
Abstract
Hepatitis C virus (HCV) pathogenesis and treatment outcomes are multifactorial phenomena involving both viral and host factors. This study was designed to determine the role of tumor necrosis factor–related apoptosis-inducing ligand receptor 1(TRAIL-R1) and interferon gamma (IFN-γ) genetic mutations in susceptibility and response to interferon-based therapy of hepatitis C virus (HCV) infection. The detection of TRAIL-R1 rs4242392 and IFN-γ rs2069707 single nucleotide polymorphisms was completed in 118 chronic HCV patients and 96 healthy controls by allele-specific polymerase chain reaction and restriction fragment length polymorphisms polymerase chain reaction. Patients were further categorized into sustained virological responder (SVR) and nonresponder (NR) groups on the basis of their response to interferon-based therapy for HCV infection. Real-time PCR was used for HCV quantification. HCV genotyping was performed by Ohno's method. The results demonstrated that the distribution of the TRAIL-R1 rs4242392TT genotype was significantly higher in the SVR group (78%) compared to the NR group (36%). It showed that chronic HCV patients possessing the TRAIL-R1 rs4242392TT genotype are better responders to interferon-based therapy (p<0.05). The prevalence of the TRAIL-R1 rs4242392TT genotype in healthy controls and chronic HCV patients was 56% and 65% respectively. It indicated that there is the TRAIL-R1 rs4242392 genetic variation plays no role in the spontaneous clearance of HCV infection (p>0.05). The distribution of IFN-γ rs2069707 was the opposite to TRAIL-R1 rs4242392 prevalence, that is, there was high distribution of the IFN-γ rs2069707GG genotype in patients and healthy controls (p<0.05), while the prevalence of IFN-γ rs2069707GG in SVR and NR groups was comparable (p>0.05). In conclusion, genetic variation of TRAIL-R1 rs4242392 is linked with response to interferon-based therapy for HCV infection, and genetic variation IFN-γ rs2069707 is associated with natural clearance of HCV infection.
Introduction
H
Cells of both the innate and adaptive immune system, such as macrophages, natural killer (NK) cells, B-cells, and T-cells, show expression of the tumor necrosis factor–related apoptosis-inducing ligand (TRAIL) gene in response to IFN-β and IFN-γ (23,43). The TRAIL gene, like the tumor necrosis factor (TNF) family, is involved in the modification of the immune system (19). TRAIL receptor 1, also known as death receptor 4 (DR-4), is a transmembrane protein of 468 amino acids (31). It has been shown in various studies that the TRAIL-R1 rs4242392 polymorphism is involved in the cancer etiology of different organs, including the breast, colon, and stomach (25). In a recent clinical study of HCV/human immunodeficiency virus (HIV) co-infected patients, it was demonstrated that the TRAIL-R1 rs4242392 genetic variation is significantly correlated with a lack of treatment response (33).
IFN-γ is a glycoprotein containing a single polypeptide component of 143 amino acids, and exists in mono- and biglycosilate forms in humans (39). IFN-γ is essentially required for T helper 1 (Th1) immune responses. It enhances antigen presentation as well as the expression of co-stimulatory molecules (34). IFN-γ is potentially used as an immune-modulatory medicine due to its pleiotropic activity. In a clinical study, it was shown that expression-level IFN-γ is linked with liver pathogenesis and lack of treatment response in chronic HCV-infected patients (12). Additionally, it was shown that IFN-γ produced by effector T-cells and NK cells inhibits HCV replication in a replicon system (16). Like other cytokines, IFN-γ mostly possesses important SNPs in its noncoding region. A genetic variation of IFN-γ positioned at +874 (rs2430561) in the intron 1 region is linked with a high production level of IFN-γ (27), and early recurrence of HCV infection in liver-transplanted patients (4).
The inter-individual differences in disease progression and treatment outcomes may be attributed to SNPs in the human genome (36). A better understanding of host genetics may allow the development of individualized therapy leading to increased cure rates and may license innovative therapeutic approaches. The current efforts were focused on finding the correlation of genetic mutations of TRAIL-R1 rs4242392 and IFN-γ rs2069707 with natural clearance and treatment response of HCV infection in a Pakistani population.
