
Abstract
Influenza A viruses (IAV) induce cytokine storm and host's intracellular redox imbalance to ensure continuous replication and survival, leading to severe immunopathology and death. The unpredictability of broad-spectrum vaccines, the emergence of drug-resistant and/or more virulent strains, the prevalence of the amantadane-resistant IAV, and the prohibitive cost of available drugs especially in resource-poor countries necessitate exploring drugs with novel action mechanisms as anti-influenza agents. This study presents the protective role of kolaviron (KV), a natural antioxidant and anti-inflammatory agent from Garcinia kola seeds, on BALB/c mice challenged with influenza A/Perth/H3N2/16/09 (Pr/H3N2) virus. KV at 400 mg/kg was administered orally to groups of BALB/c mice for 3 days, 3 h, and 1 h prior to infection with 1LD50 or 3LD50 (14-day study) and 5LD50 (6-day study) Pr/H3N2. Pr/H3N2 in the lungs was detected by hemagglutination assay, while oxidative stress and inflammatory biomarkers were assayed in both lungs and liver. Infected mice treated with KV progressively increased in weight with minimal mortality. Single-dose administration of KV at 1 h or 3 h before viral challenge and 3 days pretreatment improved lung aeration and reduced lung consolidation as well as inflammatory cells infiltration in a way that had minimal impact on viral clearance, but attenuated myeloperoxidase activity and nitric oxide production via priming of reduced glutathione levels, thus enhancing the preservation of function in the lungs and liver. This study suggests that KV may be effective for delaying the development of clinical symptoms of influenza virus, and this may be through a mechanism unrelated to those deployed by the existing anti-influenza drugs but closely associated to its antioxidant and immunomodulatory properties.
Introduction
H
The pathology of IAV in humans is consistently and almost always characterized by complex biological phenomena, including unabated production of pro-inflammatory cytokines and chemokines, resulting in predominant pulmonary hyperemia, progressive pneumonia, and loss of lung function, partly attributed to direct viral multiplication-mediated apoptosis (13,31,40). Culminating lung failure in the host with virulent strain infection or severe clinical outcome has also been attributed to the downstream apoptotic effect of the sustained pro-inflammatory host response and exacerbated tissue remodeling induced fibrosis following influenza infection (13). Meanwhile, conventional therapeutic agents that target IAV's life cycle, including neuraminidase inhibitors and adamantanes, only perform optimally and effectively when administered shortly after the manifestation of disease symptoms, before the onset of the cytokine storm (15). Evidences also abound that evolving pandemic strains outsmart the annually predicted vaccine that depends solely on informed and educated guesses from the prevailing isolates and circulating strains, thus necessitating a new antiviral bullet (5,29). The effectiveness of current approaches to treatment and prevention of influenza have also been limited by factors such as the prevalence of the adamantane-resistant influenza viruses, the prohibitive cost of available drugs especially in resource-poor countries, the unpredictability of vaccine availability, and the time lag between vaccine development (25,48).
Thus, a recent paradigm shift includes the search and quest for antiviral, redox regulator and/or anti-inflammatory strategies that could either solely emerge as new anti-influenza or adjunct therapy combined with the existing arsenal to reduce the severity and complications of influenza infections (1,5,14,18,45 –48). Data from a number of studies suggest that the use of oxygen free radicals as targets may provide an approach to the amelioration of the pathogenicity caused by influenza virus infections (15,47), and several studies have reported the anti-influenza activity of medicinally potent and disease-preventing plant polyphenols and flavonoids with antioxidant properties (9,17,24,42,47,48).
Kolaviron (KV; Fig. 1), a fraction of the defatted methanol extract of Garcinia kola seeds, has been shown to exhibit potent radical scavenging properties, metal chelating activities, and immunomodulating potential (3,19,20,22). Several studies have demonstrated the mechanism of chemoprevention of KV against numerous models of degenerative diseases to include modulation of responses to oxidative stress by stimulating phase 2 detoxification enzymes, thereby mitigating the oxidative damages to biomolecules, and downregulating NF-κB and AP-1 DNA binding activities, as well as iNOS and COX-2 expression at the molecular level (19,21,22). These latter molecular biomarkers have been strongly implicated in inflammation, immune responses, and especially immunopathology of influenza virus (4). The specific aims of this study were to investigate the protective potential of KV in influenza A/Perth/H3N2/16/09 virus-infected BALB/c mice using mortality, morbidity, and histopathology as end points, and to elucidate the responsible underscoring mechanism via the assessment of biochemical antioxidant and inflammation indices.
