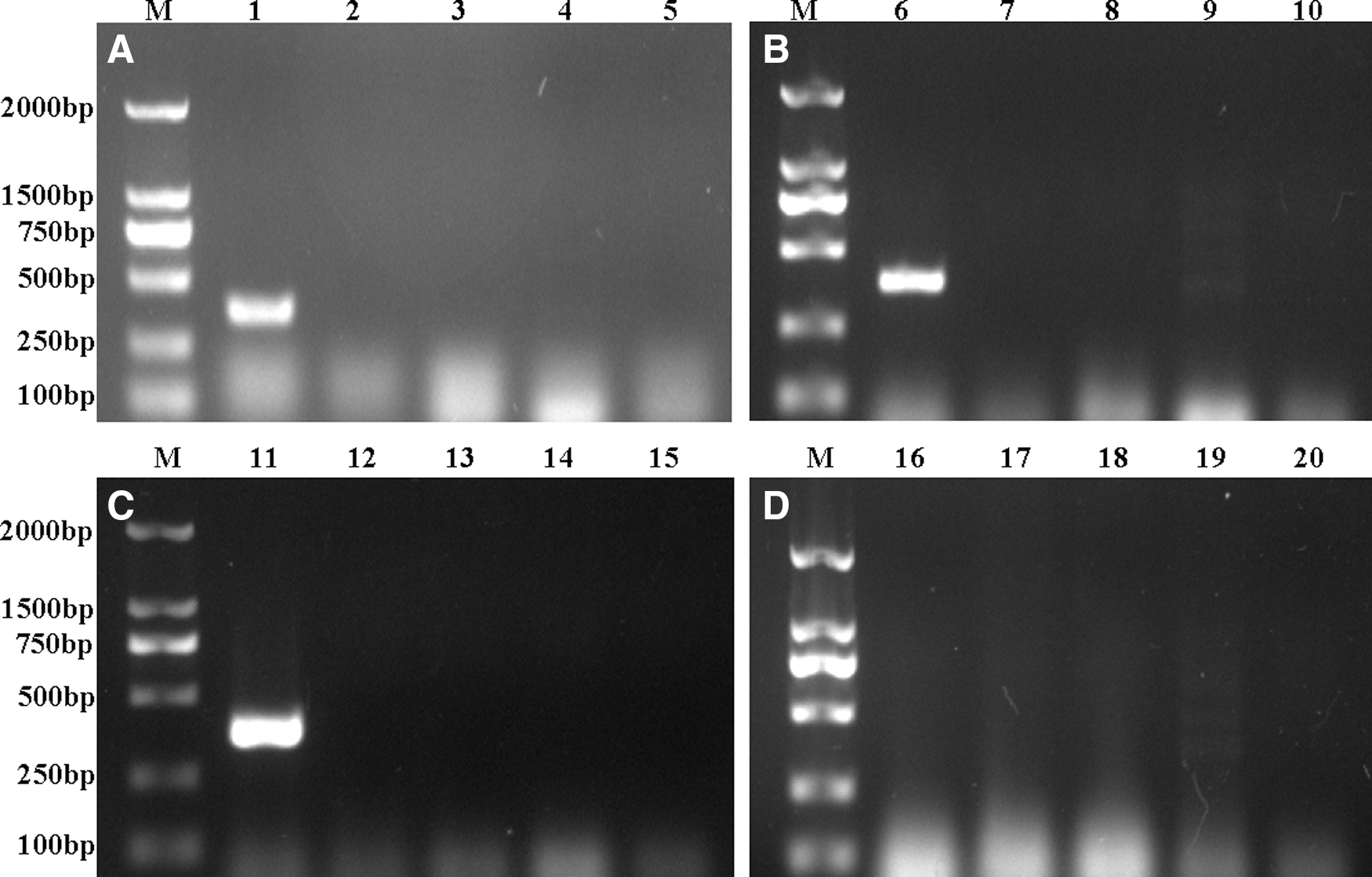
Pathogen detection by polymerase chain reaction (PCR). Viral genomes were extracted from plasma, lung, and spleen samples. cDNA was synthesized from total RNA, and PCR was used to detect suspicious viruses such as porcine circovirus type 2 (PCV2), pseudorabies virus (PRV), porcine parvovirus (PPV), porcine reproductive and respiratory syndrome virus (PRRSV), and classical swine fever virus (CSFV). Amplified fragments were detected by agarose gel electrophoresis. The specific fragment of PCV2 (381 bp) was amplified from plasma (
Association of PD-1 ligands with PMWS
Analysis of mRNA expression of porcine PD-1, PD-L1, and PD-L2 in PBMCs demonstrated a significant increase in PD-L1 (p<0.01) and PD-L2 (p<0.05) expression of PMWS-affected pigs compared with control pigs (Fig. 2). The relative level of PTEN mRNA was also notably elevated in PBMCs of PMWS-affected pigs compared with controls (p<0.01; Fig. 2). Interestingly, no difference in expression of PD-1, CTLA-4, LAG-3, or Foxp3 mRNA was found in PBMCs of pigs with PMWS compared to the uninfected pigs. These data suggest that some, but not all, immunoinhibitory processes are involved in PMWS.