Methods
Study population
The current study was approved by the research ethics committee of the institute. All studied subjects provided written informed consent for genetic testing of their blood samples. A total of 214 individuals were recruited. The blood samples of 65 chronic HCV-infected patients were collected from the Gambat Institute of Medical Sciences (Sindh), while the blood samples of 53 chronic HCV-infected patients were collected from the Saidu Teaching Hospital (Khyber Pakhtunkhwa). The inclusion criteria were the detection of HCV-RNA by real-time polymerase chain reaction (RT-PCR). Thus, collectively, 118 chronic HCV patients (from November 2010 to April 2013) were identified with any final fate of therapy. Enrolled patients were divided into the sustained virological responder (SVR) group and the nonresponder (NR) group based on treatment response. SVR patients were defined as those subjects who were negative for HCV RNA by RT-PCR 6 months after the end of therapy. The blood samples of 96 healthy individuals were taken as control samples.
SNPs detection by allele-specific polymerase chain reaction
Genomic DNA was extracted from the whole blood of all studied subjects using a DNA extraction and purification kit (catalog no: 1820-02; Invitrogen) in accordance with the provided protocol. The DNA concentration for the PCR was kept at 40–100 ng/μL.
Both of the studied SNPs (TRAIL-R1 rs4242392 and IFN-γ rs2069707) were optimized at different temperatures by using gradient PCR (Table 1). Allele-specific polymerase chain reaction (AS-PCR) was performed using a 2720 thermal cycler (Applied Biosystems). Allele-specific forward primer1 (FP1) binding TRAIL-R1 rs4242392 (accession no: NG_032107.1) was specific for the “C” allele. Allele-specific forward primer2 (FP2) binding was specific for the “T” allele. Reverse primer (RP) was common in both FP1 and FP2. The size of both FP1 and FP2 was 129 bp (Fig. 1A).
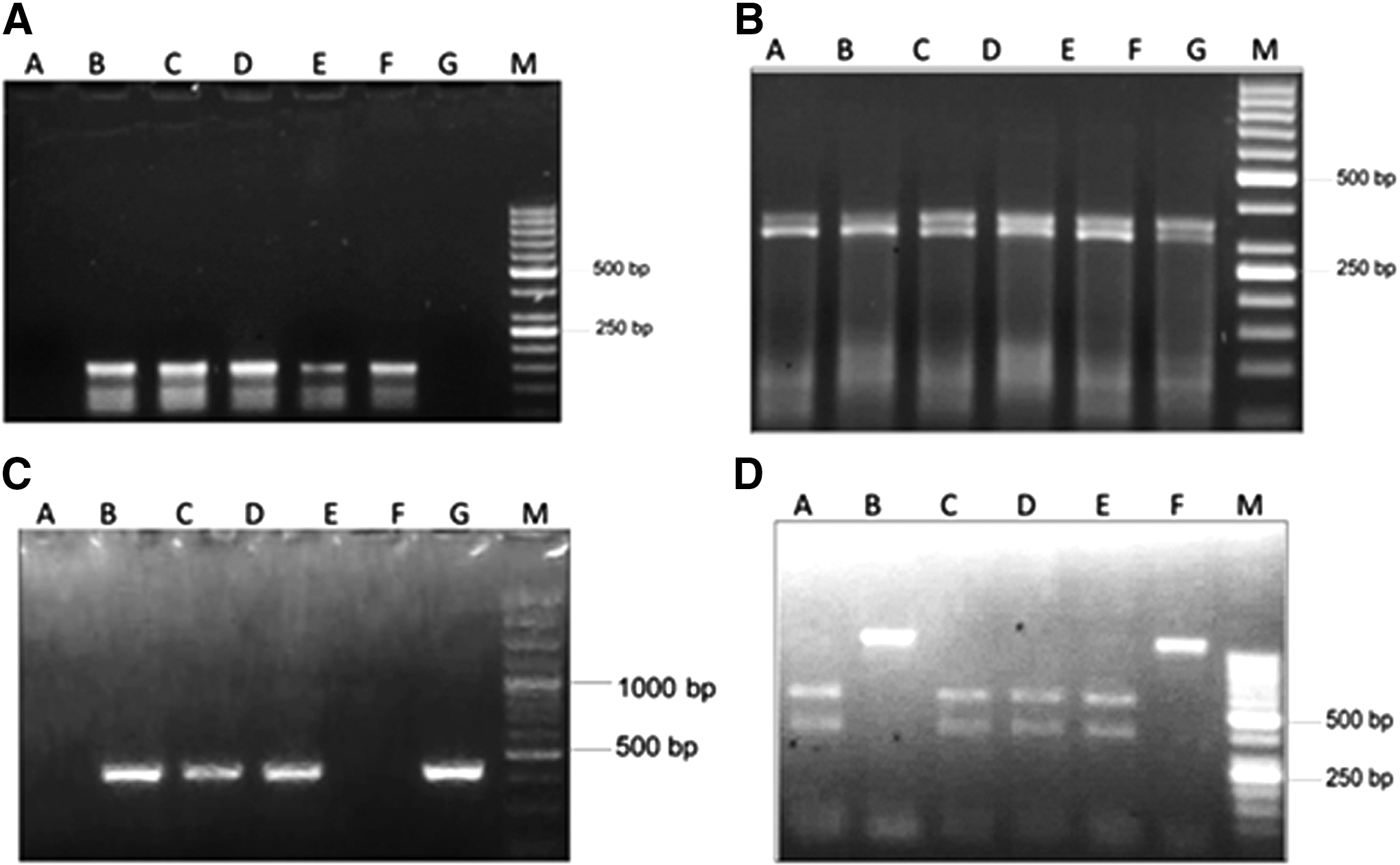
AS-PCR, allele-specific-PCR; PCR, polymerase chain reaction; RFLP-PCR, restriction fragment length polymorphisms-PCR.
