
Abstract
Avian infectious bronchitis virus (IBV), which is prevalent in many countries causing severe economic loss to the poultry industry, causes infectious bronchitis (IB) in birds. Recombinant single-chain variable fragments (scFvs) have been proven to effectively inhibit many viruses, both in vitro and in vivo, and they could be a potential diagnostic and therapeutic reagent to control IB. In this study, six anti-IBV chicken scFvs, ZL.10, ZL.64, ZL.78, ZL.80, ZL.138, and ZL.256, were obtained by screening random clones from an immune antibody library. An analysis of nucleotide sequences revealed that they represented distinctive genetic sequences and greatly varied in complementarity-determining region three of the heavy chain. Neutralization tests showed that ZL.10, which bound the S1 protein in western blots, inhibited the formation of syncytia in Vero cells 48 h post IBV infection and decreased the transcriptional level of nucleoprotein mRNA to 17.2%, while the other five scFvs, including ZL.78 and ZL.256, that bound the N protein did not. In conclusion, the results suggested that specific and neutralizing chicken scFvs against IBV, which can be safe and economical antibody reagents, can be produced in vitro through prokaryotic expression.
Introduction
A
Recombinant single-chain variable fragments (scFvs) are a new form of genetically engineered antibody that has only one variable region of the immunoglobulin heavy and light chains that are connected by a linker peptide. ScFvs retain the specificity of the original immunoglobulin despite removal of the constant regions, and they penetrate tissues more easily and exhibit low immunogenicity due to their small size (1). It has been shown that scFvs can specifically differentiate viruses in studies of immunological diagnoses (15,18,29). Moreover, scFvs can effectively neutralize, either in vitro or in vivo, many viruses, including foot and mouth disease virus, human immunodeficiency virus type 1, severe acute respiratory syndrome virus, hepatitis C virus, and hepatitis B virus (16,19,24,30,35). ScFvs can inhibit viral infection or replication by blocking the function of viral structural proteins or the proteins and enzymes that are vital to replication (2,5,34). All of these studies suggested that recombinant scFvs could be an effective diagnostic or therapeutic reagent for IBV infection. However, until now, most antivirus scFvs were generated from mammalian species and aimed to cure human disease, whereas few scFvs were from chickens.
Because of the peculiar mechanism of immunoglobulin gene diversification in birds, antibody repertoire in chicken can be accessed more readily than that of any mammals other than the camelids (6). Thus, the complete spectrum of rearranged variable fragments can be amplified by polymerase chain reaction (PCR) using only two pairs primers designed around the conserved regions flanking the unique functional VH and VL genes (1). Moreover, many conserved proteins that are not immunogenic in mammals yield high-titer antibodies when used to immunize chickens (8,10). Chicken scFvs have been previously generated against bluetongue virus, the SARS-CoV spike protein, prion proteins, and infectious bursal disease virus (7,17,23,29).
Although anti-IBV chicken scFvs have been studied previously, only several antinucleoprotein scFvs have been obtained, and their functional, biological activities, such as neutralization, have not been studied either in vivo or in vitro. In our study, an immune expression library was constructed using a prokaryotic expression vector to screen anti-IBV scFvs, and their neutralizing activity in Vero cells was further analyzed.
Materials and Methods
Viral strains, vector and vaccine
The IBV M41 strain was provided by Professor Jianhe Sun at Shanghai Jiaotong University, China. Strain M41 was propagated for 9–10 days in embryonated chicken eggs, and purified by sucrose density gradient centrifugation.
Chicken immunization
Two 4-week-old female white leghorns were immunized with IBV M41 vaccines (Merial, Inc., Milan, Italy) according to the manufacturer's instructions. Three immunizations were conducted at intervals of 2 weeks until enzyme-linked immunosorbent assay (ELISA) antibody titers reached 32,000. Chickens were then euthanized, and the spleens were harvested immediately. Peripheral blood lymphocytes (PBLs) were isolated from anticoagulated blood using hydroxypropyl methylcellulose (Huajing, Shanghai, China) according to the manufacturer's instructions.
Construction of expression plasmids
RNA was extracted from 50 mg of spleen tissue and 3×109 PBL cells using the TRIzol reagent (Invitrogen, Carlsbad, CA), and 20 μg of pooled RNA was reverse transcribed to synthesize the first-strand cDNA using an MMLV Reverse Transcription Kit (Takara, Kyoto, Japan). ScFvs antibodies were constructed according to published protocols, with prokaryotic expression vector pOPE101 (1). Briefly, VH and VL genes were amplified from cDNA by PCR using two pairs of chicken-specific primers, and joined to form scFvs gene using a (Gly4Ser)3 linker by overlap extension PCR. The scFvs gene was then cloned into vector pOPE101 (Progen Biotechnik, Heidelberg, Germany) using the NotI and NcoI restriction sites. The correct expression vector was identified by PCR, restriction enzyme digestion, and sequencing. The recombinant vector pOPE101-scFv was electroporated into Escherichia coli strain JM109 (Takara) for expression. Positive transformants were identified by digesting plasmid DNA with the appropriate restriction endonucleases.
