
Abstract
Early responses against viruses, such as avian influenza virus (AIV), may be induced by Toll-like receptor (TLR) pathways. In the present study, an in ovo model was employed to study the antiviral activities of TLR ligands. It was hypothesized that administration of TLR ligands in ovo at the appropriate dose and time can reduce AIV titer in embryonated chicken eggs. Moreover, the study aimed to determine the mechanisms involved in the TLR-mediated antiviral responses in the chorioallantoic membrane (CAM). Embryonated eggs (10–14 day old) were treated with TLR2, 4, 7, and 21 ligands using different doses and times pre- and post-AIV infection. The results revealed that treatment of embryonated chicken eggs with TLR ligands reduced AIV replication. Further analysis showed that TLR ligands induced interferon (IFN)-γ and IFN stimulatory genes in the CAM, which may have played a role in the reduction of the AIV titer. The timing and dose of TLR ligands administration had significant impacts on the outcome of the treated eggs. In conclusion, the present study demonstrated that the in ovo route may be employed to determine the antiviral characteristics of TLR ligands against AIV.
Introduction
H
In general, each TLR binds to pathogen-associated molecular patterns (PAMPs) or their synthetic analogs. Activation of TLR signaling pathways leads to a variety of cellular responses, including the production of interferons (IFNs). It has been shown in mice that stimulation of TLRs with their ligands prior to influenza virus infection increases resistance to highly pathogenic influenza viruses (36,49,52). In chickens, administration of TLR ligands induces the expression of antiviral and pro-inflammatory cytokines and IFN-stimulated genes (ISGs), such as oligoadenylate synthase (OAS) (41). In addition, it has previously been demonstrated that administration of TLR ligands, including polyinosinic:polycytidylic acid (polyI:C), CpG oligodeoxynucleotides (CpG ODN), and lipopolysaccharide (LPS), to chickens 24 h prior to infection with AIV significantly reduced viral shedding (42). Further results demonstrated that poly I:C, CpG ODN, and LPS treatment enhanced the protective effect of vaccination against influenza virus in vivo (38). The molecular mechanisms responsible for the observed reduction in viral shedding are not completely understood.
There is a wide range of available synthetic ligands that could be assessed as antiviral agents in chickens, and the efficacy of these TLR ligands may be different from each other. Moreover, the potency of different ligands in chickens is likely to differ from what is seen in mammalian species (3,6,40). In order to determine the antiviral efficacy of TLR ligands in chickens, these ligands have been screened in vivo and in vitro (3,42). However, the in ovo route offers an alternative method to screen and assess the efficacy of TLR ligands as antiviral agents in chickens. Previous studies have demonstrated host responses in embryonated chicken eggs following infection with various pathogens, which confirms the ability of the chick embryo to mount a response (2,15,21). The best example of the ability of the chicken embryo to mount an immune response is in ovo vaccination to control infectious diseases in chickens, such as infectious bursal disease (IBD), coccidiosis, and Marek's disease (9,22).
To date, there is little information available about innate responses following in ovo administration of TLR ligands (7,47). Previously, it was shown that the TLR7 ligand, Loxoribine, inhibits the replication of AIV in ovo (45). AIV replicates in cells of the chorioallantoic membrane (CAM), and virus particles are released into the allantoic fluid (37). It has been shown that chicken embryos, more specifically the CAM, can respond to stimuli or pathogens (11,12). Previous studies demonstrated that the CAM expresses various cytokines such as IFN-γ, IL-1β, IL-8, and IL-12. Moreover, some cells of the immune systems, such as macrophages, are present in the CAM (11,12,19,20). As a result, there is a possibility that TLR ligands induce antiviral responses in the CAM where AIV replicates, and hence may interfere with replication of the virus.
The present study examined whether activation of innate immune responses of embryonated chicken eggs by TLR ligands can influence AIV replication. To this end, it was demonstrated that pretreatment of embryonated eggs with TLR ligands can restrict virus replication in the eggs. Moreover, to expand the understanding of TLR ligand activities, possible mechanisms associated with TLR-mediated antiviral effects in the CAM were assessed.
Materials and Methods
AIV
The A/Duck/Czech/56 (H4N6) low pathogenic AIV was used in the present study. The virus was propagated in 11-day-old embryonated chicken eggs by inoculation through the allantoic cavity (47). Briefly, embryonated chicken eggs were candled, and 100 μL of allantoic fluid containing 0.2 hemagglutination (HA) units of H4N6 AIV was injected. The allantoic fluid was harvested 72 h later.