Allele-specific FP1 binding IFN-γ rs2069707 (accession no: NG_015840.1) was specific for the “C” allele. Allele-specific FP2 binding was specific for the “G” allele. RP was common in both FP1 and FP2. The size of both sets of primers was 436 bp (Fig. 1C). The primers were designed using online Primer 3 software (Table 1). To validate the primer sequences scientifically, they were checked for their specificity by the National Centre for Biotechnology Information (NCBI) primer BLAST.
SNPs detection by restriction fragment length polymorphisms polymerase chain reaction
Restriction fragment length polymorphisms polymerase chain reaction (RFLP-PCR) analysis was also performed for the detection of the genetic variation of TRAIL-R1 rs4242392 and IFN-γ rs2069707. Using a theoretical restriction map program of online software Webcutter v2.0, restriction endonucleases ClaI (R012S, 4 IU/μL) and RsaI (R047S, 5 IU/μL) were used. The digestion of TRAIL-R1 rs4242392 650-bp PCR product by ClaI produced two bands of 308 bp and 342 bp (Fig. 1B). The digestion of IFN-γ rs2069707 1080 bp PCR products by RsaI produced two bands of 475 bp and 605 bp (Fig. 1D). The recipes of restriction digestion are shown in Table 2. AS-PCR and RFLP-PCR are routinely used for the detection of SNPs (21).
BSA, bovine serum albumin; RE, restriction endonuclease.
HCV genotyping
Hepatitis C viral RNA was extracted using a viral RNA isolation kit (catalog no: 09006100; RTA), according to the manufacturer's protocol. Viral RNA was amplified with a MyiQ2 two-color RT-PCR detection system (Bio-Rad). A gene proof HCV PCR kit designed for the quantification of HCV RNA was used according to the manufacturer's protocol. HCV genotyping was performed by Ohno's method (30).
Statistical analysis
Statistical analysis of the data was performed using Graph Pad Prism v5 and SPSS for Windows v13.0 (SPSS, Inc.). The Hardy–Weinberg equilibrium (HWE) was applied to analyze expected frequencies of alleles and genotypes in healthy controls, SVRs, and NRs. Pearson's chi-square analysis was performed to find any statistical association of genetic variations of studied genes with natural clearance and treatment outcomes of HCV infection. An independent t-test was applied to compare means±standard deviation of the studied groups. Two-tailed p-values of ≤0.05 were considered to be significant in chi-square tests.