Expression and extraction of soluble scFvs
The transformed clones were inoculated into 2 mL of Luria-Bertani (LB) medium containing 100 μg/mL ampicillin, and cultured at 37°C overnight. Then, 20 μL of culture medium was removed and added to 2 mL of fresh medium and cultured at 37°C with shaking at 250 rpm to an optical density 600 (OD600) of 0.6–0.8. The induction was started by the addition of 50 μM isopropyl-β-D-thiogalactopyranoside (IPTG; Sigma-Aldrich, St. Louis, MO), followed by shaking at 250 rpm for 6 h at 30°C. The bacteria were then harvested by centrifugation at 4,000 g for 10 min at room temperature.
Periplasmic proteins of E. coli were extracted using the osmotic shock method as previously described, with minor modifications (7). Briefly, cell pellets were resuspended in 500 μL of TES buffer, pH 8 (500 mM sucrose, 30 mM Tris, 1 mM EDTA) at room temperature, with brief vortexing every 5 min. The bacteria were then pelleted for 10 min at 12,000 g at 4°C. The supernatant was discarded and the pellet was resuspended in 300 μL of ice-cold 5 mM MgSO4 and incubated for 20 min on ice, with brief vortexing every 5 min. The solution was then centrifuged for 30 min at 12,000 g and 4°C, and the supernatant was collected for further analysis.
Screening of clones
To examine the ability of scFvs to bind IBV, the periplasmic extractions of random clones were analyzed in an indirect ELISA as follows. Two hundred nanograms of IBV M41 was coated in sodium bicarbonate buffer (pH 9.6) in a 96-well plate at 4°C overnight. After coating, 100 μL of periplasmic extractions of each clone were added and reacted at 37°C for 2 h. The plates were then probed with a myc-tagged mouse mAb (1:2,000 dilution; RayBiotech, Norcross, GA) at 37°C for 1 h, followed by a horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (1:5,000 dilution; Dingguo, Beijing, China) at 37°C for 1 h. The plates were washed three times with phosphate-buffered saline-Tween (PBST) (0.5% [v/v] Tween-20) after each step. One hundred microliters of 3,3′,5,5′-tetramethylbenzidine (TMB) (HRP)-substrate (Tiangen, Beijing, China) was used for detection, and 50 μL of H2SO4 (2 M) was used to stop the reaction. The absorbance, which was proportional to the concentration of free anti-IBV scFvs in each well, was measured at 450 nm.
Sequence analysis
Positive clones were cultured in LB/ampicillin broth overnight at 37°C with shaking at 250 rpm. Plasmids were extracted from bacterial pellets using the AxyPrep Plasmid MiniPrep Kit (Axygen Biosciences, Hangzhou, China), and were then sequenced using forward 5′-CTATTGCCTACGGCAGC-3′ and reverse 5′-AAGGCCCAGTCTTTCGAC-3′ primers by Sangon Biotech Co., Ltd. (Shanghai, China). The deduced amino acid sequences were aligned with the germline gene form chicken.
Purification of scFvs
ScFvs were purified from E. coli product nickel affinity chromatography (Yihong, Shanghai, China) according to the manufacturer's instructions. Briefly, the nickel column was equilibrated using buffer (20 mM Na2HPO4, 0.5 M NaCl), and nonspecific proteins were eluted using one application of 10 mM imidazole-containing buffer (20 mM Na2HPO4, 0.5 M NaCl, 10 mM imidazole), followed by three washes with 50 mM imidazole buffer. The six-histidine-tagged scFvs were eluted using phosphate-buffered saline (PBS), pH 7.4, containing 100 mM EDTA. After dialysis against PBS buffer, scFvs proteins were concentrated to 0.1–0.3 mg/mL by ultrafiltration using Centricon YM-10 concentrators (Millipore, Bedford, MA).
Western blotting
The scFv-expressing lysates, periplasmic extractions, and purified scFvs were subjected to sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) to analyze protein content. Periplasm extracted from a clone containing the pOPE101 vector was used as a negative control. In addition, the binding of scFvs to IBV was further analyzed by western blotting. Briefly, 1 μg of purified IBV particles were respectively subjected to 12% (w/v) SDS-PAGE, and then transferred onto a nitrocellulose membrane, and incubated with 500 ng of purified scFvs at 37°C for 2 h. Binding was indirectly detected through a myc-tagged mouse mAb (1:2,000 dilution) and HRP-conjugated goat anti-mouse IgG (1:4,000 dilution). The bands were exposed to X-ray films and visualized using the SuperSignal West Pico Trial Kit (Pierce, Rockford, IL). Anti-IBV polyclonal serum was used as a positive control, but in this case, binding was detected through HRP-conjugated donkey anti-chicken IgG (1:4,000 dilution).