TLR ligands
Pam3CSK4 (synthetic triacylated lipoprotein) and R848 were purchased from InvivoGen (San Diego, CA). Synthetic class B CpG ODN 1826 [5′- TCC ATG ACG TTC CTG ACG TT-3′], non-CpG ODN [5′-TGC TGC TTG TGC TTT TGT GCT T-3′], LPS from Escherichia coli 0111:B4, and E. coli 026:B6 were purchased from Sigma-Aldrich (Oakville, Canada). These ligands were selected, as they have previously been shown to stimulate chicken TLRs (40,44).
Embryonated egg treatment with TLR ligands and infection with AIV
Embryonated chicken eggs were treated with different doses of TLR ligands as described in Table 1. Ten-day-old embryonated chicken eggs were candled, their surface was disinfected with 70% ethanol, and TLR ligands were administered directly into the allantoic fluid using a 25 gauge, 1 inch needle. The eggs were then sealed with Permount mounting media. After 24 h of incubation, embryonated chicken eggs were infected with 0.2 HA units of H4N6 AIV. The allantoic fluid was harvested 72 h later, and the virus titer was quantified via the hemagglutination test (51).
Optimum dose of each TLR ligand.
CpG ODN, CpG oligodeoxynucleotides; LPS, lipopolysaccharide; TLR, Toll-like receptor.
In order to determine the effect of in ovo administration of TLR ligands on the replication of H4N6 AIV in embryonated chicken eggs, eggs were treated with the optimum dose of the various TLR ligands (as determined above) at different time points. Specifically the time points included co-administration of TLR ligands and H4N6 AIV as well as treatment 72, 48, 36, 24, and 12 h prior to infection, and 36, 24, and 12 h post-infection. The virus titer in the allantoic fluid was quantified using an end-point dilution in Madin–Darby canine kidney (MDCK) cells and was expressed as 50% tissue culture infective dose (TCID50)/mL according to the Reed–Muench formula (51). There were 10 biological replicates in each group.
Gene expression of the CAM stimulated with TLR ligands
Eleven-day-old embryonated chicken eggs were treated with either LPS from E. coli 026:B6 (2 μg/egg), Pam3CSK4 (10 μg/egg), R848 (10 μg/egg), or CpG ODN 1826 (2 μg/egg). The control groups were treated with non-CpG ODN (20 μg/mL) or phosphate-buffered saline (PBS) only. At 3, 8, and 18 h post-treatment, the surface of eggs containing viable embryos (determined by candling) were disinfected with 70% ethanol. Then, the shell was cut in half with scissors under sterile conditions. The adhering CAM was collected for RNA extraction. There were six replicates in each group.
RNA extraction and cDNA synthesis
Total RNA was extracted from the CAM using Trizol reagent (Life Technologies, Burlington, Canada), according to the manufacturer's recommendations. Total RNA was treated with the DNA-Free™ kit (Ambion, Austin, TX), and 1 μg of RNA was used for cDNA synthesis using Superscript II First Strand Synthesis kit (Life Technologies) and oligo-dT primers, according to the manufacturer's protocol.
Real-time polymerase chain reaction
Quantitative real-time polymerase chain reaction (PCR) was performed on diluted cDNA (1:10 in diethylpyrocarbonate-treated water) using a SYBR green dye in a LightCycler 480 II (Roche Diagnostics, Laval, Canada) as previously described (4,41). Briefly, the amplification conditions consisted of preincubation for 10 min at 94°C, followed by 45 cycles for 10 sec at 95°C, 55–64°C annealing as described in Table 2 for each of the primers for 5 sec, and elongation and signal acquisition (single mode) at 72°C for 10 sec. Melt curve analysis was done in three steps: 95°C for 10 sec, cooling to 65°C for 1 min, and heating to 97°C. Specific sequences of primers were described previously, and are listed in Table 2 (1,3,4,5,39,43,50).
IFN, interferon; IRF, IFN regulatory factor; OAS, oligoadenylate synthase.
Statistical analysis
Statistical analysis of the virus titer was performed by one-way analysis of variance followed by Tukey's post hoc test for multiple comparisons to examine the effects of TLR ligands. p-Values of <0.05 were considered to be statistically significant.
For gene expression, fold changes and standard errors were calculated using REST (Relative Expression Software Tool) software v2009 (Qiagen, Valencia, CA). The REST software compared control (PBS) and treatment groups using pairwise fixed reallocation randomization based on the PCR efficiencies and the mean crossing point deviations between the PBS control and treatment groups.
Results
The CAM expresses TLRs
The CAM from the control group (PBS treated) was examined first, and it was discovered that the cells within the CAM expressed TLR2, 4, and 21 at the transcript level at 3, 8, and 18 h after PBS injection. TLR7 was also expressed at all three time points, although to a lesser extent (Fig. 1).