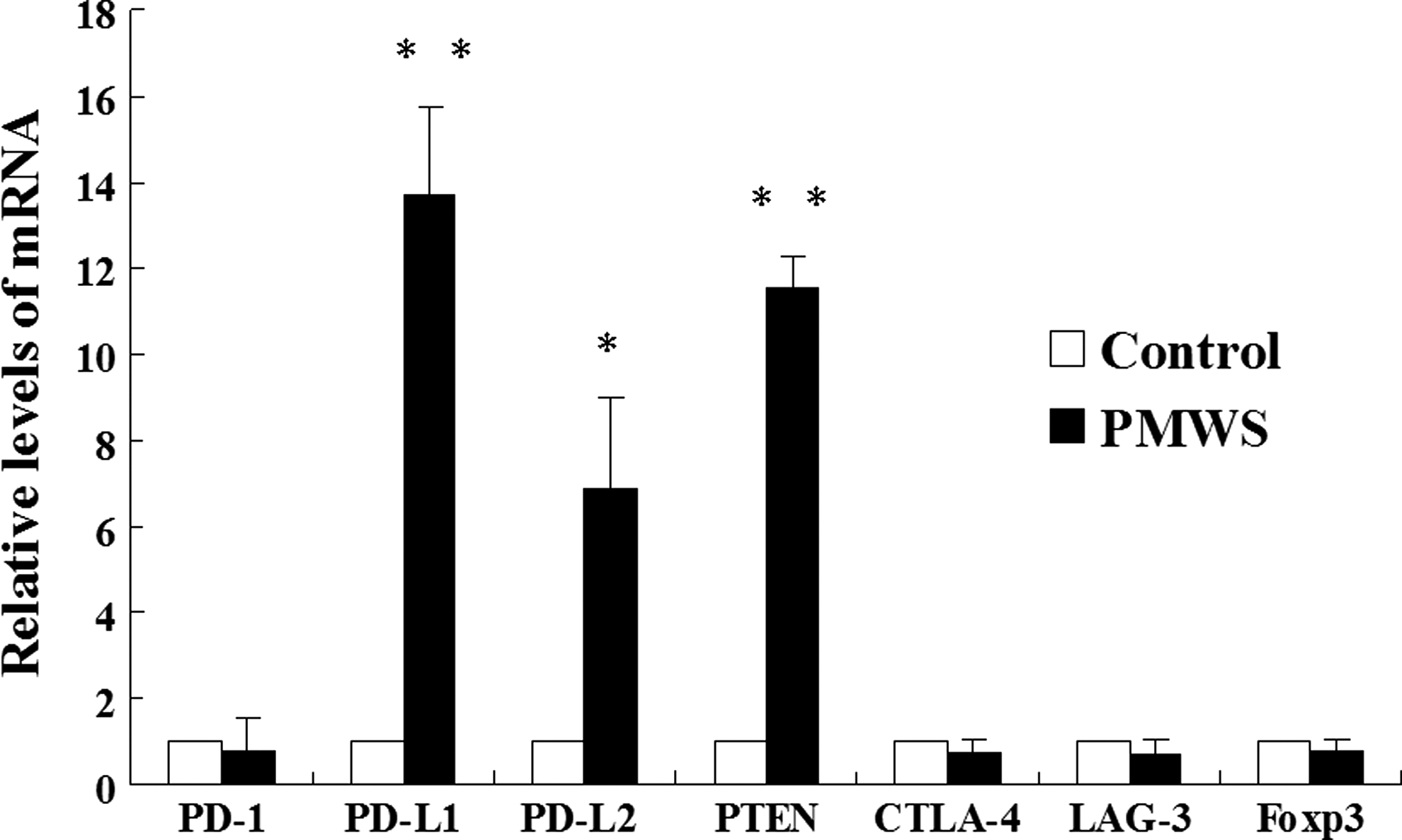
Transcriptional changes in PD-1, PD-L1, PD-L2, PTEN, CTLA-4, LAG-3, and Foxp3 levels in postweaning multisystemic wasting syndrome (PMWS)-affected pigs. Peripheral blood mononuclear cells (PBMCs) were isolated from healthy and PMWS-affected pigs, and total RNA was extracted from them. Residual DNA was removed from the total RNA samples, and first-strand cDNA was synthesized from 1 μg of total RNA. Results are expressed as relative fold changes of PD-1, PD-L1, PD-L2, PTEN CTLA-4, LAG-3, and Foxp3 mRNA using β-actin as an internal control. Values are shown as the mean±standard deviation (SD) from triplicate samples. Differences between groups were considered statistically significant at p<0.05 (*p<0.05; **p<0.01).