Neutralization of scFvs
The Vero cell line was cultured in Dulbecco's modified Eagle's medium (DMEM) containing 10% (v/v) fetal bovine serum and 1% (w/v) penicillin and streptomycin. Before use, the allantoic fluid of IBV M41 was diluted with DMEM, filtered through a 0.22-μm microfilter, and treated with 1% penicillin and streptomycin for 30 min. A 100 μL serial dilution of IBV M41 was added to a 96-well plate precultured with 0.5×105 Vero cells in each well. After 3 days of culturing, the viral 50% tissue culture infectious dose (TCID50) was calculated using the Reed and Muench method. Subsequently, 1 μg of each scFv was incubated with 100 TCID50 of IBV M41 for 1 h at 37°C. The antibody–virus mixture was then added to six-well tissue culture plates prepared with 1×106 Vero cells/well, and incubated at 37°C in 5% (v/v) CO2 for 30 min, and the virus was then replaced with fresh growth medium. Immune sera were used as positive controls, and an scFv that did not react with IBV in an ELISA was used as a negative control. The cytopathic effect was observed every 12 h. After 48 h of infection, RNA was extracted from the cells for real-time quantitative PCR (RT-qPCR) analysis.
Real-time quantitative PCR
The total RNA from each cell sample was extracted from six-well plates using the TRIzol reagent (Invitrogen). RNA was previously treated with DNAse I (Takara) for 20 min. RNA integrity and quality were tested by Experion™ Automatic electrophoresis system. RNA was reverse transcribed into cDNA using MMLV reverse transcriptase and oligo(dT) primers. Forward and reverse primers and a TaqMan® probe were designed as previously described to detect the nuclear protein of IBV (22). The reaction was conducted in a PTC-200 DNA Engine thermal cycler (Bio-Rad, Hercules, CA) at 95°C for 10 min, and 40 cycles of 95°C for 15 sec, and 60°C for 35 sec, in 25 μL volumes containing Premix Ex Taq™ (2×, 12.5 μL), primers (10 μM, 1 μL), TaqMan probe (10 μM, 1 μL), cDNA template (2 μL), and nuclease-free water (7.5 μL). Amplification plots were recorded, analyzed, and the quantification cycle (Cq) determined with Opticon Monitor software, version 3 (Applied Biosystems, Foster City, CA). All procedures were performed in triplicate and the mean values were calculated. The percentage of IBV nucleoprotein mRNA expression was analyzed in comparison with the infected cells (normal control).
Results
Construction of the expression plasmid
VH and VL gene fragments were amplified from cDNA from immunized chicken spleens, and were then combined to form an scFv through a 45-base pair (bp) linker sequence by overlap extension PCR. The molecular sizes of the VH, VL, and scFv genes were ∼400, 350, and 750 bp, respectively, in agarose gel electrophoresis. The scFv gene was cloned into the pOPE101 vector, and the recombinant plasmid pOPE101-scFv was then transformed into JM109 E. coli cells for expression. The combined library size, which was calculated according to the total number of independent transformants, was 3.6×106. The proportion of self-ligation of the vector was less than 5%, and the sequences of 25 randomly chosen clones were unique.
Production of soluble antibody
The optimal conditions, such as temperature, host, and culture media, for scFv expression using this vector were investigated by Hust (12). ScFvs are both expressed in the cytoplasm and periplasm after induction, which are soluble forms in the periplasm. The molecular mass of antibody fragments, as confirmed by SDS-PAGE, was 31 kDa (Fig. 1), which corresponded with the predicted molecular weights. Expressing at a low temperature (25°C) and a low concentration of IPTG (50 μM) were helpful for periplasmic-expressing scFvs.