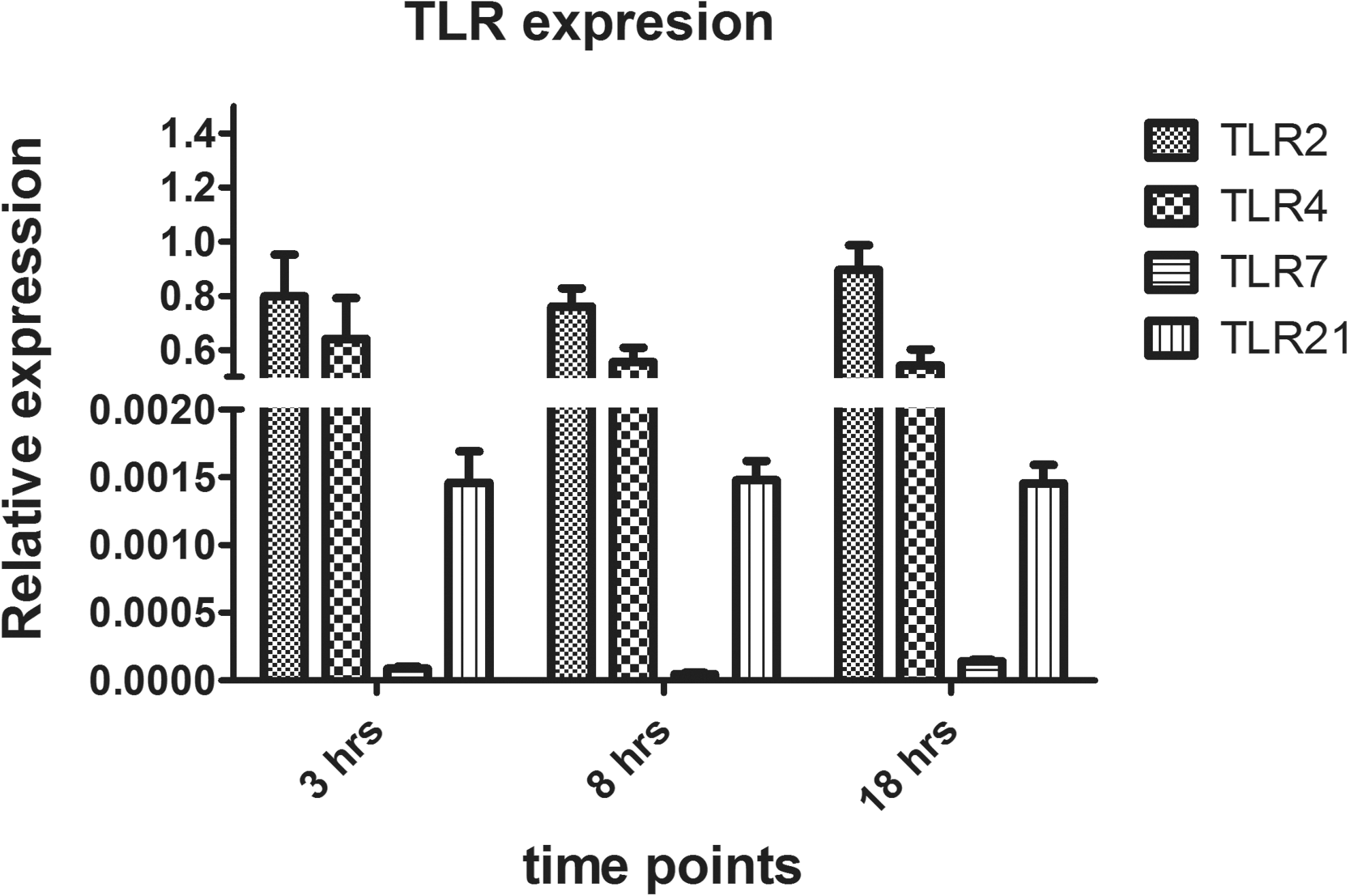
Relative expression of Toll-like receptor (TLR) genes in the chorioallantoic membrane (CAM). Eleven-day-old embryonated eggs were treated with phosphate-buffered saline (PBS; 100 μl/egg). Gene expression of TLR2, 4, 7, and 21 was evaluated relative to the housekeeping gene β-actin at 3, 8, and 18 h after PBS injection into the embryonated eggs. There were six replicates at each time point.
