
Abstract
Several studies have shown a potential association between the Herpesviridae members, the Epstein–Barr virus (EBV) and Human herpes virus 6 (HHV-6), and an increased risk of autoimmune disease development. Because of the ability of these viruses to cause recurrent infections, various viral antigens, including viral DNA, are consistently shed. These antigens may then play a role in triggering autoimmune processes or contributing to autoimmune mechanisms. Therefore, this study examined whether the DNA of EBV or that of HHV-6A is capable of triggering IL-17, the autoimmune-associated cytokine, in mice. BALB/c mice were intraperitoneally injected with various copy numbers of either EBV or HHV-6A DNA. One group was injected with sterile water (the DNA solvent), and another was left uninjected. A mouse group that was administered DNA obtained from Staphylococcus epidermidis was included to ensure that any observed effects would pertain to the viral DNA tested. Mice were sacrificed and their sera were examined using an enzyme-linked immunosorbent assay for IL-17 and IL-23, as pro-autoimmune cytokines, IL-10, as an anti-inflammatory cytokine, and IFN-γ, as a pro-inflammatory cytokine, on days 3, 6, and 9 post-injection. All mouse groups injected with different copy numbers of EBV DNA or HHV-6A DNA displayed higher IL-17 levels than did the group injected with water on days 3, 6, and 9 post-injection. The highest IL-17 levels appeared to coincide with a marked increase in IL-23 and a decrease in IL-10 levels. Unlike the S. epidermidis DNA, which increased IFN-γ levels but not IL-17 or IL-23 levels, the viral DNA tested increased all three mediators, indicating that triggering Th17 responses is a specific property of EBV and HHV-6A DNA. In conclusion, EBV and HHV-6A viral DNA are capable of enhancing the production of the pro-inflammatory cytokine IL-17, which has been shown to play a role in autoimmune diseases.
Introduction
I
Contrary to its potentially beneficial effects in counteracting bacterial and fungal infections, IL-17 is believed to be involved in the pathogenesis of various autoimmune diseases. These include multiple sclerosis (MS), rheumatoid arthritis (RA), systemic lupus erythematosus (SLE), and inflammatory bowel disease (IBD) among other autoimmune diseases. Elevated levels of IL-17 have been detected in sera and target tissues of patients with these diseases among others (29). Mice with IL-17 or IL-17 receptor deficiency are less predisposed to experimental autoimmune encephalomyelitis (EAE) induction; these mice develop EAE with milder symptoms (21,29), including delayed onset, diminished severity, and early recovery (17). Moreover, targeting IL-17 has shown promising therapeutic potential in clinical trials for RA (11) among other autoimmune or immune-mediated diseases (32).
The authors have recently reported that previous Human herpes virus 6 (HHV-6) infections enhance the risk of MS (9,26). HHV-6 belongs to the Herpesviridae, a large family of enveloped double-stranded DNA viruses capable of establishing latency and causing recurrent infections. Evidence indicates that another member of the Herpesviridae family, the Epstein–Barr virus (EBV), also has strong correlations with MS, SLE, and RA among other autoimmune diseases (1,2,6,14,17,19,22,24,25). Due to the ability of these viruses to cause recurrent infections, various viral antigens, including viral DNA, are consistently shed. These antigens may then play a role in triggering autoimmune processes or contributing to autoimmune mechanisms such as enhancing IL-17 production. Studies have previously shown that DNA from other members of the Herpesviridae have immunostimulatory capabilities (23). These observations prompted the assessment of whether DNA from HHV-6 and from EBV was capable of inducing the pro-autoimmune cytokines, IL-17 and IL-23, in mice. The effect of such a treatment on the production of IFN-γ, as a pro-inflammatory cytokine, and on IL-10, as a potential immunoregulatory cytokine that may play a role in circumventing autoimmune responses (4), was also assessed.
Materials and Methods
Treatment of BALB/c mice with viral DNA
To study the effect of EBV and HHV-6 type A (HHV-6A) viral DNA on IL-17 and IL-10 production, nine groups of 4–6-week-old female BALB/c mice were used. Each group included nine mice. Mice were obtained from the Animal Care Facility at the American University of Beirut (AUB) and treated according to the guidelines of the Institutional Animal Care and Use Committee (IACUC) at AUB.