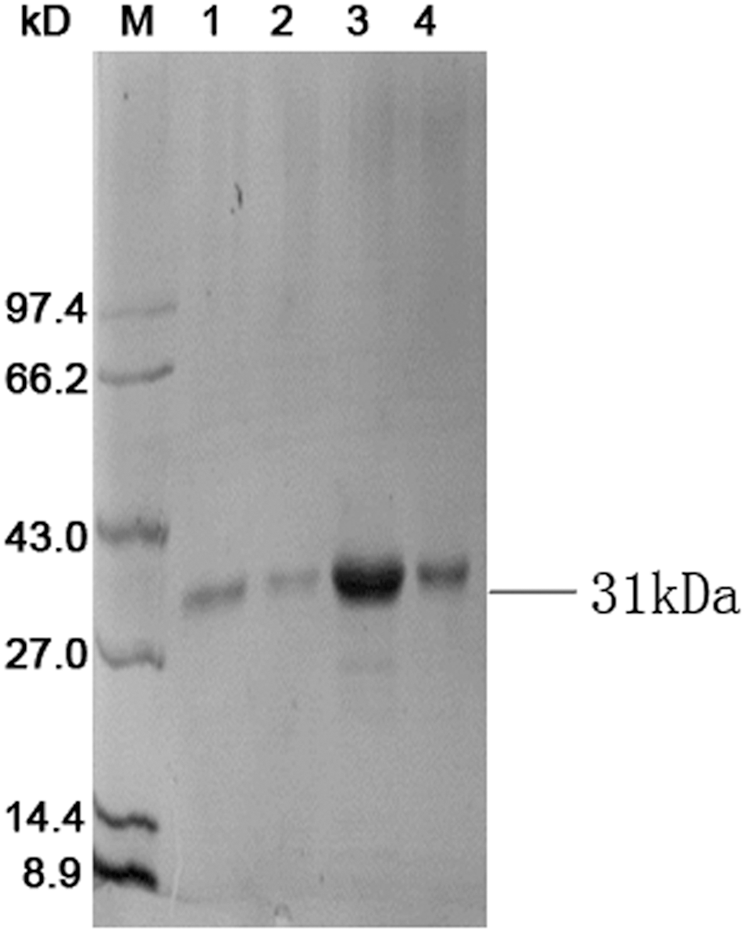
Expression of pOPE101-scFv. Sodium dodecyl sulfate–polyacrylamide gel electrophoresis analysis of the purified single-chain variable fragment (scFv). The gel was stained by Coomassie brilliant blue. M, Protein ladders; lane 1–4, purified scFv.
Screening and sequencing
Soluble scFvs were extracted from the periplasm by the osmotic shock method and analyzed for their ability to bind IBV by indirect ELISA. After screening of 800 clones, six clones (ZL.10, ZL.64, ZL.78, ZL.80, ZL.138, and ZL.256) were identified as positive (Fig. 2). The amino acid sequences, deduced from nucleotide sequences of these six clones, were aligned with the germline sequence from chicken. The frequency of changed amino acids of these six clones was similar to those in natural antibodies. Most of the changes were in the complementarity-determining regions (CDRs), but more changes were observed in CDR H3 (Fig. 3). All these scFvs contained amino acid insertions, ranging from 5 to 11 residues, in CDR H3, while ZL.10, ZL.138, and ZL.256 exhibited deletions of two amino acids. These results showed that the sequence differences were predominantly localized in CDRs and nucleotide insertions only occurred in CDR H3.
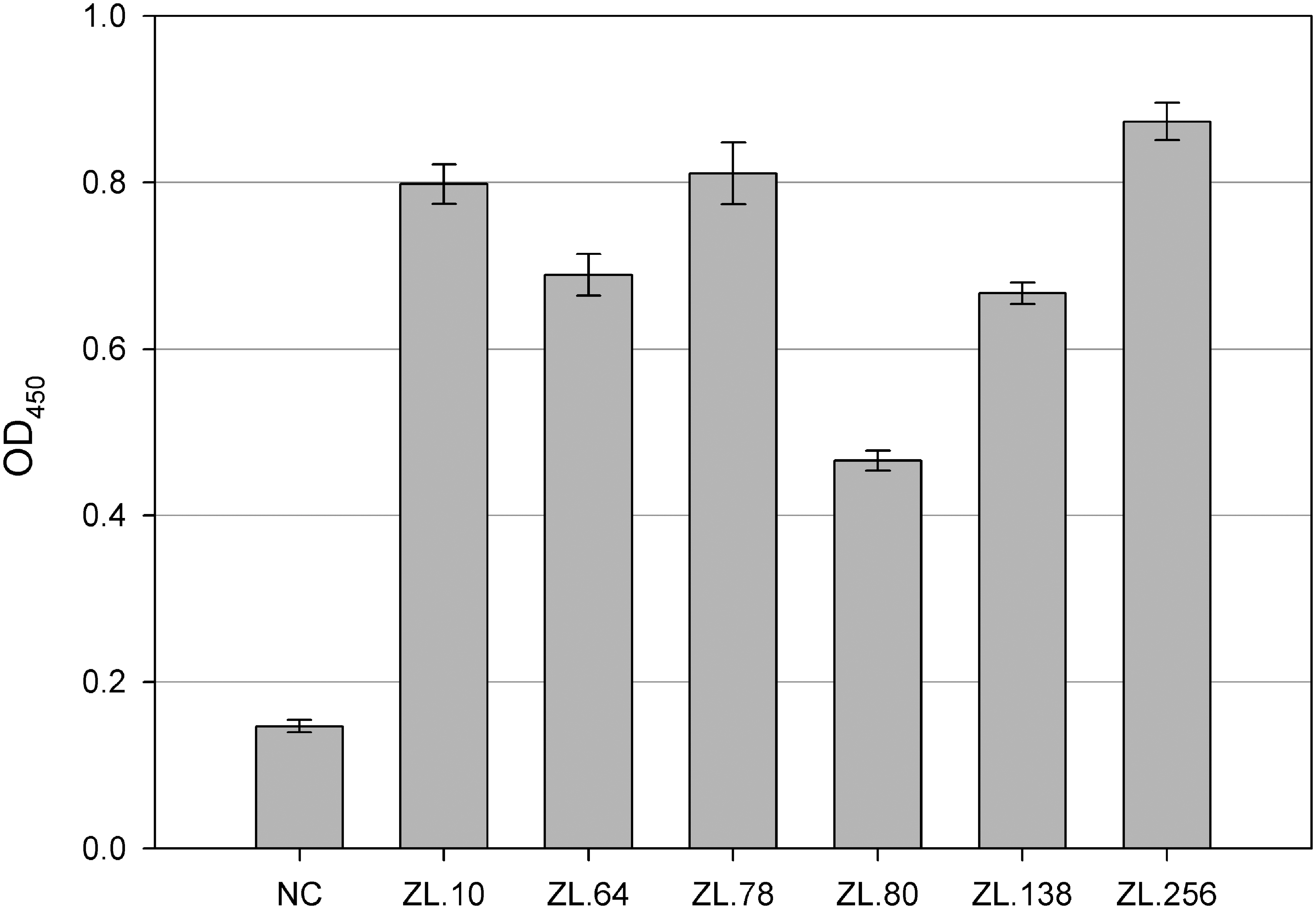
Enzyme-linked immunosorbent assay (ELISA) reaction of anti-IBV scFvs. Two hundred nanograms of avian infectious bronchitis virus (IBV) was coated at 4°C overnight and 100 μL of periplasmic extractions of each clone were reacted at 37°C for 2 h. A myc-tag mouse mAb and horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG were subsequently used for detection. NC is the negative control, which had an optical density 450 (OD450) below 0.2. Six clones (ZL.10, ZL.64, ZL.78, ZL.80, ZL.138, and ZL.256) screened from the antibody library reacted with IBV, which was evidenced by an OD450 above 0.4. The data represent the average of three experiments, with the standard deviation indicated by error bars.

Amino acid sequence and nucleotide sequence alignment of anti-IBV scFvs. Heavy-chain variable (VH) and light-chain variable (VL) region sequences were compared with that of the chicken germline sequences, as shown in
Antigen binding
The results of western blotting showed that immune serum identified three structural proteins of IBV, including the spike1 (S1), nucleocapsid (N), and membrane (M) proteins, and their molecular masses were 92, 52, and 37 kDa, respectively. Moreover, the results showed that ZL.10 and ZL.80 reacted with the S1 protein, while ZL.78 and ZL.256 reacted with the N protein. However, ZL.64 and ZL.138, like the negative control, did not react with any structural proteins of IBV (Fig. 4). ZL.64 and ZL.138 may only react with conformational epitopes, but not linear epitopes.
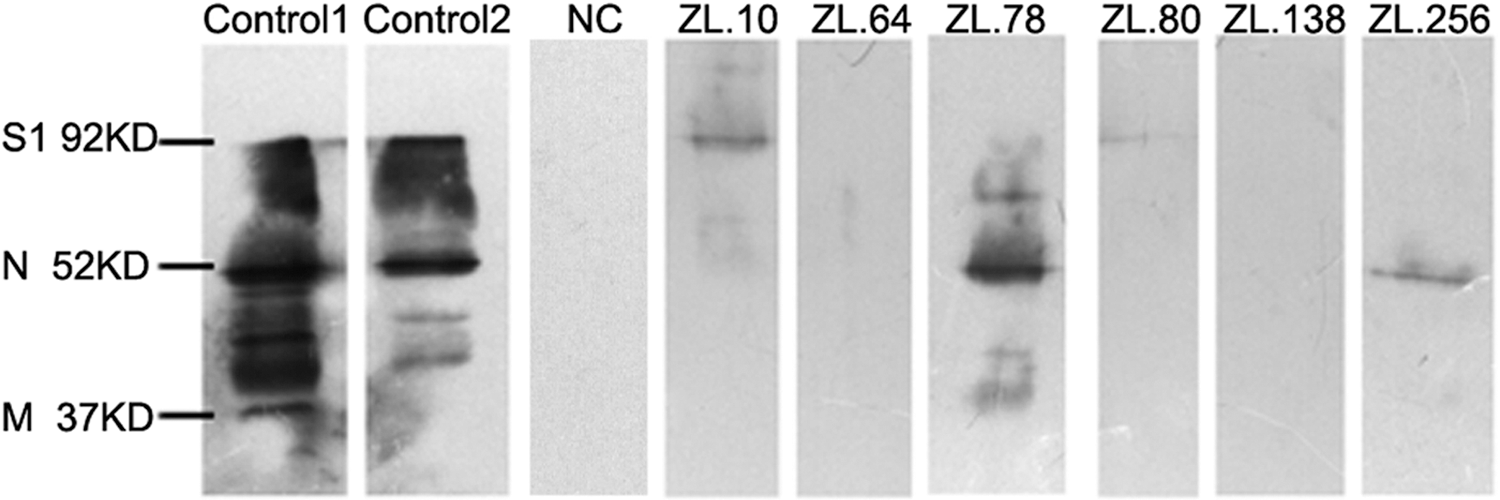
Western blotting analysis of the IBV-binding ability of scFvs. Immune serum and six anti-IBV scFvs were incubated with purified IBV to analyze their binding activity. Control 1 and Control 2 were incubated with serum1 and serum 2, which were isolated from two immunized chickens. NC is the negative control.