TLR2, 4, 7, and 21 ligands decrease viral replication in embryonated eggs
To examine the effects of TLR ligands on virus titer, embryonated eggs were treated with three different doses of TLR2, 4, 7, and 21 ligands, as listed in Table 1, for 24 h prior to infection. The treatment of embryonated eggs with the high dose of LPS, CpG ODN 1826, and Pam3CSK4 significantly reduced the virus titer in the allantoic fluid (Fig. 2). However, the mortality of the embryos was higher in these treatment groups compared to those that received lower doses of TLR ligands (Table 3). As a result, the high dose was excluded from subsequent experiments. There was also a significant decrease in viral titer in the allantoic fluid of embryonated eggs treated with the intermediate dose of LPS, CpG ODN 1826, R848, and Pam3C SK4 compared to eggs administrated PBS (Fig. 2). The low dose of LPS and CPG ODN 1826 also significantly reduced the virus titer in the embryonated eggs. However, the low dose did not reduce viral replication as efficiently as the intermediate dose (p<0.05). As a result, the intermediate dose of all the TLR ligands was selected for further experiments.
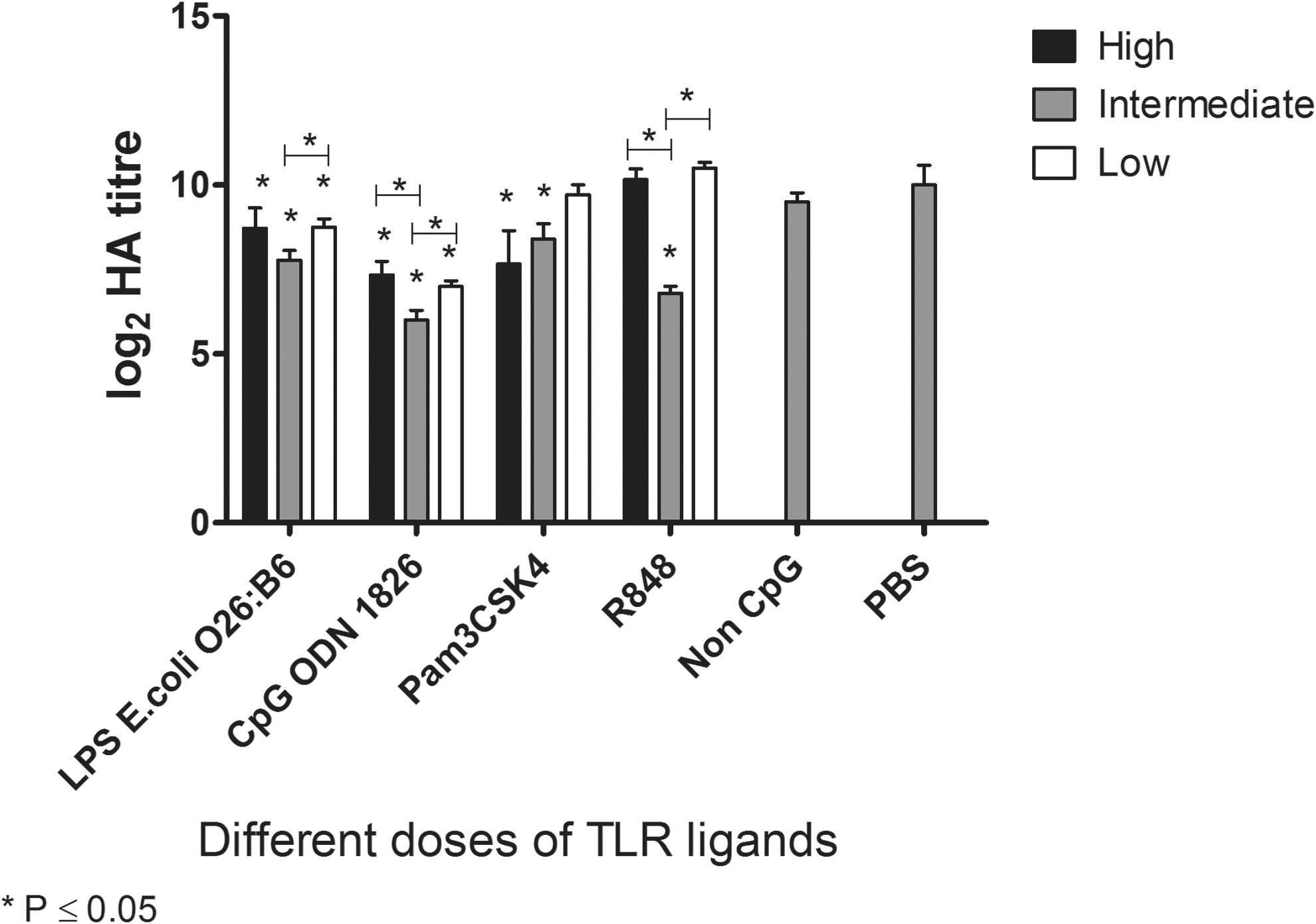
Treatment of 10-day-old embryonated chicken eggs with different doses of TLR ligands. Embryonated eggs were treated with three doses of Pam3CSK4 (100, 10, and 1 μg/egg), lipopolysaccharide (LPS) from Escherichia coli 026:B6 (20, 2, and 0.2 μg/egg), R848 (50, 10, and 1 μg/egg), class B CpG ODN 1826 (20, 2, and 0.2 μg/egg), and non-CpG ODN (20 μg/egg) for 24 h. The control group received PBS (100 μl/egg). The embryonated eggs were then infected with the A/Duck/Czech/56 (H4N6) low pathogenic avian influenza virus (AIV) for 72 h. Virus titer is represented by the log2 hemagglutination (HA) unit. Each TLR ligand dose was compared to the PBS group using one-way analysis of variance (ANOVA), with significant differences (p≤0.05) being indicated by an asterisk. PBS, phosphate-buffered saline.
