
Abstract
JC Polyomavirus (JCPyV) is a widespread polyomavirus that usually resides latently in its host. As reactivation of the virus upon immune-modulating conditions holds serious risk, it is of importance to properly determine who is infected with this virus. Assessment of infection with JCPyV currently is based on the detection of antibodies against the major capsid protein VP1. However, specific antibodies against the peptide JCPyV_VP2_167-15mer have been shown to hold potential as a novel serological marker for infection with JCPyV. We have immunized rabbits with this peptide and the resulting hyperimmune serum was further characterized by detailed epitope mapping. The results demonstrated that the rabbit immune response is polyclonal in nature, recognizing two different epitopes in the 15-mer peptide. The strongest epitope consisted of L173PALTSQEI181, while a second moderate epitope consisted of D171DLPALT177. While some of the essential amino acid residues are the same as the ones for human plasma samples (P174, L176), some others are different. L173, T177, and I181 are essential for the rabbit hyperimmune serum, but not for human plasma samples, while E180 was essential for the human plasma samples and not for the rabbit hyperimmune serum. In conclusion, we generated polyclonal rabbit antibodies with strong reactivity against JCPyV_VP2_167-15mer recognizing at least two different epitopes in this peptide.
Introduction
J
Materials and Methods
Ethics statement
For the rabbit immunizations, all animals were handled in strict accordance with good animal practice as defined by the Belgian National Animal Welfare Regulations and in compliance with the Federation of European Laboratory Animal Science Associations (FELASA) requirements. FELASA (
Generation of rabbit polyclonal antibodies to JCPyV_VP2_167-15mer
Antigens were prepared by Eurogentec by coupling synthetic peptide JCPyV_VP2_167-15mer to keyhole limpet hemocyanin (KLH)-conjugated glutathione (JCV VP2-EP121567-KLH-GLUTA). This antigen was used for immunization of a specific pathogen-free rabbit (SB1538 and SB1539). The rabbit was immunized intramuscularly with a primary immunization supplemented with Freund's complete adjuvant followed by booster immunizations at 3, 7, and 11 weeks together with Freund's incomplete adjuvant. The immunization was performed by Eurogentec. Preimmune serum was collected on the day of the primary immunization (PPI), a small bleed was collected 10 days after the first booster immunization (PP), and a large bleed (GP) was collected 10 days after the second booster immunization. A final bleed (SAB) was collected 10 days after the third booster immunization.
Affinity purification of polyclonal antibodies
Affinity purification was performed by Eurogentec by coupling 10 mg synthetic peptide JCPyV_VP2_167-15mer to 2 mL Toyopearl AF-Amino-650 affinity resin. 50 mL final bleed serum was applied to a column containing the peptide-coupled resin. After a washing step with phosphate-buffered saline (PBS), specific antibodies were eluted with 100 mM glycine, pH 2.5 and diluted in PBS 1×, 0.01% thimerosal, and 0.1% bovine serum albumin. Antibody purity was verified and confirmed using Bioanalyzer (Agilent).
Peptide serology assays
For the determination of peptide-specific serum antibody levels, peptide ELISA was developed and set up as follows. Ninety-six-well flat bottom plates (Nunc C96 Maxisorp; VWR) were coated with 100 μL of 1 μg/mL Streptavidin from Streptomyces avidinii (Sigma) in Dulbecco's PBS without calcium and magnesium (Gibco). The plates were incubated at 4°C for 16 h. The plates were rinsed once with 200 μL PBS+0.05% Tween-20 (washing buffer) and blocked for 1 h at 37°C with 200 μL of 10-fold diluted Blocker Casein (Pierce) in PBS (blocking solution). Upon removal of the blocking solution, plates were incubated with continuous shaking for 2 h at room temperature with 100 μL of the selected biotinylated peptide, which were diluted at 1 μg/mL in blocking solution. The plates were rinsed three times with washing buffer, thereby eliminating unbound peptide. Then, the different wells were covered with 100 μL of rabbit serum samples diluted in blocking solution, as indicated. Each sample was analyzed in duplicate. In control wells, blocking solution was added instead. The plate was incubated at 37°C for 1 h. After incubation, a new triple rinsing cycle was repeated as described above. Then, the secondary antibody solution was added to each well. The solution contained a goat anti-rabbit IgG peroxidase conjugate (Santa Cruz), diluted 1:5,000 in blocking solution. The reaction mixture was incubated at 37°C for 1 h. At the end of the incubation period, the plates were rinsed three times with washing buffer and treated with 100 μL SureBlue™ TMB Microwell Peroxidase Substrate (KPL). After 10 min of incubation, the colorimetric reaction was stopped with 100 μL 1 N HCl. The plate was then read by the spectrophotometer (SpectraMax; Molecular Devices) at a wavelength (λ) of 450 nm. Samples were considered positive if OD values were higher than twofold the OD value of the blank sample (i.e., log2 test/ctrl>1).