Cytokine expression in PMWS-affected PBMCs
To investigate the immunosuppressive phenotype further, expression of porcine IL-10, IL-2, and IFN-γ mRNA was analyzed using the PBMCs of PMWS-affected and control pigs. A slight increase in expression of IL-2 and IFN-γ was observed in the PBMCs of PMWS-affected pigs compared to controls (Fig. 3), although statistical significance was not reached. Interestingly, the relative level of IL-10 mRNA (p<0.01) was notably elevated in the PBMCs of pigs suffering from PMWS compared with uninfected pigs, which positively correlated with levels of PD-L1 and PD-L2.
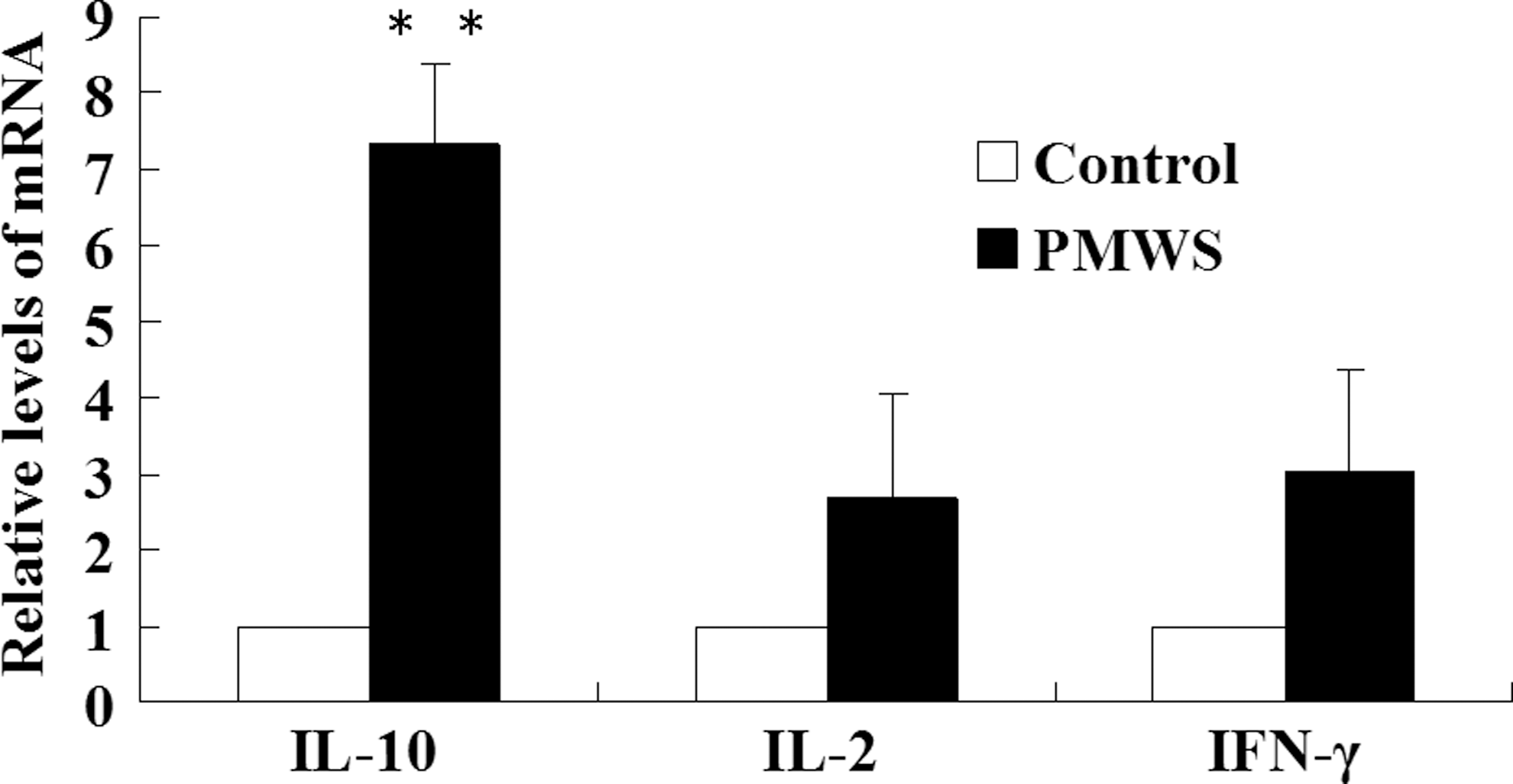
Transcriptional changes in IL-10, IL-2, and IFN-γ levels in PMWS-affected pigs. PBMCs were isolated from healthy and PMWS-affected pigs, and total RNA was extracted from them. Residual DNA was removed from the total RNA samples, and first-strand cDNA was synthesized from 1 μg of total RNA. Results are expressed as relative fold changes of IL-10, IL-2, and IFN-γ mRNA using β-actin as an internal control. Values are shown as the mean±SD from triplicate samples. Differences between groups were considered statistically significant at p<0.05 (**p<0.01).