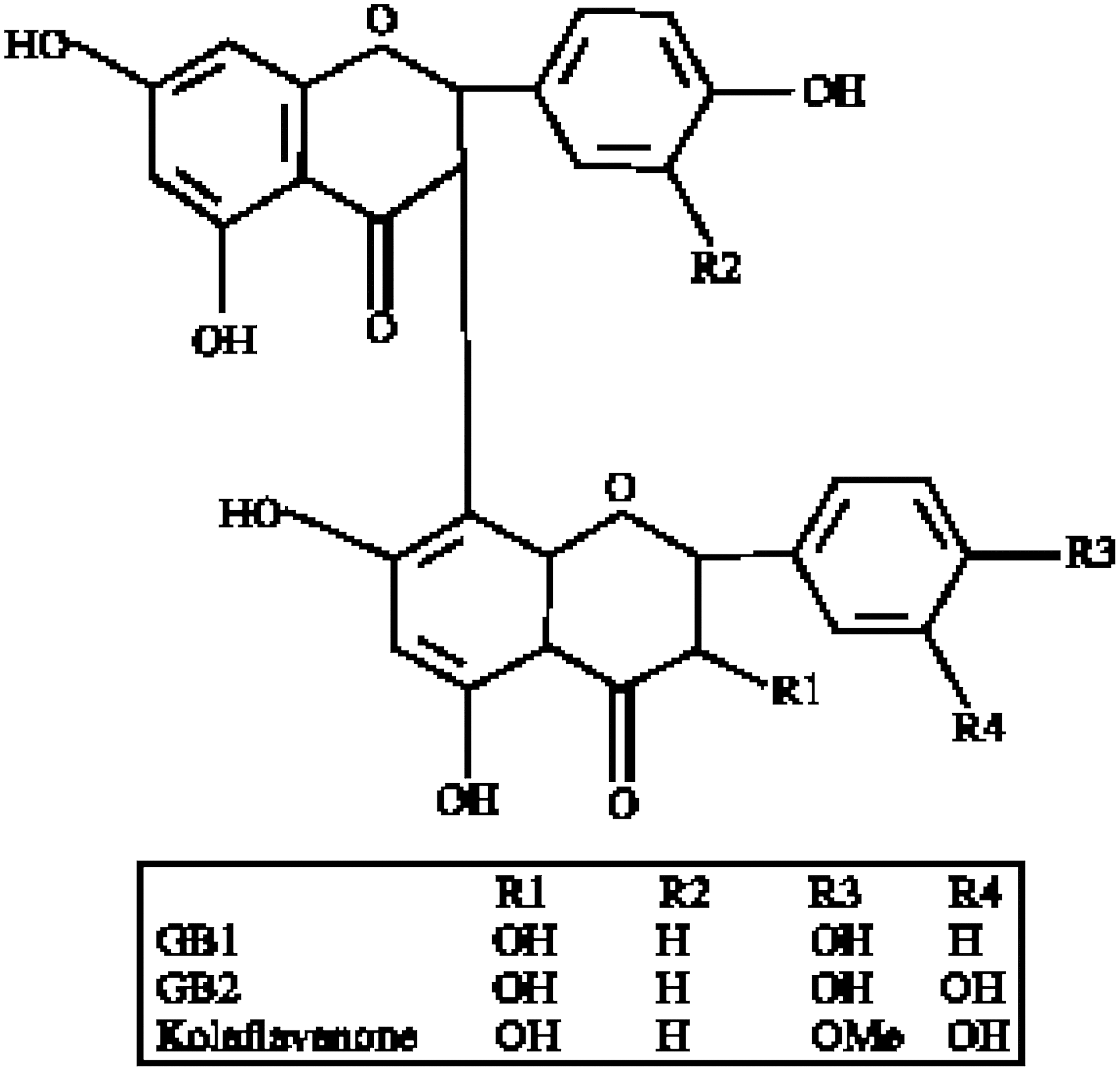
Structure of kolaviron (KV).