Results and Discussion
A rabbit was immunized with JCPyV_VP2_167-15mer. A dilution series was made of the rabbit PPI, preimmune serum, PP, small bleed, GP, large bleed, SAB, final bleed, and PA, purified antibodies. All dilutions were tested for reactivity to JCPyV_VP2_167-15mer. Figure 1 shows that hyperimmune sera from the immunized rabbit have high reactivity to the peptide while no response could be detected in the preimmune serum. Also the purified antibodies have high antibody reactivity to the peptide. Based on these data, a 2,000-fold dilution of small bleed (PP) was chosen for further experiments.
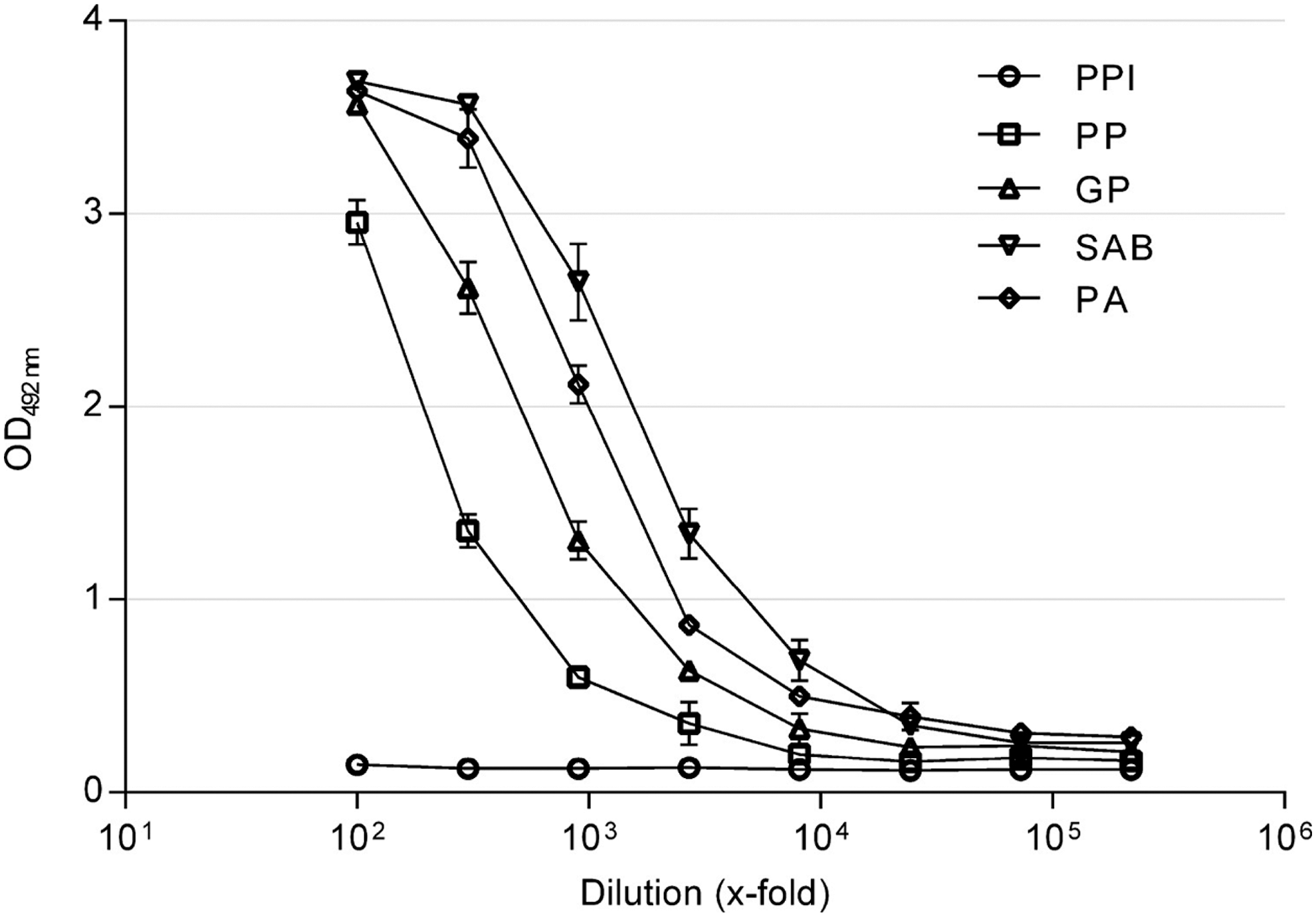
Immunoreactivity against JCPyV_VP2_167-15mer of several dilutions of rabbit preimmune serum (PPI), hyperimmune sera upon immunization with JCPyV_VP2_167-15mer peptide (large bleed [GP]; affinity-purified antibodies [PA]; small bleed [PP]; final bleed [SAB]). All dilutions were analyzed in triplicate; mean absorbance and standard deviation are reported. CV% of corresponding OD values was <20%.