Neutralizing activity
IBV M41 was propagated in Vero cells, and syncytia occurred at 36 h postinfection. The TCID50, which was calculated according to the Reed and Muench method, was 103.5. The result of neutralizing activity showed that ZL.10 effectively inhibited the infection, as only a few syncytia were observed in cells at 48 h postinfection, while immune serum completely inhibited the infection, as no syncytia were observed at 48 h postinfection. However, the other five scFvs, as well as the negative control, showed no neutralizing activity (Fig. 5).
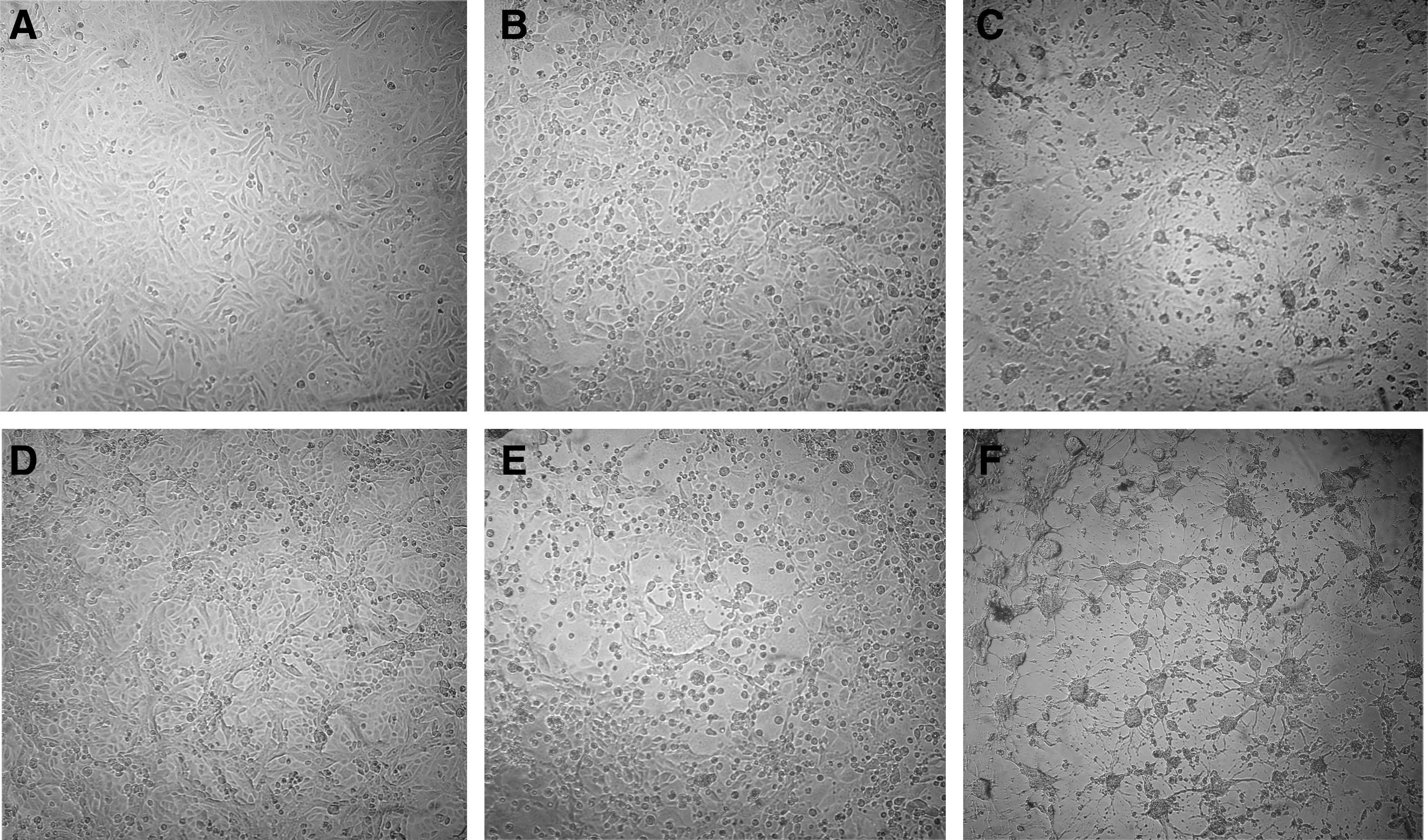
Inhibition of the cytopathic effect by ZL.10.