To determine the optimal time for TLR ligand treatment, nine different time points (72, 48, 36, 24, and 12 h prior to infection, time of infection, and 12, 24, and 36 h post-infection) were selected, and TLR ligands were administered before the virus titer in allantoic fluid was assessed. The treatment of embryonated eggs with LPS from E. coli O26:B6 at 72, 48, and 24 h prior to infection, at the time of infection, and at 24 h post-infection significantly reduced the virus titer in the allantoic fluid (Fig. 3A). Treatment of embryonated eggs with LPS from E. coli O111:B4 at the time of infection or at 12, 24, and 36 h post-infection significantly reduced the virus titer in the allantoic fluid (Fig. 3A). Moreover, eggs treated with Pam3CSK4 at 36 and 24 h prior to infection had lower virus titers compared to eggs that received PBS (p<0.05; Fig. 3B). Treatment of embryonated eggs with CpG ODN 1826 at 36, 24, and 12 h prior to infection significantly reduced the virus titer in the allantoic fluid (Fig. 3C). Eggs treated with R848 at 36 and 24 h prior to infection had significantly lower virus titers compared to the eggs that received PBS at the corresponding time points (Fig. 3D).
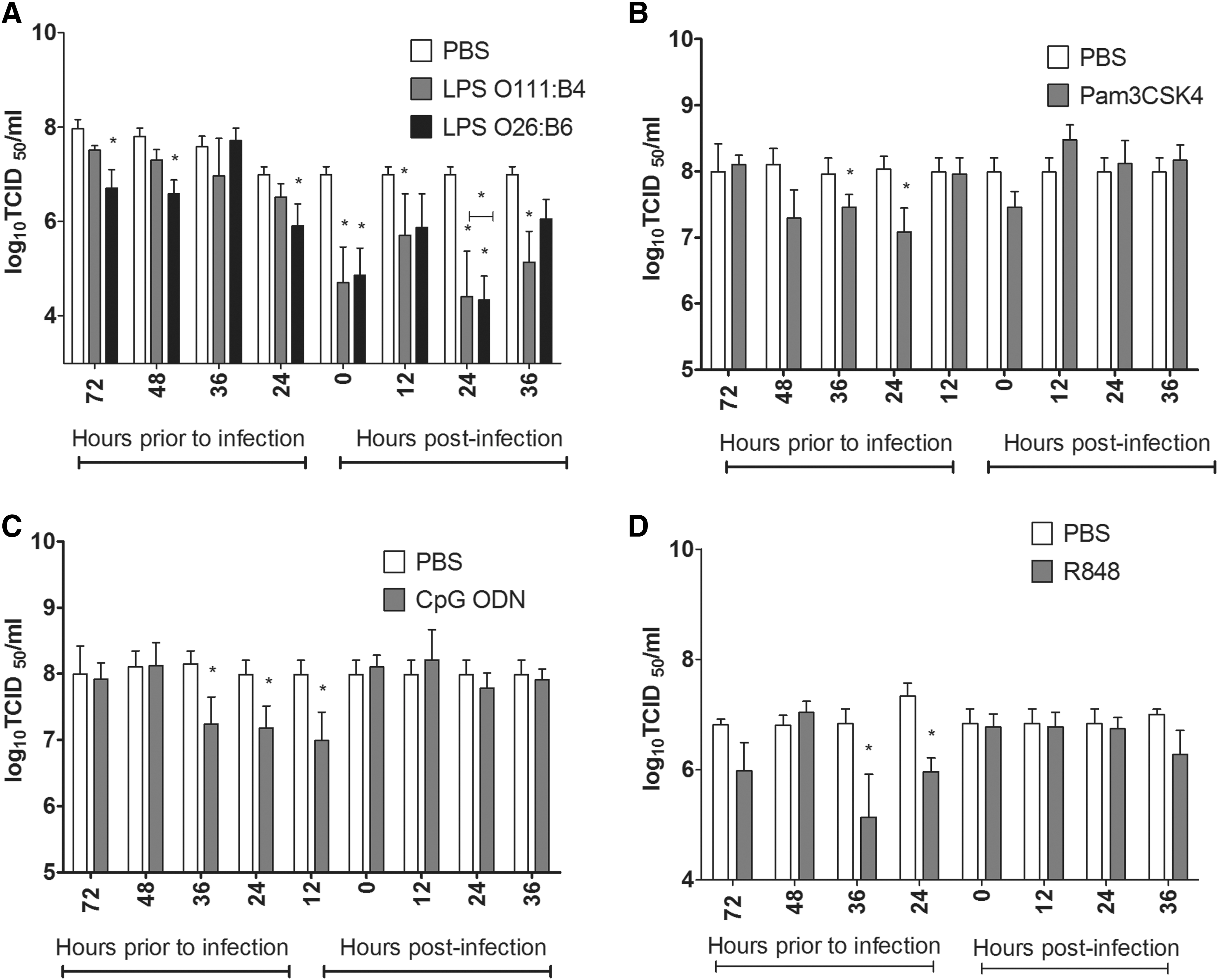
Treatment of embryonated eggs with TLR ligands at different time points. The embryonated eggs were treated with 2 μg/egg of LPS from E. coli 026:B6 (
Induction of gene expression in the CAM by TLR ligands
The expression of IL-1β was significantly increased in the CAM treated with LPS at 3, 8, and 18 h post-treatment by 630-, 137-, and 58-fold, respectively (Fig. 4A). The expression of IL-1β by the CAM from eggs that received Pam3CSK4 was significantly increased at 3, 8, and 18 h post-treatment with 169-, 48-, and 33-fold increases, respectively. Moreover, R848 induced the expression of IL-1β by the CAM at 3, 8, and 18 h post-treatment with 30-, 22-, and 9-fold increases, respectively (Fig. 4A). The expression of IL-1β in the CAM incubated with non-CpG ODN was upregulated at 18 h post-treatment (p≤0.05).

Relative expression of various genes in the CAM; each graph