EBV and HHV-6A DNA were obtained from Advanced Biotechnologies (Columbia, MD). Viral genome copy numbers were specified by the manufacturer. Mouse group 1 received no injections, while group 2 received 100 μL sterile water, the DNA solvent. Thus, both groups served as negative controls. Groups 3, 4, and 5 received 36×103, 72×103, and 144×103 copies of EBV DNA respectively. Groups 6, 7, and 8 similarly received 36×103, 72×103, and 144×103 copies of HHV-6A DNA respectively. Mice in group 9 each received 28.3 pg of DNA obtained from an isolate of Staphylococcus epidermidis (amount equivalent to 144×103 copies of EBV DNA based on an EBV genomic molecular weight of 1×108). Purity of all DNA preparations utilized was verified by spectrophotometery. All injections were administered intraperitoneally, and each consisted of 100 μL.
Three mice per day from each group were sacrificed on days 3, 6, and 9 post-injection. Blood samples were drawn after cardiac puncture. Sera were then collected and assessed by enzyme-linked immunosorbent assay (ELISA) in duplicate as described below.
ELISA for IL-17, IL-10, IL-23, and IFN-γ
Mouse IL-17, IL-10, and IL-23 levels were assessed by ELISA using specific kits from Abcam (Cambridge, England), whereas IFN-γ was analyzed using a Single Analyte ELISArray Kit (SABiosciences, Frederick, MD).
Statistical analysis
Unpaired t-tests were calculated with the GraphPad t-test calculator. Viral DNA-treated mouse groups were compared to the sterile water-injected group per day post-injection. p-Values of <0.05 were considered statistically significant.
Results
Mouse serum IL-17 levels
BALB/c mouse groups injected with 36×103, 72×103, or 144×103 EBV genome copies all showed an increase in IL-17 levels compared to the group injected with water on days 3, 6, and 9 post-injection (Fig. 1A). A range of 2- to 11-fold increase was noted, with the highest level being observed in the group of mice administered 144×103 EBV DNA copies on day 6 post-injection (p=0.002). Levels in the group receiving 36×103 EBV DNA copies and in the group administered 144×103 copies peaked on day 6 post-injection, whereas levels in the group that was injected with 72×103 copies peaked on day 3 post-injection.
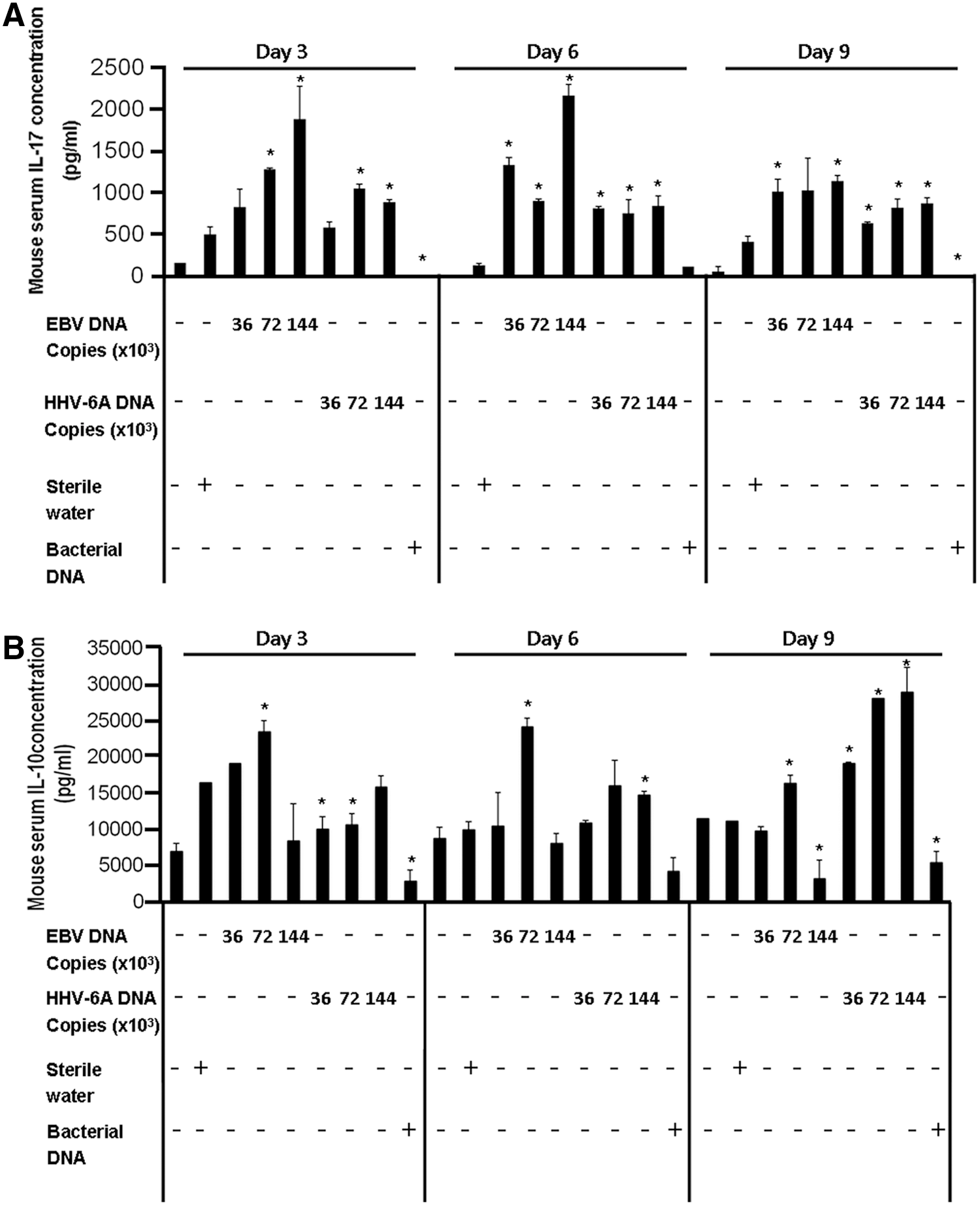