Materials and Methods
Plant material and extraction of KV
The test extract was prepared from Garcinia kola seeds. The seeds, obtained from Ile Ife, Nigeria, were peeled, sliced, air-dried, and pulverized. The extraction of KV (from Garcinia kola) was done according to the method of Iwu et al. (27) with some modifications from Farombi (20). Briefly, the pulverized seeds were defatted with petroleum ether and extracted with methanol. The extract was concentrated and diluted to twice its volume with distilled water and extracted with ethyl acetate (6×250 mL). The concentrated ethyl acetate fraction gave a yellow solid tagged KV (Fig. 1). KV was identified by direct comparison with 1H nuclear magnetic resonance (NMR), 13C NMR, and electron ionization (EI)-mass spectral results with previously published data (27). The purity and identity of KV was determined by thin-layer chromatography using silica gel GF 254-coated plates with a solvent mixture of methanol:chloroform at a ratio of 1:4 v/v. The purity of isolated KV was 96%.
Virus propagation
Well-characterized isolate of IAV, A/H3N2/Perth/16/09 (Pr/H3N2), was obtained from the WHO Influenza Reference Centre, Medical Research Council, London. The isolate was propagated in the chorioallantoic fluid of 10-day embryonated chicken eggs (obtained from Chi Farms Ltd., Ajanla Ibadan, Nigeria), subjected to hemagglutination assay (HA) for viral titer determination and later adapted in mice. The median lethal dose (LD50) of the harvested allantoic fluid and mouse-adapted lung homogenate was also determined after 14 days. Stock virus was aliquoted and stored at –80°C until use.
Protective effect of KV in A/H3N2/Perth/16/09 (1LD50 or 3LD50) challenged BALB/c mice: animal inoculation
This study was carried out according to the guidelines and approval of institutional animal care and use research ethics committee (approval number FVM/A.6/201402). BALB/c mice (10–13 g) were obtained from the breeding facility of the Department of Biochemistry, University of Ibadan, Nigeria. They were anesthetized with ketamine (10 mg/kg) intraperitoneally (i.p.) before 32 μL of mouse adapted Pr/H3N2 was instilled intranasally. Before viral inoculation, a 14-day study to determine the dose lethal to 50% of experimental animals was carried out, and LD50 was calculated in accordance with the stipulated guidelines of Reed and Muench (41). KV, a natural biflavonoid antioxidant and anti-inflammatory agent, extracted from the edible and masticatory seeds of Garcinia kola, was investigated for its potential morbidity-delaying and mortality-inhibiting roles at 400 mg/kg. KV, dissolved in dimethyl sulfoxide (DMSO) and diluted with corn oil, was administered orally to groups of mice (n=8–10) 1 h and 3 h prior to viral challenge with 1LD50 Pr/H3N2. Infected untreated mice received an equivalent volume of 2.5% DMSO in corn oil 1 h before viral inoculation as mock control. General condition, including weight loss, reduced activity, ruffled fur, difficulty breathing (tachypenea and labored respiration), diarrhea, and survival (mortality), were monitored daily for 14 days. An expanded protective study was also carried out to include consecutive treatment for 3 days before the mice were infected with a higher infectious dose of 3LD50 of the same virus, in addition to the previously mentioned schedule. Mice were sacrificed at 14 days post infection (dpi). Any mouse that showed signs of disease and weight loss of >25% of its body weight was humanely euthanized.
Postmortem pathological analyses with histopathology and hydrostatic lung test
Following mortality or at 14 dpi, lungs were collected and immediately processed for postmortem pathological analyses, including macropathological examination, hydrostatic lung test, which was performed to determine the degree of aeration, lung weight measurement to determine the severity of infection cum inflammation in the lungs, and histopathology. For histopathology, a portion of the lungs was collected and infused with 4% neutral-buffered formaldehyde and later stained with hematoxylin and eosin. The other portion was immediately frozen on dry ice, and stored at –80°C until analyzed for lung viral detection.
Pulmonary virus detection
The frozen lungs were later thawed, homogenized in 1 mL of cold phosphate-buffered saline (PBS), and clarified by centrifugation (2,200 g) at 4°C. Viral multiplication in the lungs was detected and quantified by inoculating 100 μL of lung homogenates into 9-day-old embryonated chicken eggs, three eggs per mouse, incubated at 37°C for 72 h, after which the allantoic fluids were harvested and assayed for hemagglutination with chicken red blood cells (RBC; 0.5%) in 96-well plates in duplicate. In HA assay, agglutination with chicken erythrocytes is the direct consequence of the interaction between the viral hemagglutinin and sialic acid receptors on the surface of the cells.