To find the more precise location of the epitope that is recognized by the rabbit hyperimmune serum, epitope mapping experiments were performed, similar to what was done previously using human plasma samples (9). For this investigation, a peptide ELISA was employed to test the rabbit hyperimmune serum using different specifically designed synthetic peptides. A series of 7-mers, 9-mers, 11-mers, and 13-mers, overlapping with all but one residue was tested (Fig. 2A). This test showed that two different epitopes in the 15-mer appear to be recognized by the hyperimmune serum. On the one hand, the sequence LPALTSQEI was capable of retaining high immunoreactivity, but on the other hand also the peptide DDLPALT showed some moderate but consistent immunoreactivity, which is in contrast to the situation in human plasma samples, where only the LPALTSQEI epitope is recognized. It must be noted, however, that only three human plasma samples were used for these epitope mapping studies. As such, it cannot be excluded that the anti-JCPyV_VP2_167-15mer antibodies from other individuals also recognize more than one epitope in this peptide.
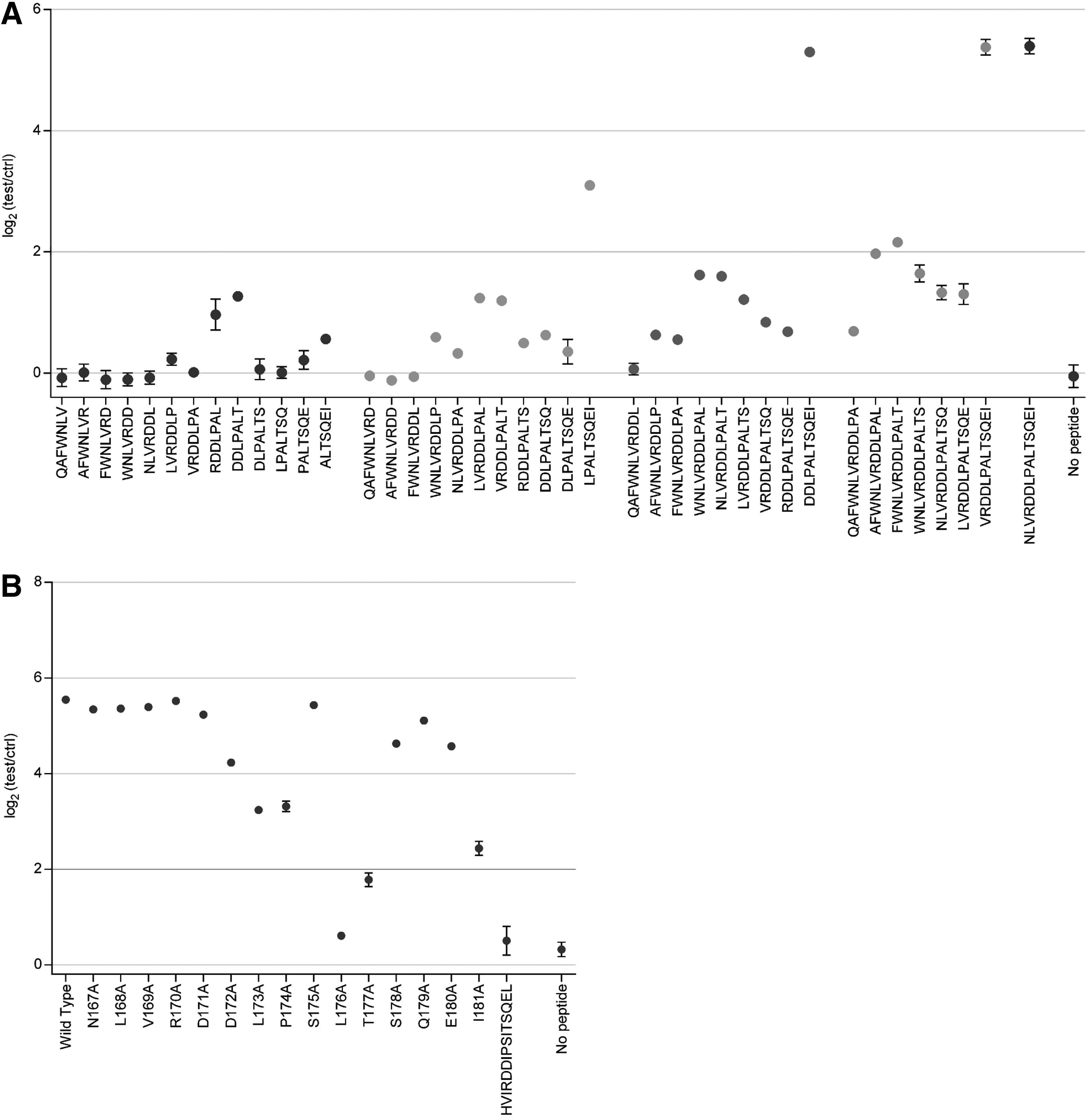
In a second approach each of the amino acids in JCPyV_VP2_167-15mer was replaced individually by alanine (alanine walking) (Fig. 2B). Using this approach, the contribution to the immunoreactivity of the peptide of each specific residue is determined (3,4). The data obtained using these peptides in the peptide ELISA indicate that five residues in JCPyV_VP2_167-15mer are important for antibody recognition: L173, P174, L176, T177, and I181. Replacement of any of these residues by alanine resulted in at least partial elimination of immunoreactivity of the peptide. While some of these residues are the same as the ones identified using human plasma samples (P174, L176), some of the essential residues are different. L173, T177, and I181 are essential for the rabbit hyperimmune serum, but not for human plasma samples, while E180 was essential for the human plasma samples and not for the rabbit hyperimmune serum. Most single residue replacements to alanine did not completely eliminate the immunoreactivity of the peptide, indicating again that the hyperimmune serum is polyclonal. Indeed, replacement of essential residues in one of the two epitopes (e.g., T177A) resulted in a response similar to the minimal DDLPALT epitope, indicating that in this case only the latter epitope is recognized. To determine the contribution of the essential amino acid residues to each of both epitopes, one would need to perform alanine walking experiments on both minimal epitopes separately (LPALTSQEI and DDLPALT).
As this polyclonal nature might also impact the specificity of the hyperimmune serum toward, for example, BKPyV_VP2, we also investigated the immunoreactivity of the rabbit hyperimmune serum against the heterologous BKPyV peptide (HVIRDDIPSITSQEL). There was no reactivity toward this peptide, indicating the rabbit hyperimmune serum has no cross-reactivity with BKPyV_VP2 (Fig. 2B).