Proliferation of PBMCs in PMWS-affected pigs
Reduced PBMCs cellularity is also a key characteristic of immunosuppression in PMWS-affected pigs. Considering the increase in antiproliferative proteins (PD-Ls and PTEN), we sought to determine if this had an effect on PBMC proliferation. CFSE was used to label cells, and the dilution of cellular label was detected by flow cytometry, providing a measure of mean fluorescence intensity (MFI) of the proliferating cells. In PMWS-affected pigs, the MFI of the PBMCs was notably decreased compared to that of controls (p<0.01; Fig. 4), suggesting that expansion of these cells is markedly reduced in pigs suffering from PMWS. Taken together, these data further confirm the apparent immunosuppressive phenotype in PMWS pigs and implicate both PD-L proteins and PTEN in the process.
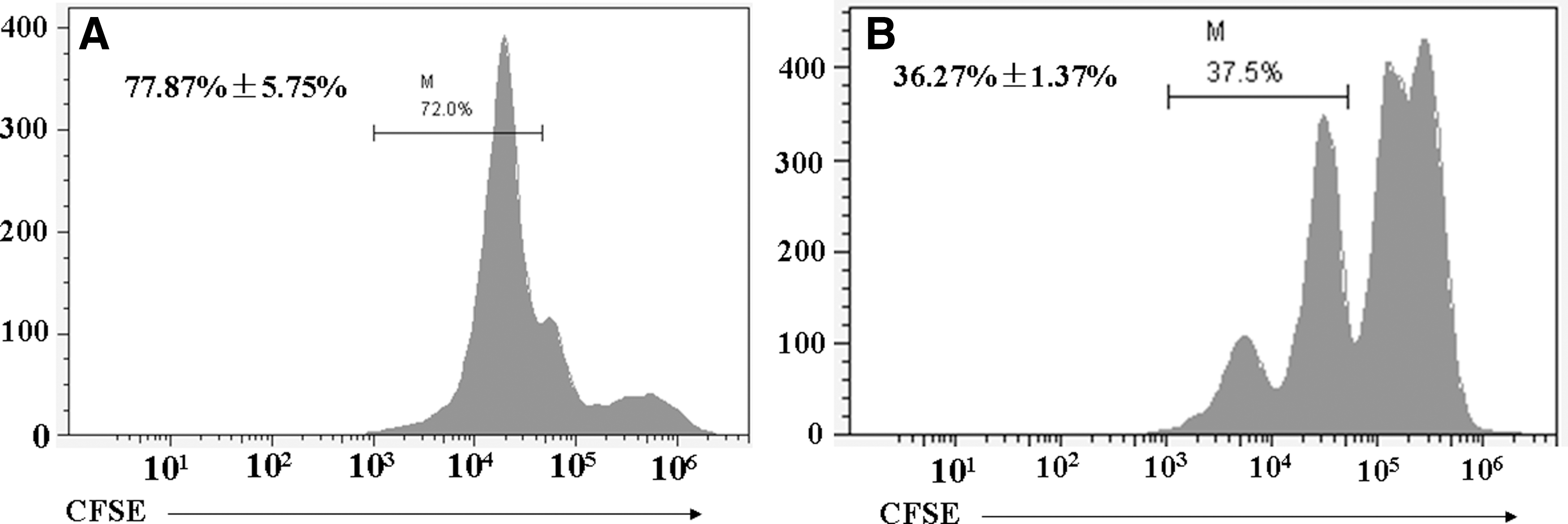
Impaired PBMC proliferation with pigs suffering PMWS. PBMCs were labeled with carboxyfluorescein diacetate succinimidyl ester and then stimulated by concanavalin A. Cells were harvested after culture for 5 days at 37°C 5% CO2 in RPMI-1640 culture medium supplemented with 10% fetal bovine serum. Cells were analyzed by flow cytometry. The mean fluorescence intensity of the proliferating PBMCs from diseased pigs
Discussion
Recently, the etiology of PMWS has been clarified through confirmation of PCV2 as the pathogen (1). Despite several studies demonstrating immunosuppression in PMWS-affected pigs characterized by leucopenia, impairment of lymphoid tissues, and imbalance of cytokines mRNA expression (7,20,21,23), much is still unknown about the pathogenesis of this disease. PD-1 is an immune inhibitory receptor that delivers negative signals upon interaction with its ligands (4). PD-1 and its ligands are key regulators of T- and B-cell activation and tolerance, followed by induction and maintenance of peripheral tolerance (10). To elucidate the immunopathological mechanisms in PMWS-affected pigs further, this study provides the first description of alterations in the PD-1 pathway in PBMCs in pigs naturally affected by PMWS.