The expression of IFN regulatory factor (IRF)1 in the CAM isolated from LPS or R848 treated eggs was significantly increased at 3, 8, and 18 h post-treatment (Fig. 4B). The expression of IRF1 in the CAM after treatment with Pam3CSK4 was significantly increased at 3 h post-treatment, while the expression of this gene was upregulated at 18 h post-treatment in the group treated with CpG ODN 1826 (Fig. 4B). The expression of IRF7 in the CAM after treatment with either LPS or R848 was significantly increased at 3, 8, and 18 h post-treatment (Fig. 4C). In addition, the expression of IRF7 by the CAM treated with Pam3CSK4 was significantly upregulated at 3 and 8 h post-treatment with six- and twofold increases, respectively. The expression of IRF7 by the CAM treated with CpG ODN 1826 was significantly upregulated at 18 h post-treatment with a twofold increase (Fig. 4C).
TLR ligand treatment of embryonated eggs did not increase the expression of IFN-α or IFN-β by the CAM (Fig. 4D and E). The expression of IFN-α by the CAM isolated from embryos treated with LPS or R848 was significantly downregulated at 3 h post-treatment, both with a fourfold decrease (Fig. 4D).
LPS induced 20-, 8-, and 5-fold increases in IFN-γ expression by the CAM at 3, 8, and 18 h post-treatment, respectively (Fig. 4F). R848 induced significant expression of IFN-γ at 8 and 18 h post-treatment with 13- and 2-fold increases, respectively. The expression of IFN-γ by the CAM isolated from eggs incubated with CpG ODN 1826 was significantly increased (five fold) compared to untreated eggs at 18 h post-treatment, while Pam3CSK4 induced a three fold increase in IFN-γ expression at 8 h post-treatment (Fig. 4F).
The expression of 2′-5′ OAS, an ISG, by the CAM isolated from embryonated eggs stimulated with LPS, Pam3CSK4, and R848 was upregulated at 3 h post-treatment with 28-, 4-, and 4-fold increases, respectively (p≤0.05; Fig. 4G). The expression of 2′-5′ OAS by the CAM incubated with LPS and R848 was significantly increased at 8 h of incubation compared to untreated cells, with 74- and 8-fold increases, respectively. OAS expression by the CAM incubated with R848 was also significantly upregulated (49-fold) at 18 h post-treatment (Fig. 4G).