Serum IL-17 and IL-10 levels in mice injected with Epstein–Barr virus (EBV) or Human herpes virus 6 (HHV-6A) DNA. BALB/c mice were injected with either EBV or HHV-6A DNA. A group injected with DNA from Staphylococcus epidermidis was included as control. Mice were sacrificed in triplicate from each group on days 3, 6, and 9 post-injection, and their sera were analyzed for interleukin levels by enzyme-linked immunosorbent assay (ELISA). *p<0.05 compared to a sterile water-injected group. (
Similar to mice given EBV DNA, groups administered 36×103, 72×103, or 144×103 HHV-6A genome copies all showed an increase in serum IL-17 levels compared to the group injected with water on the days tested during the monitoring period. A range of 1.2- to 7-fold increase was noted, with the highest level being observed in the group that received 72×103 copies on day 3 post-injection (p=0.017). Levels in the group that received 36×103 copies were at their highest on day 6, whereas groups administered 72×103 or 144×103 copies of HHV-6A DNA were at their highest on day 3. HHV-6A DNA was selected for the studies presented herein, since it is the subtype of HHV-6 that has been most associated with autoimmune diseases.
The mouse group injected with DNA obtained from S. epidermidis did not display any increase in IL-17 levels on days 3, 6, and 9 post-injection. This treatment was utilized as a non-viral DNA control group.
Mouse serum IL-10 levels
Levels of IL-10 obtained upon injection of BALB/c mice with 36×103 or 72×103 copies of EBV DNA were not consistent with a relevant pattern (Fig. 1B). However, injecting 144×103 copies of EBV DNA appeared to lower IL-10 levels on days 3, 6, and 9 post-injection; this decrease in IL-10 coincided with the highest levels of IL-17 detected amongst the various treatments tested. No such effect was seen with HHV-6A DNA. Therefore, EBV DNA appears to be a better suppressor of IL-10 production.
A group of mice injected with DNA obtained from S. epidermidis had levels of IL-10 that were lower than those of the water-injected group on all days of testing.
Mouse serum IL-23 levels
All BALB/c mouse groups injected with EBV DNA displayed an increase in IL-23 levels compared to the group injected with water on days 3, 6, and 9 post-injection (Fig. 2A). A range of 1.8- to 8.3-fold increase was noted, with the highest levels being observed in the group of mice administered 144×103 EBV DNA copies on all days post-injection.