Oxidative stress, inflammation and immunopathology modulating effects of KV in BALB/c mice challenged with A/H3N2/Perth/16/09 (5LD50)
BALB/c mice weighing 13–15 g, six animals per group, were used for this study. BALB/c mice were pretreated with KV (400 mg/kg) at 1 h, 3 h, or 3 days before 32 μL of Pr/H3N2 (5LD50) was instilled. Infected untreated mice received equivalent volume of 2.5% DMSO in corn oil 1 h before viral inoculation as mock control. Mice were mildly anesthetized with ketamine (10 mg/kg; i.p.) prior to viral inoculation. General condition, including weight loss, reduced activity, ruffled fur, difficulty breathing (tachypenea and labored respiration), diarrhea, cyanosis, and survival rates (mortality), were monitored daily for 6 days. Any mouse that showed signs of disease and weight loss of >25% of its body weight was humanely euthanized. At 6 dpi, the animals were euthanized, and the lungs and liver were harvested, weighed, and processed for postmortem and pathological analyses, and homogenized.
Evaluation of biomarkers of inflammation and oxidative stress
Mice were euthanized on day 6 post challenge. Animal tissues (lungs and liver) were rinsed with 1.15% KCl/4°C solution. Then, 60% tissue homogenates were prepared with 0.1 M phosphate buffer (pH 7.4/37°C). Supernatants for biochemical assays were prepared after separation of the nuclei and mitochondrial fractions at 10,000 g/4°C 12 min from 60% tissue homogenate. Reduced glutathione (GSH) was determined according to Jollow et al. (28). Lipid peroxidation was determined as malondialdehyde (MDA) according to the procedures described by Varshney and Kale (49). The nitrite (NO2 –) level in the tissues was estimated as an index of nitric oxide (NO) production. Quantitation was based on the Griess reaction as described by Crespo et al. (12). Myeloperoxidase (MPO) activity, an indicator of polymorphonuclear leukocyte accumulation and activation, was determined by the method describe by Bradley et al. (6).
Hemagglutination inhibition properties of KV
The titer of the stock virus used for this assay was first determined in HA by the addition of 50 μL of 0.5% chicken RBC suspension in PBS to equal volume of twofold serially diluted stock virus in a 12×8 microtiter plate. The solution was gently mixed and incubated at room temperature for 30 min, and the end point, the last well in which a sufficient amount of the virus agglutinates the RBC, was determined. To determine the inhibitory properties of KV on the attachment of influenza virus to sialic acid–containing receptors, the stock virus was first diluted to 8HA/50 μL and then back-titrated. A total of 25 μL of 0.5% DMSO (vehicle control), KV (400 or 1,000 μg/mL), or PBS (viral control) was added to equal the volume of already dispensed 8HA/50 μL stock virus; the solution was incubated for 30 min at room temperature after which a suspension of 50 μL of chicken erythrocytes was added. The whole reaction mixture was incubated for another 30 min. The end point for hemagglutination inhibition in the KV-treated wells was the settling of RBC at the base of the V-bottom plate as observed by a microtiter plate reader. In another plate, hemagglutinnation inhibition of KV, at both concentrations, was investigated on the serially diluted stock suspension following the same procedure described above.
Statistical analysis
Values of quantitative data were expressed as mean±standard deviation. Comparison among groups was done by subjecting obtained data to the F-test (analysis of variance) using SPSS Statistics for Windows (version 20.0). Values were considered as statistically significant at a p-value of<0.05. The survival rate was analyzed by the Kaplan–Meier method, and its significance was evaluated by the original log–rank method.
Results
KV improves morbidity while mortality onset is delayed
The first experiment to have a glimpse of the protective potential of KV on influenza virus infection in BALB/c mice was carried out with a dose that was lethal to half of the mice population (1LD50), which was determined from a preliminary study. Mice were infected with A/H3N2/Perth/16/09 1 h or 3 h after a single dose pretreatment of KV at 400 mg/kg and observed for 14 dpi. Following the viral challenge, infected untreated mice lost weight appreciably (Fig. 2A and B) resulting in 50% mortality (Fig. 2C); nonetheless, the difference was not significant. KV treatment at 3 h prior to viral instillation conferred complete protection, while treatment at 1 h prior prolonged mortality onset with a mean day to death of 13.75 days (Fig. 2C and F). Though the virus was detected in the lungs of the treated mice (Fig. 2D), the weight of the lungs was significantly reduced (Fig. 2E). Figure 3 shows that the lungs of KV-treated mice were more aerated with patchy cellular infiltration and pulmonary inflammation.