The expression of IFN-induced transmembrane protein (IFITM)5, another ISG, in the CAM isolated from embryonated eggs stimulated with LPS or Pam3CSK4 was upregulated at 3 h post-treatment both with approximately two fold increases (p≤0.05; Fig. 4H). Furthermore, IFITM5 expression was upregulated in the CAM stimulated with LPS and R848 at 8 h post-treatment with 3-, 1.5-, and 2-fold increases, respectively. The expression of IFITM5 by the CAM incubated with non-CpG was upregulated at 8 and 18 h post-treatment with 1.4- and 1.7-fold increases, respectively (p≤0.05; Fig. 4H). The expression of IFITM5 by the CAM incubated with LPS and R848 was significantly increased at 18 h of incubation compared to the untreated control group, both with approximately a twofold increase (p≤0.05, Fig. 4H).
The expression of inducible nitric oxide synthase (iNOS) by the CAM isolated from embryos treated with LPS and Pam3CSK4 was significantly increased at 3 h post-treatment by 13- and 10-fold, respectively (Fig. 4I). Additionally, a significant increase in iNOS expression was also observed with LPS and R848 treatment groups at 8 h post-treatment with 8- and 562-fold changes, respectively (Fig. 4I).
Discussion
In the present study, an in ovo model was established to study the effects of TLR ligands on AIV replication in embryonated chicken eggs. In 1957, Isaacs and Lindenmann discovered that the CAM is able to produce interfering agents, which inhibit the replication of influenza virus (19). Later studies showed that the CAM expresses some cytokines, including IL-1β, IL-8, IL-10, and IFN-γ, in response to infection with pathogens, such as Candida albicans and Aspergillus fumigatus (20,21). However, there has been no information on the response of the CAM to TLR ligands. This study examined whether TLR ligands administered at various doses and times pre- and post-infection could interfere with the replication of a H4N6 AIV. The results demonstrated that TLR2, 4, 7, and 21 are expressed by the CAM, and TLR ligand treatment of embryonated eggs induced antiviral and pro-inflammatory cytokine gene expression and reduced viral titer, suggesting that the CAM has the potential to recognize PAMPs.
It was discovered that the use of the optimal dose of TLR ligands was an important factor in their ability to inhibit AIV replication in the embryonated eggs without causing high levels of mortality. This was demonstrated with the fact that administering low amounts of the TLR2, 7, and 21 ligands did not reduce the influenza virus titer in embryonated eggs, while administration of high doses of these ligands caused high mortality in embryos. Therefore, an optimal dose of LPS, Pam3CSK4, R848, and CpG ODN was used for further studies. The lack of response from the low dose may be due to rapid degradation of some ligands, especially CpG ODN, in the allantoic fluid. Further studies are required to determine the toxicity and pharmacokinetics of these ligands to have a better understanding of the metabolism of these ligands in embryonated eggs.
The timing of TLR ligand administration was also a critical factor highlighted in this study. Consistent with previous studies, treatment of embryonated eggs with LPS prior to infection with AIV reduced AIV titer (35,36). The source of the LPS had a significant impact on the timing required to limit viral titers. Treatment of embryonated eggs with LPS from E. coli O26:B6 as early as 72 h prior to infection and as late as 24 h after infection was able to reduce AIV titer, while LPS from E. coli O111:O4 reduced virus titers more effectively at the time of infection or up to 36 h after infection. Distinct TLR-mediated signaling following stimulation of TLR4 with different sources of LPSs in mice and human has been previously reported (53). In addition, earlier study reported a divergence in the ability of different types of LPSs to induce antiviral responses in mice against influenza virus (36). In fact, TLR4 is able to respond to distinct types of Lipid A, the lipid component of LPS, affecting the magnitude of responses following TLR4 activation resulting in the production of different cytokines (28,31). As a result, there is a possibility that different sources of LPS may induce different cytokines and ISGs at different time points that reduce AIV replication in embryonated eggs. In contrast, the other TLR ligands, including Pam3CSK4, R848, and CpG ODN, only reduced virus titer when they were administrated prior to infection. The transient responses generated by Pam3CSK4, R848, and CpG ODN may be due to the different profile of induced responses or could be related to differences in the bioavailability, half-life, and clearance of the different TLR ligands. In fact, serum enzymes increase TLR ligand metabolism and decrease their half-life (10). For example, CpG ODN or single-stranded RNA (ssRNA) can be rapidly degraded when exposed to serum, and this may explain, to some extent, the duration of response after administration of these ligands. Furthermore, the administration of TLR ligands prior to infection may provide sufficient time to induce antiviral responses, including IFN and ISG and non-ISG proteins. In fact, it was observed that treatment with the various ligands increased the expression of a number of genes associated with antiviral responses. Although not measured directly, it is known from other studies that these and other antiviral factors can block viruses in different stages of their replication cycle (14). Conversely, the treatment of embryos with TLR ligands, such as LPS E. coli O26:B6, post-infection could induce ISGs that interfere with the translation, assembly, and release of AIV (14,17).