Many studies have demonstrated that PD-1 and its ligands are selectively upregulated in exhausted T-cells during chronic viral infection, resulting in host immunosuppression (24). In this report, relative levels of PD-L1 and PD-L2 mRNA were remarkably increased in PBMCs of pigs suffering from PMWS, while no difference was observed in expression of PD-1 mRNA. Butte et al. demonstrated that B7-1 is an additional binding partner for PD-L1 but not for PD-L2. This PD-L1-specific interaction results in inhibition of T-cell activation (4). As every immune response requires co-stimulation through the B7-1/B7-2:CD28 co-stimulatory axis (5), it is proposed that the higher expression of PD-L1 mRNA in the PBMCs of PMWS pigs accelerates the interaction between B7-1 and PD-L1 to mediate B7-1:CD28 binding, resulting in inhibition of T-cell activation.
Another interesting finding concerns PTEN expression, which was found to be significantly augmented in the PBMCs of pigs suffering PMWS compared to controls. This is in accordance with previous studies showing that activation of the PD-1/PD-Ls pathway can result in augmentation of PTEN expression and decreased PIP3/AKT signaling, effectively resulting in attenuated T-cell activation (9). Other immunoinhibitory receptors, such as CTLA-4, LAG-3, and Foxp3, were not increased in the PBMCs of pigs suffering from PMWS compared to controls, suggesting that these receptors do not participate in the pathogenesis of PMWS.
Many studies have shown that PD-L1 and PD-L2 can inhibit T-cell proliferation and cytokine production (8,14), which are the main characteristics of T-cell activation. The current data show that expression of IL-2 and IFN-γ was not remarkably increased in PBMCs from PMWS pigs compared to controls, consistent with previous reports detailing cytokine mRNA expression profiles in lymphoid tissues of pigs naturally affected by PMWS (7). Expansion of PBMCs was also reduced, presumably due to impaired lymphocyte function through increased expression of PD-L1 and PD-L2.
In addition, expression of IL-10 was significantly upregulated in PBMCs of PMWS pigs, consistent with a previous study showing overexpression of IL-10 mRNA in PMWS-affected thymi (7). Interestingly, a positive correlation between PD-L1, IL-10, and immunosuppresion has been noted in other studies. Brooks et al. showed that persistent lymphocytic choriomeningitis virus (LCMV) infection in mice results in a significant upregulation of IL-10 by APCs, leading to impaired T-cell responses. Furthermore, therapeutic administration of an IL-10 blocking antibody restored T-cell function and eliminated viral infection (3). Taken together, these findings suggest that upregulation of IL-10 is involved in virally induced immunosuppression and could be a new therapeutic target in PWMS.
This study has demonstrated that expression of PD-L1 and PD-L2 is upregulated in the PBMCs of PMWS-affected pigs. Cytokine IL-10 mRNA levels were also markedly elevated, while expression of IL-2 and IFN-γ was not affected. Proliferation of PBMCs was also decreased, suggesting that upregulation of PD-Ls correlates positively with host immune inhibition in pigs affected by PMWS. Future study should explore how PD-Ls act to modulate immune adjustment in PMWS-affected pigs.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (No. 31272539; No. 31201877).
Author Disclosure Statement
No competing financial interests exist.