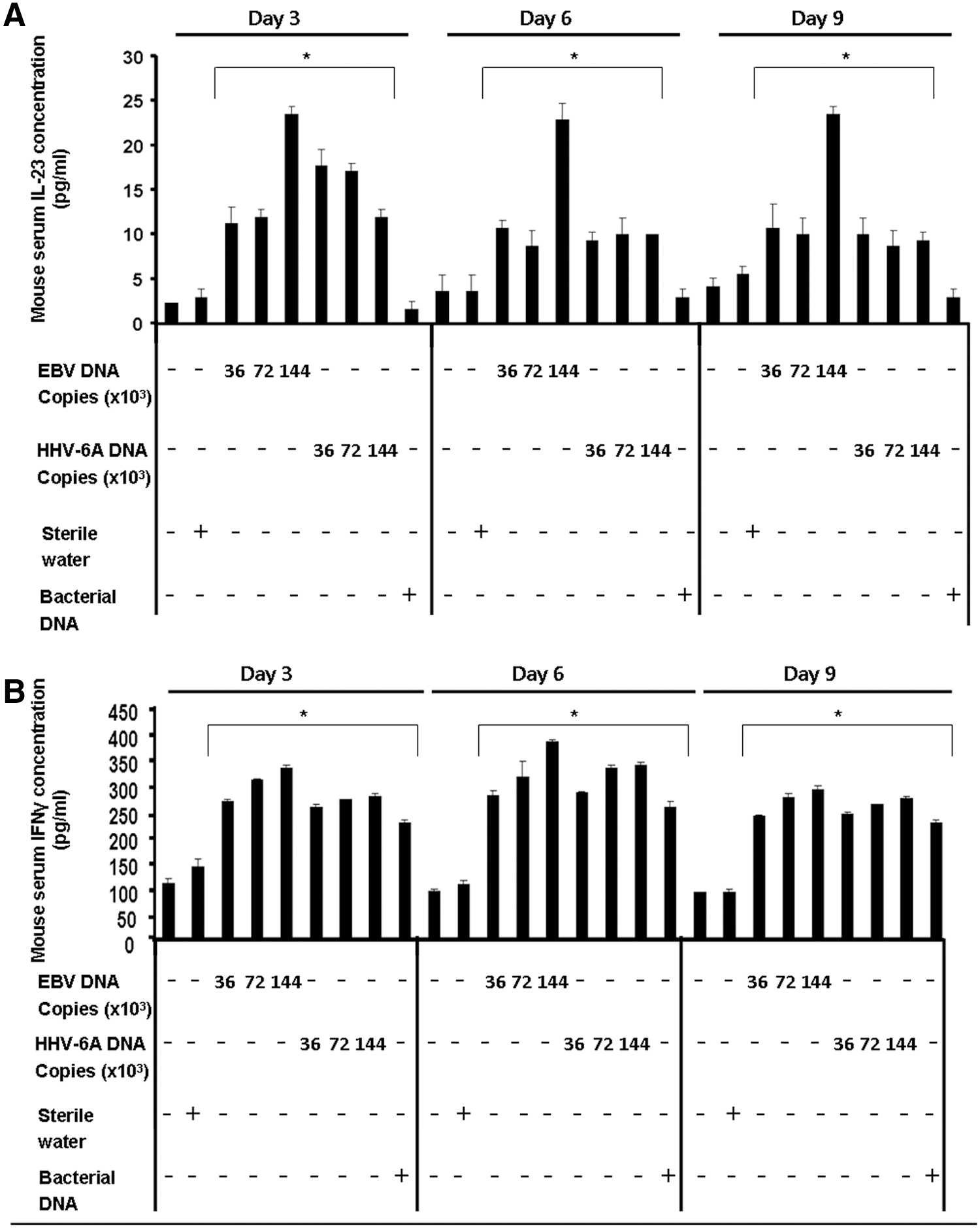
Serum IL-23 and IFN-γ levels in mice injected with EBV or HHV-6A DNA. BALB/c mice were injected with either EBV or HHV-6A DNA. A group injected with DNA from Staphylococcus epidermidis was included as control. Mice were sacrificed in triplicate from each group on days 3, 6, and 9 post-injection, and their sera were analyzed for interleukin levels by ELISA. *p<0.05 compared to a sterile water-injected group. (
Similar to mice given EBV DNA, groups administered 36×103, 72×103, or 144×103 HHV-6A genome copies all showed an increase in serum IL-23 levels. A range of 1.6- to 6.2-fold increase was noted with the highest level detected on day 3 post-injection.
The mouse group injected with DNA obtained from S. epidermidis did not display any increase in IL-23 levels compared to the water-injected group.
Mouse serum IFN-γ levels
All mouse groups that received microbial DNA displayed increased serum levels of IFN-γ compared to the water-injected group. Hence, unlike the observations with IL-17 and IL-23, both Herpesviridae DNA and S. epidermidis DNA were capable of inducing IFN-γ; S. epidermidis DNA did not induce an increase in IL-17 or IL-23 levels as denoted above. IFN-γ levels in all groups injected with microbial DNA peaked on day 6. Herpesviridae DNA resulted in a 1.8- to 3.5-fold increase in IFN-γ levels, while S. epidermidis DNA led to a 1.6- to 2.4-fold increase in this mediator.
Discussion
All mouse groups injected with different copy numbers of EBV DNA or HHV-6A DNA displayed higher IL-17 levels than the group injected with water on days 3, 6, and 9 post-injection. This indicates that Herpesviridae DNA, particularly that of HHV-6A and EBV, may trigger pro-autoimmune pathways, since it leads to increased levels of the autoimmunity-related cytokine IL-17. This type of DNA may also suppress the anti-inflammatory cytokine IL-10. However, this appears to be DNA copy number dependent.