The relative transcription level of IBV nucleoprotein mRNA was further analyzed by RT-qPCR and calculated according to the Pfaffl method (26). The transcription level of nucleoprotein mRNA decreased to 5.3% and 17.2% in cells infected with IBV pretreated with immune serum and ZL.10, respectively. However, no significant changes were noted in cells infected with IBV pretreated with the other scFvs or the negative control (Fig. 6). All these results illustrated that ZL.10 can partially inhibit the infection of Vero cells by IBV.
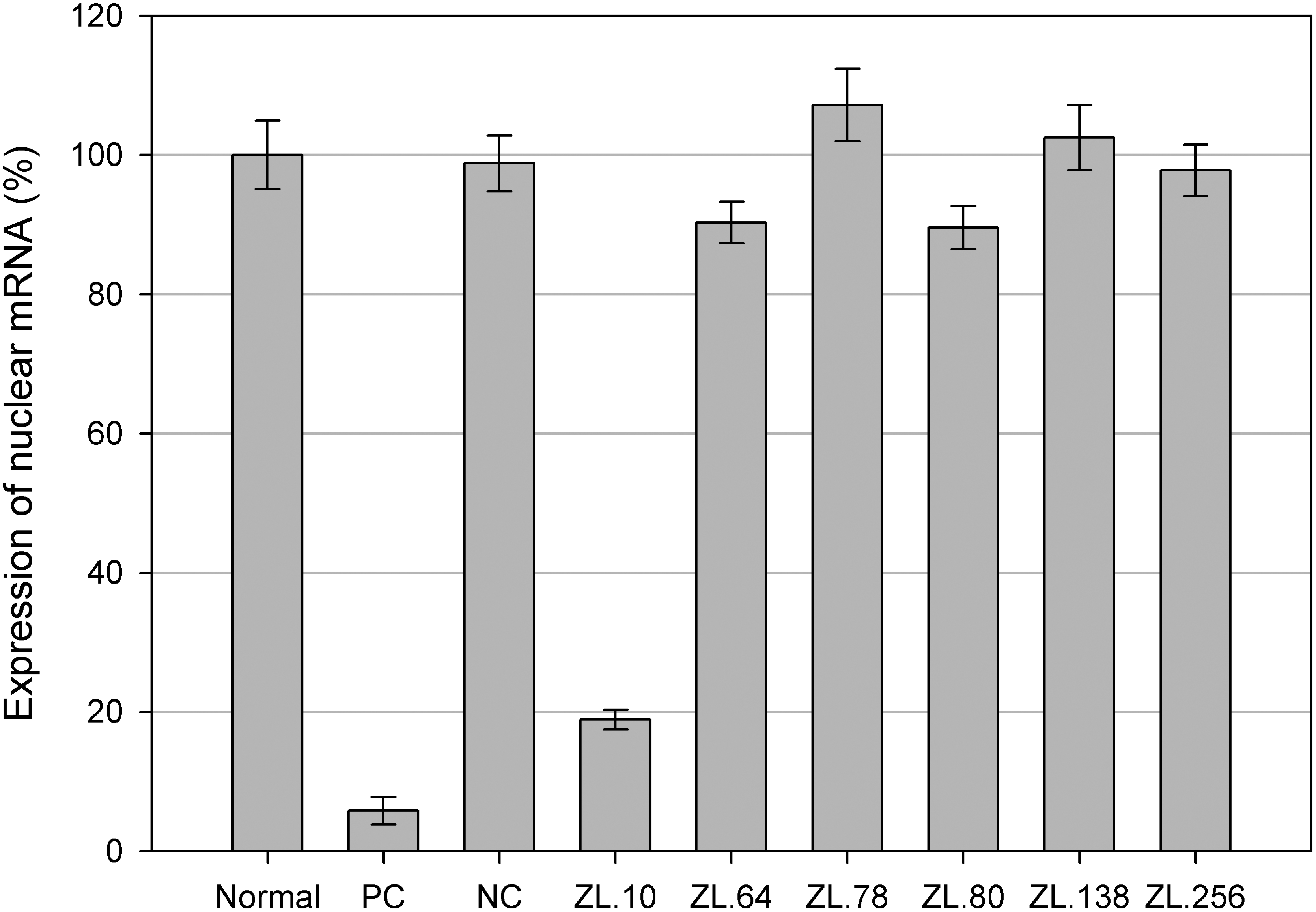
Expression of IBV nucleoprotein mRNA in groups preincubated with or without scFvs. The expression of IBV nucleoprotein mRNA was detected by real-time quantitative PCR (RT-qPCR) 48 h post-IBV infection in each group. It was decreased by pretreatment with ZL.10, as well as by an immunized serum control (PC), but no significant changes were noted for the other five anti-IBV scFvs and the scFv-negative control (NC), or in the group without pretreatment (Normal). All assays were performed in triplicate and data are expressed as means. Error bars indicate standard deviations.