Taken together, these data suggest that the immune response upon immunization of a rabbit with JCPyV_VP2_167-15mer appears to be polyclonal of nature. This rabbit hyperimmune serum is now available for further investigation and its use in immunohistochemical staining or western blot analysis of JCPyV_VP2 protein in human samples might be evaluated. However, one should keep in mind that these antibodies might display some minimal nonspecific binding to other proteins than JCPyV_VP2, especially to other polyomavirus VP2 proteins. This hyperimmune serum could potentially also be used as positive control in an immunoassay for detection of antibodies directed against JCPyV_VP2_167-15mer in human serum samples.
An important consequence of the polyclonal nature of the immune response in rabbits is the question whether this could also be the case in humans. Our previous work only identified one single epitope in the plasma of three subjects, but it cannot be excluded that some subjects generate different antibodies upon JCPyV infection, each recognizing different residues in the peptide upon presentation by an antigen-presenting cell. It might even be possible that for some subjects the antibodies produced upon JCPyV infection recognize a single but different epitope in the JCPyV_VP2_167-15mer. In this respect, it has been recognized earlier that immunization of rabbits does not always result in antibodies with exactly the same epitope. Immunization of multiple animals with the same antigen resulted in similar but not identical epitopes (6). An important aspect that should also be considered when comparing the rabbit antibody response described in this study with the antibody response observed in humans upon JCPyV infection is the fact that rabbits were immunized with a peptide-protein conjugate. This conjugation process results in the peptide being presented to the immune system in a different context compared to the peptide being part of the viral capsid.
Conclusion
In this study we have generated a rabbit hyperimmune serum to JCPyV_VP2_167-15mer and determined the minimal epitope and essential amino acid residues in this epitope. We found that at least two epitopes are recognized and that the epitope with the highest immunoreactivity, D171DLPALTSQEI181, was similar but not identical to the epitope that was found previously in human plasma samples upon exposure to intact JCPyV.
Footnotes
Acknowledgments
This work was funded by a grant from the Flemish Agency for Innovation by Science and Technology (IWT). This organization played no role in the writing of the article and in the decision to submit the article for publication. The authors also would like to thank the following persons for their contribution in useful discussions: Tom Van Loy, Liesbeth Van Wesenbeeck, Hanne Meeuws, Bieke Van Dorst, and Kristiane Schmidt (Janssen Diagnostics, Belgium). O.L. and L.J.S. designed the study setup. O.L. performed the experimental work. O.L. performed the analysis of the data. L.T. and L.J.S. contributed to revising the article critically for important intellectual content and gave final approval of the version. All authors read and approved the final article.
Author Disclosure Statement
Authors are current employees of Janssen Diagnostics BVBA or Janssen Research and Development, both being Johnson and Johnson Companies and may own stock or stock options in the company.