The highest level of IL-17 observed with EBV DNA was on day 6 post-injection, whereas the highest level observed with HHV-6A DNA was on day 3 post-injection. Consequently, EBV DNA seems to require more time than HHV-6A DNA to trigger IL-17 production. However, overall, EBV DNA appears to be a better trigger of IL-17 than HHV-6A DNA, since IL-17 levels are higher in the EBV DNA injected groups compared to the HHV-6A DNA injected groups.
Injecting 144×103 copies of EBV DNA resulted in a decrease in IL-10 levels on days 3, 6, and 9 post-injection. This decrease coincided with the highest levels of IL-17 detected. This may indicate that a particular mechanism is triggered at this concentration of EBV DNA that results in depressing IL-10 expression. This mechanism may also be behind the increase in IL-17 levels seen with 144×103 copies of EBV DNA on days 3, 6, and 9. IL-23, a cytokine produced by dendritic cells and macrophages and associated with the Th17 cell expansion, is also capable of suppressing IL-10 production (1,6,18). To investigate whether IL-23 triggered IL-17 expression and suppressed that of IL-10 with this concentration of EBV DNA, its levels in mouse sera were assessed. The levels of IL-23 largely mirrored those of IL-17, also being at their highest with 144×103 copies of EBV DNA on days 3, 6, and 9. The level of IL-23 in this group at these time points was about 23 pg/mL. On the other hand, the rise of IL-17 on days 3, 6, and 9 upon injection of 36×103 or 72×103 copies of EBV DNA was not accompanied by a decrease in IL-10; levels of IL-23 with these treatments at these time points was 8.6–12 pg/mL. This indicates that 36×103 or 72×103 copies of EBV DNA trigger IL-23 levels that are sufficient to increase IL-17 production but not to decrease that of IL-10.
Worth noting is that the mouse group injected with bacterial DNA had very low IL-10 and IL-17 levels. Therefore, the effects observed are rather specific to the viral DNA at the tested concentrations. Moreover, assessment of Th1 responses via examining IFN-γ production revealed that both EBV and HHV-6A DNA can trigger this mediator. While inter-regulatory inhibitory dynamics have been described for IFN-γ and IL-17 (28,30), both mediators have been reported to rise concomitantly after particular challenges, vaccinations, and in some disease states (7 –9,12,20). The inhibitory effects of IFN-γ and IL-17 on each other may thus be challenge dependent or reliant on a threshold of activation that was not attained by the cytokine levels detected. Several previous studies have demonstrated that the Herpesviridae are capable of modulating the immune response potentially via affecting Toll-like receptor (TLR) signaling (10,13,26,27). Immunostimulatory effects of DNA from HSV-1, another member of the Herpesviridae family, via triggering Th1 responses has also been reported (15,16,23). None of these have, however, examined the effect of DNA, be it viral or otherwise, on Th17 responses. S. epidermidis DNA was capable of increasing IFN-γ levels but not those of IL-17 or IL-23; the Herpesviridae DNA tested, on the other hand, increased the levels of all three mediators, indicating that triggering Th17 responses is a specific property of EBV and HHV-6A DNA.
Various cellular mediators may respond to the presence of viral DNA and result in the production of IL-17. Multiple pattern recognition receptors have been documented to respond to viral DNA, including the TLRs. TLR3 and TLR9 have been previously implicated in the production of Th1 pro-inflammatory cytokines in response to DNA from HSV-1(15). Whether they play a role in triggering Th17 responses remains to be seen. Other potential candidates include the novel DNA binding factors and the nucleotide-binding domain-leucine-rich repeat-containing molecules, which recognize viral nucleic acids. The DNA-dependent activator of IFN-regulatory factors (DAI), also known as ZBP-1, may also mediate similar responses. The absence in melanoma 2 (AIM2) protein has also been shown to bind cytosolic viral double-stranded DNA (31) directly and may serve as a potential mediator as well.
In conclusion, EBV and HHV-6A viral DNA are capable of triggering IL-17, as indicated by the present findings. Further research is required to elucidate the precise roles played by viral DNA in triggering this cytokine. Understanding these roles may help in assessing disease risk groups, formulating disease and relapse prevention measures, and contributing to drug design and development.
Footnotes
Acknowledgments
The authors wish to thank the Lebanese National Council for Scientific Research (CNRS) for funding this work.
Author Disclosure Statement
No competing financial interests exist.