Discussion
Monoclonal chicken scFv antibodies can help to develop valuable reagents for both scientific and clinical applications, while passive administration of neutralizing monoclonal antibodies can be used for emergency prophylaxis and treatment of viral disease. The neutralizing recombinant anti-IBV chicken scFvs obtained in this study illustrate the practical utility of an expression antibody library for producing diagnostic or potentially therapeutic reagents. The results further showed that an scFv against the S1 protein can specifically inhibit viral replication as well as immunized serum in vitro. Neutralizing anti-IBV chicken scFvs were generated from an expression antibody library, and these scFvs could serve as efficacious diagnostic and therapeutic reagents to control IBV.
Immune libraries derived from immunized or convalescent animals contain abundant and mature antibodies; thus, they are useful for screening for functional antibodies (9,21). Therefore, we constructed immune libraries using the prokaryotic expression vector pOPE101, and the scFv gene that was derived from the spleens and PBLs of immunized chickens. Six anti-IBV scFvs were obtained by screening 800 random clones from this antibody library, thereby demonstrating a relatively high ratio of positive clones in the library. Nucleotide sequence analysis showed that the CDRs exhibited a high degree of sequence diversity among the different clones, indicating the importance of CDRs in antigen recognition (10). The increased amino acid length in CDR H3 of these six scFvs, in comparison with the germline sequence, was great, ranging from 6 to 19 amino acids, which may be due to the large differences of gene, caused by VDJ rearrangement and variations in the D gene during gene formation process (28). However, the changed amino acid length in CDR L3 was few, as only two amino acids were deleted in ZL.10, ZL.138, and ZL.256, which may be due to the lack of D elements in the formation of the VL gene. Moreover, the variety of CDR H3 and CDR L3 may be due to conversion with an upstream pseudogene and somatic mutation induced by antigen (1,14,27). These results are consistent with the conclusion that CDR3 H3 is particularly important in determining antigenic specificity. There were also invariant amino acids in the individual CDRs of each scFv, which were often considered as conserved and important for structural integrity (1).
The neutralization tests in our study showed that only the binding of ZL.10 to the S1 glycoprotein effectively inhibited IBV as well as the positive serum, which was consistent with a previous study that showed that the S1 glycoprotein contains serotype-specific neutralization epitopes (25). In contrast, the other five scFvs, including ZL.64 and ZL.78 that bound to the N protein, have no apparent neutralization, which was in accordance with a previous conclusion that the S protein, but not other structural proteins, is associated with humoral immunity and induces protection in vaccinated chickens (13,31). The additional bands recognized by the clone ZL.78 in Figure 4 is due to the cross reaction that happened between ZL.80 and the nonstructural proteins of IBV. Many kinds of mechanisms have been proved for the neutralization activity of antiviral antibodies, including virus aggregation, masking of receptor attachment sites, steric inhibition of receptor binding, or stabilization of the capsid to inhibit virus uncoating (34). Moreover, it has been shown that intact antibodies, neutralizing or even non-neutralizing, protect animals against lethal infection because they can lyse virus-infected cells by antibody-dependent cellular cytotoxicity (ADCC) (4). However, scFvs have no ADCC function, as they lack the Fc, and their neutralization ability in vivo was, therefore, directly correlated with their neutralization ability in vitro (32), which is convenient for the screening of functional scFvs.
In summary, the present study demonstrated that functional anti-IBV chicken scFvs could be obtained by screening an expression antibody library, and that they can be produced safely and economically in a bacterial host (11). Neutralization tests showed that binding of ZL.10 to the S1 protein effectively decreased the replication of IBV in Vero cells, while the binding of scFvs to other IBV proteins did not. These results illustrated that the inhibition of IBV infection by scFvs may be due to the blocking of IBV binding to the receptor of Vero cells (31,33). Because scFvs in vivo may be affected by other neutralizing mechanisms, such as binding to the N or M protein that inhibits the replication of IBV (3), their neutralization activity still needs to be validated in vivo. The development of scFvs may provide a new class of potent and selective anti-IBV agents for diagnosing and curing IB.
Footnotes
Acknowledgments
Funding: This work was funded by the project “Development of a chicken antibody library and screening for a single-chain variable fragment derived from immunization against a main pathogen,” which was supported by the Shanghai Key Laboratory of Veterinary Biology, and by a grant XT2012010 from Ningxia Medical University. The authors thank Professor Jianhe Sun at Shanghai Jiaotong University for the donation of IBV M41.
Author Disclosure Statement
No competing financial interests exist.