Results
Association of viral load and HCV genotyping with therapy
Blood specimens of HCV patients 6 months after the end of therapy were analyzed for viral load to discriminate SVRs and NRs on the bases of viral load. In a total 118 chronic HCV patients, 80 (68%) patients attained SVR after completing the planned course of combinational therapy (IFN-α and ribavirin). The mean baseline viral load in SVR and NR patients was 4.3±4.3×106 IU/mL and 4.6±7.7±106 IU/mL respectively. Ninety-six chronic HCV patients (81.35%) were found to possess genotype 3, while the remaining 22 chronic HCV patients (18.64%) possessed other genotypes. HCV genotype 3 (p<0.05) and viral titer (p<0.05) were found to possess a significant correlation with treatment response. The clinical demographic details of patients are shown in Table 3.
HCV, hepatitis C virus; NRs, nonresponders; SVRs, sustained virological responders.
Association of TRAIL-R1 rs4242392 with HCV pathogenesis and treatment response
All the blood samples of HCV patients and healthy individuals were tested for the TRAIL-R1 rs4242392 genotypes by AS-PCR followed by RFLP-PCR analysis. Both techniques show similar results in all experiments. The distributions of different genotypes of TRAIL-R1 rs4242392 were according to the HWE. The distribution of the TRAIL-R1 rs4242392 genotypes TT, CT, and CC in chronic HCV patients was 65%, 33%, and 1%, while in healthy controls it was 56%, 39%, and 5% respectively. Pearson chi-square analysis of the TRAIL-R1 rs4242392 genotypes showed no significant association with natural clearance of HCV infection (p>0.05). The distribution of the TRAIL-R1 rs4242392 genotypes TT, CT, and CC in the SVR group was 78%, 21%, and 0%, while in the NR group it was 36%, 58%, and 5% respectively. Statistical analysis of genotype distribution showed a significant association of the TRAIL-R1 rs4242392TT genotype with the treatment response of chronic HCV infection (p<0.05; Table 4).