Following embryonic treatment with LPS, Pam3CSK4, and R848, the expression of IL-1β was increased in the CAM. The induction of pro-inflammatory cytokines in the CAM may recruit other cells, such as macrophages, to the site of virus replication. Recruited chicken macrophages in the CAM may be the source of iNOS in this membrane. It has been demonstrated that NO inhibits the replication of some viruses, for example, human herpes simplex virus in mouse embryonic fibroblasts, infectious laryngotracheitis virus in chicken macrophages, and AIV in the early stages of the virus life cycle (16,27,32). The TLR signaling network and subsequent innate responses rely on transcription factors such as IRF. The results presented here demonstrated that expression of IRF1 and IRF7 was significantly upregulated in the CAM following treatment of embryonated eggs with the various TLR ligands. Downstream IFNs and ISGs are then expressed to inhibit or restrict viral replication. It has been shown that TLR ligands induce the expression of type I IFNs in chickens (30,42,44,45). However, in the present study, TLR ligands did not cause a significant increase in type I IFNs but did induce the expression of some ISGs, such as OAS, IFITM5, and iNOS.
There is a possibility that type I IFNs might be increased at later time points that were not examined in this study. Moreover, alternative activation mechanisms may exist for subsets of ISGs following TLR stimulation. In spite of the lack of type I IFN expression, the expression of IFN-γ was upregulated by all of the TLR ligands. The direct antiviral activity of IFN-γ against AIV in chickens has not been characterized. However, IFN-γ has a wide range of activities, such as stimulation of macrophages and natural killer cells. In addition, IFN-γ induces NO production and upregulates some ISGs such as 2′-5′ OAS, dsRNA-activated protein kinase, and RNaseL (18,33,42,46). As a result, IFN-γ may inhibit the replication of AIV in embryonated eggs through the induction of NO production and upregulation of ISGs that interfere with AIV replication. Examination of genes further downstream in the TLR pathway demonstrated that Pam3CSK4, LPS, and R848 induced the expression of IFITM5 and 2′-5′ OAS in the CAM. The IFITM family inhibits the replication of influenza virus by disrupting the fusion of the viral and cellular membranes (8). Although, CpG ODN induces the expression of 2′-5′ OAS in chicken spleen and lung (42), CpG ODN 1826 did not induce 2′-5′ OAS in the CAM. This difference may be because of the type of cells present in the CAM (29). Overall, the expression of the aforementioned genes indicates the potency of TLR ligands in the induction of antiviral responses against AIV in embryonated eggs.
TLR ligands induce the expression of genes downstream in the TLR pathway via common adaptors and transcriptional factors (23). In spite of the similarities in the signaling pathways, differences in gene expression and reduction of viral titers were observed, most notably for LPS. Previously, it has been shown that LPS induces a wider range of genes compared to CpG ODN. In fact, LPS is able to induce more than 100 specific genes in mouse macrophages in addition to the genes that are commonly induced by CpG ODN and LPS (13). Similarly, with the nine genes examined in this study, it was observed that LPS increased the expression of more genes and to a greater extent than CpG ODN. In addition, there is a possibility that other genes that were not examined in this study may have longer lasting responses following LPS treatment. Regardless of the mechanisms, the ability of LPS from E. coli O26:B6 to reduce viral titers for a longer duration of time is of importance and could have some practical implications. Further studies are required to evaluate the stability of TLR ligands in the allantoic fluid. Several factors including pH and presence of enzymes in the allantoic fluid can influence stability of TLR ligands. LPS may be more stable in this condition compared to other ligands or may induce different antiviral mechanisms.
In conclusion, the findings of the present study suggest that TLR ligands when administered in ovo can interfere with the replication of AIV. The timing of administration of the TLR ligands strongly influenced the ability of the embryonated eggs to limit viral replication, such that only LPS reduced viral titers when administered at the same time as AIV or post-viral infection. Furthermore, all TLR ligand treatments reduced viral titers, although LPS did so to the largest extent. Overall, the results of this study confirmed the utility of the in ovo route for assessing the potency of TLR ligands as antiviral agents.
Footnotes
Acknowledgments
Funding for this study was provided by the Ontario Ministry of Agriculture, Food, and Rural Affairs (OMAFRA). Neda Barjesteh is a recipient of an OMAFRA scholarship. We would like to thank Mehdi Emam for statistical analysis and Michael St. Paul for his expertise.
Author Disclosure Statement
No competing financial interests exist.