There was significant variation in the distribution of the TRAIL-R1 rs4242392 “T” allele between the SVR group (n=143, 89%) compared to the NR group (n=50, 65%; p<0.05). However, no significant association was found in the prevalence of the TRAIL-R1 rs4242392 “T” allele among healthy individuals (n=145, 75%) compared to chronic HCV patients (n=193, 81%; p>0.05; Fig. 2).
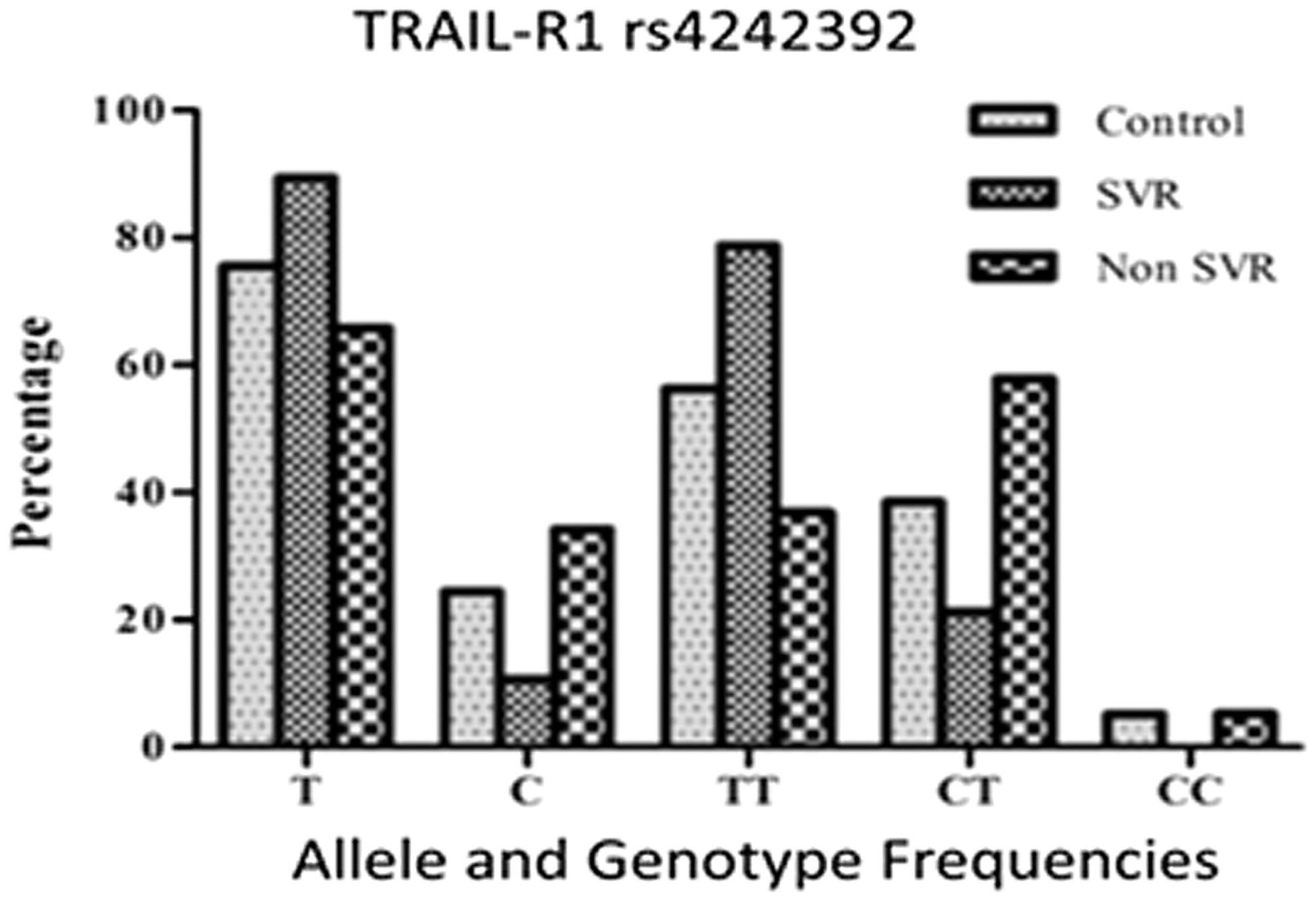
Prevalence of TRAIL-R1 rs4242392 polymorphism in healthy individuals, SVR and non-SVR. SVR, sustained virological responder.
Association of IFN-γ rs2069707 with HCV pathogenesis and treatment response
The expected frequencies of all IFN-γ rs2069707 genotypes in healthy controls, SVRs, and NRs were according to the HWE. The prevalence of the IFN-γ rs2069707 genotypes GG, GC, and CC in healthy controls and chronic HCV patients was 96%, 4%, and 0% and 1%, 20%, and 79% respectively. Pearson chi-square analysis of the IFN-γ rs2069707 genotypes for healthy controls and chronic HCV patients demonstrated significant associations of all genotypes (p<0.05). There was a significant difference in the prevalence of the IFN-γ rs2069707 “G” allele in healthy individuals (n=188, 98%) compared to chronic HCV patients (n=26, 11%). The p-value for the distribution of “G” allele frequencies in healthy controls and chronic HCV patients was significant (p<0.05; Table 4).
The prevalence of the IFN-γ rs2069707 genotypes GG, GC, and CC in SVR and NR groups was 1%, 21%, and 78% and 0%, 18%, and 82% respectively. The study showed no significant differences in the distribution of IFN-γ rs2069707 genotypes in SVR and NR groups (p>0.05). The occurrence of the “G” allele in the SVR group was n=19 (11%), and in the NR group it was n=7 (9%); both were comparable (p>0.05; Fig. 3).
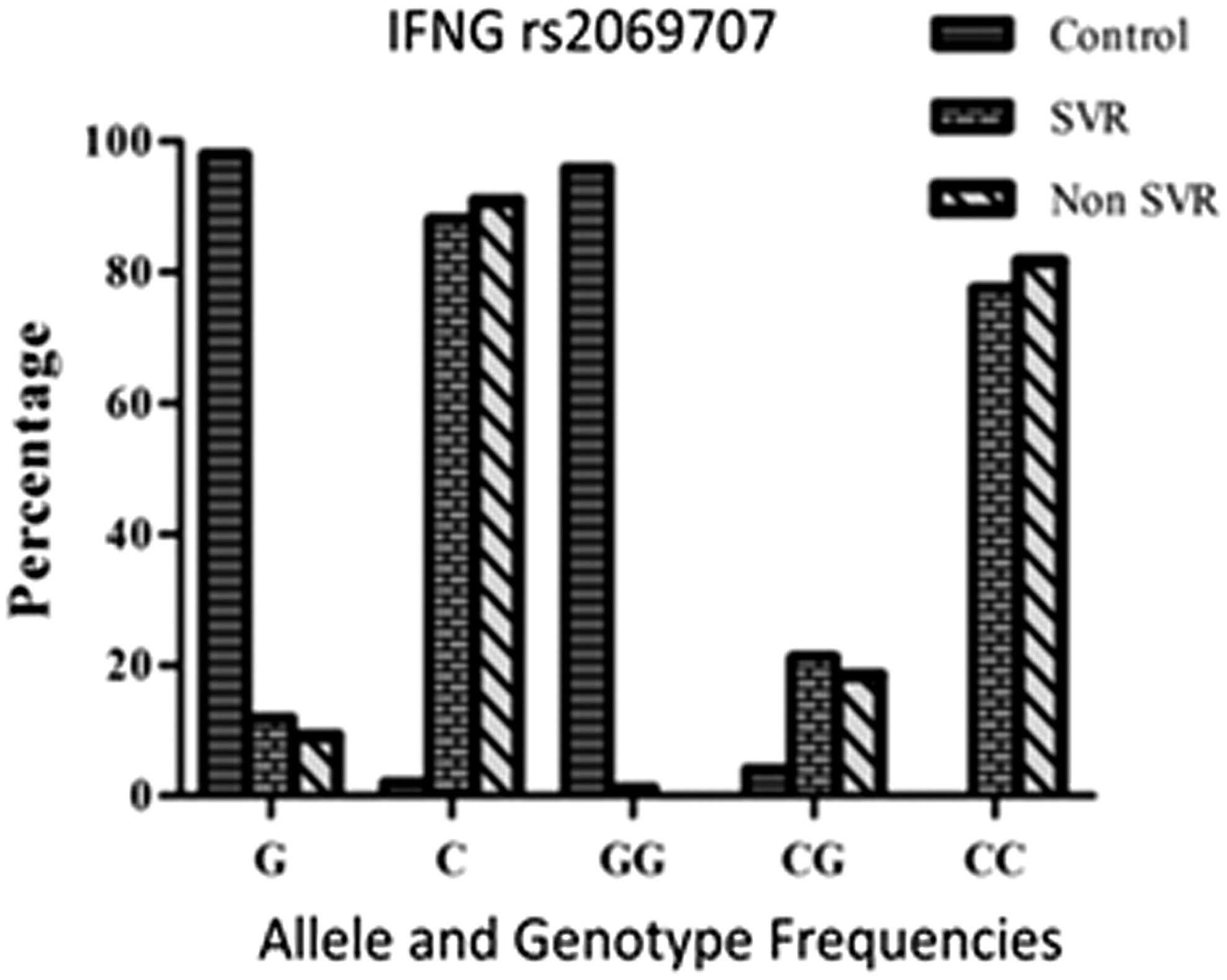
Alleles and genotype frequencies of IFNG rs2069707 SNP in healthy individuals, SVRs and nonresponders. SNP, single nucleotide polymorphism.
Discussion
Despite the discovery of new direct-acting antiviral agents, interferon and ribavirin are still important in the treatment of HCV infection (9). Therefore, the prediction of IFN/ribavirin-based therapies for HCV infection remains essential. The published data from worldwide studies have also established a significant correlation of viral titer and HCV genotypes with interferon-based therapies for HCV infection (13,17). HCV genotypes 1, 4, and 6 are difficult to treat and are often associated with a lower response, while HCV genotypes 2 and 3 have a better therapy response (1,42). The distribution of HCV sub-genotypes in the current study was 55% for 3a, 26.3% for 3b, 11.9% for 2a, 4.4% for 1a, and 3.4% for co-infected 3a+3b. Previous reports have also shown that the predominant HCV genotype in the Pakistani population is HCV genotype 3 (2). Similarly, early high viral load is linked with poor treatment response (20). The present study shows that viral genotype (p<0.05) and viral titer (p<0.05) are significantly associated with interferon plus ribavirin treatment of HCV infection. A number of host factors have been investigated and even used as biomarkers (28). Host genetic variations have gained attention in affecting HCV pathogenesis and response to IFN-based therapy in addition to stage of liver fibrosis, viral titer, and HCV genotypes (38). Other parameters such as age and gender were insignificantly associated with treatment outcomes of HCV infection (Table 3). Reports have shown that age and gender are controversially associated with interferon-based therapy for HCV infection (24).
Host genetic variations are important regarding HCV infection and response to interferon-based therapy (20). Recently, it has been shown that various SNPs in the vicinity of the IL28B gene were significantly linked with HCV pathogenesis and response to interferon-based therapy in worldwide populations (15). Genetic testing of IL28B has been added to the treatment guidelines for HCV genotype 1 in European countries (8). Similarly, IL23 promotes interferon-α responsiveness in HCV/HIV co-infected patients (29). The correlation of infectious diseases (mycobacterium tuberculosis, AIDS, hepatitis, etc.) and many other diseases with different SNPs can be made as relevant pharmacogenomic marks for drug therapy (11).
A key influence of IFN-α treatment for HCV infection is an enhanced expression of TRAIL that is also linked with SVR (35). IFN-induced T-cells and NK cells are likely to destroy virally infected cells with TRAIL receptors in SVR patients. Consistent with this model, HCV should be cured with IFN/ribavirin treatment, as it causes upregulation of TRAIL expression. TRAIL-R1 transduces apoptotic signal upon fusion with soluble or membrane-bound TRAIL ligands (5). The polymorphism in TRAIL-R1 may influence the therapy response of HCV infection. The distribution of the TRAIL-R1 rs4242392TT genotype in SVR and NR groups was n=63 (78%) and n=14 (36%) respectively (Table 4). A significant correlation between TRAIL-R1 rs4242392TT and combinational therapy HCV infection was shown in the study population (p<0.05). A recent report demonstrated that the presence of TRAIL-R1 rs4242392 exhibited a lack of responsive to HCV therapy in patients infected with both HIV and HCV (33). The difference in already reported results and the current results may be attributed to several factors such as initial viral load, HCV genotype, and most importantly the ethnic differences between the studied populations. Previously, it has been shown that the IL28B genetic mutation rs8099917 is dependent on ethnicity (26) while the IL28B genetic mutation rs12979860 is independent of ethnicity (10). Thus, the role of SNPs on disease progression and treatment response is highly dependent on ethnicity.
Several SNPs in IFN-inducible genes leading to altered gene expression or function were found to be associated with divergent responses to IFN treatment (7). The expression level of type 1 cytokine IFN-γ rs2069707 is correlated with liver pathogenicity as well as the possibility of no response to IFN-α therapy (12). In peripheral blood mononuclear cells of chronically infected HCV treatment NRs, the expression of IFN-γ was considerably lower compared to treatment responders (18). It was suggested that the high expression of IFN-γ was connected with liver fibrosis of HCV-infected patients (32). Studies have shown that the “G” variant at INF-γ −764 (rs2069707) is common in Caucasian Americans and may possibly be involved in frequent spontaneous recovery from HCV infection (40).
The present study showed no significant variation in prevalence of IFN-γ rs2069707 SNP in the SVR group (n=1, 1%) and the NR group (0%) of chronic HCV-infected patients (p>0.05). However, there were significant variations in the prevalence of IFN-γ rs2069707 SNP in healthy controls and chronically infected HCV patients (p<0.05; Table 4). Thus, there is a strong association between IFN-γ genotype rs2069707GG and the spontaneous clearance of HCV.
Conclusion
The current results suggested that TRAIL-R1 genetic variation rs4242392 might possess a role in interferon-based therapy for chronic HCV-infected patients, while the IFN-γ gene SNP rs2069707 is likely to contribute to the natural clearance of HCV infection.
Footnotes
Acknowledgments
We thank the National University of Sciences and Technology (NUST) H-12, Islamabad, Pakistan, for financial support for this study.
Author Disclosure Statement
No competing financial interests exist.