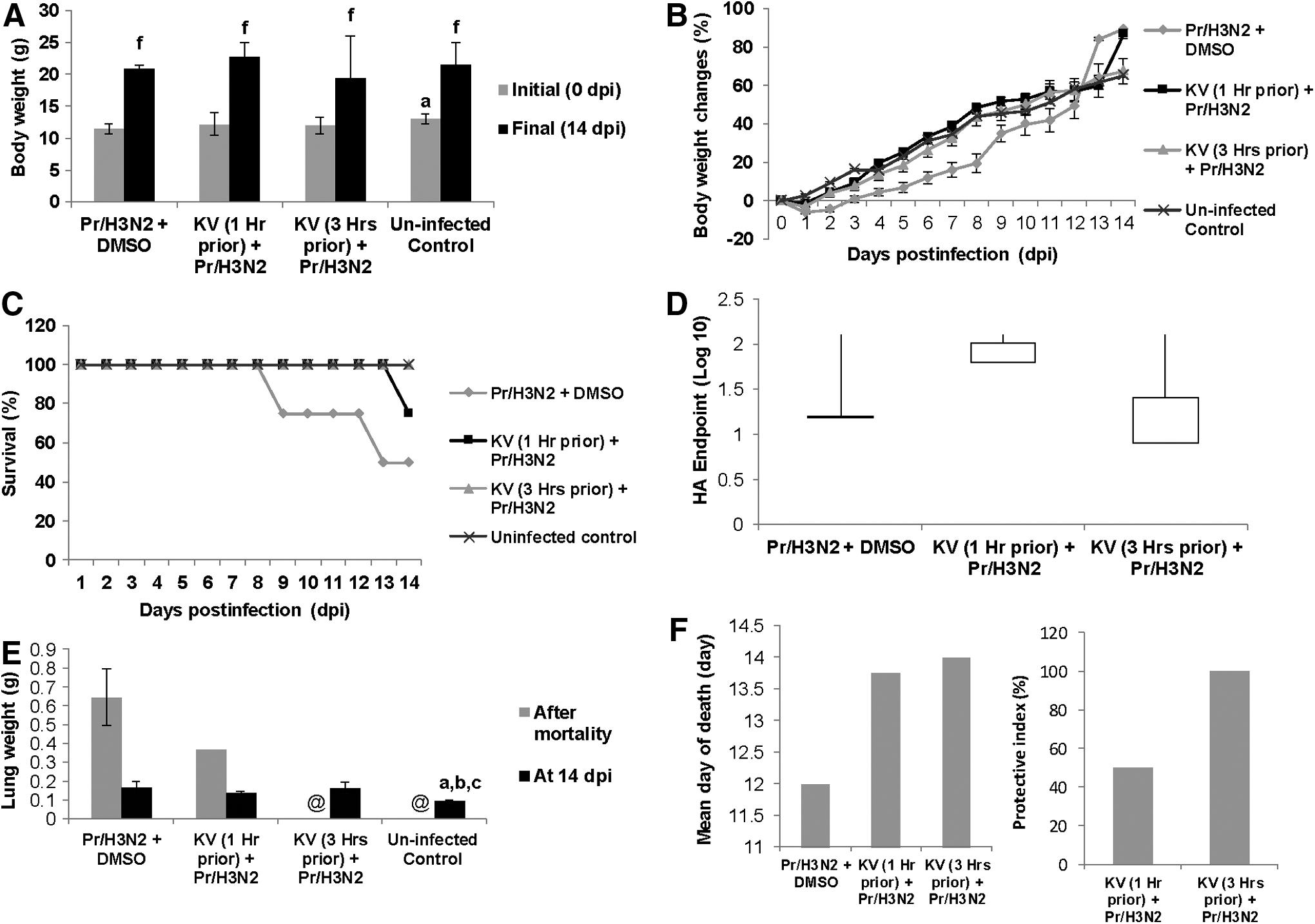
Pretreatment with KV protects mice against H3N2 influenza virus. BALB/c mice were pretreated with KV at 1 and 3 h before A/Perth/16/09 virus (1LD50) instillation, and monitored daily for body weight changes (

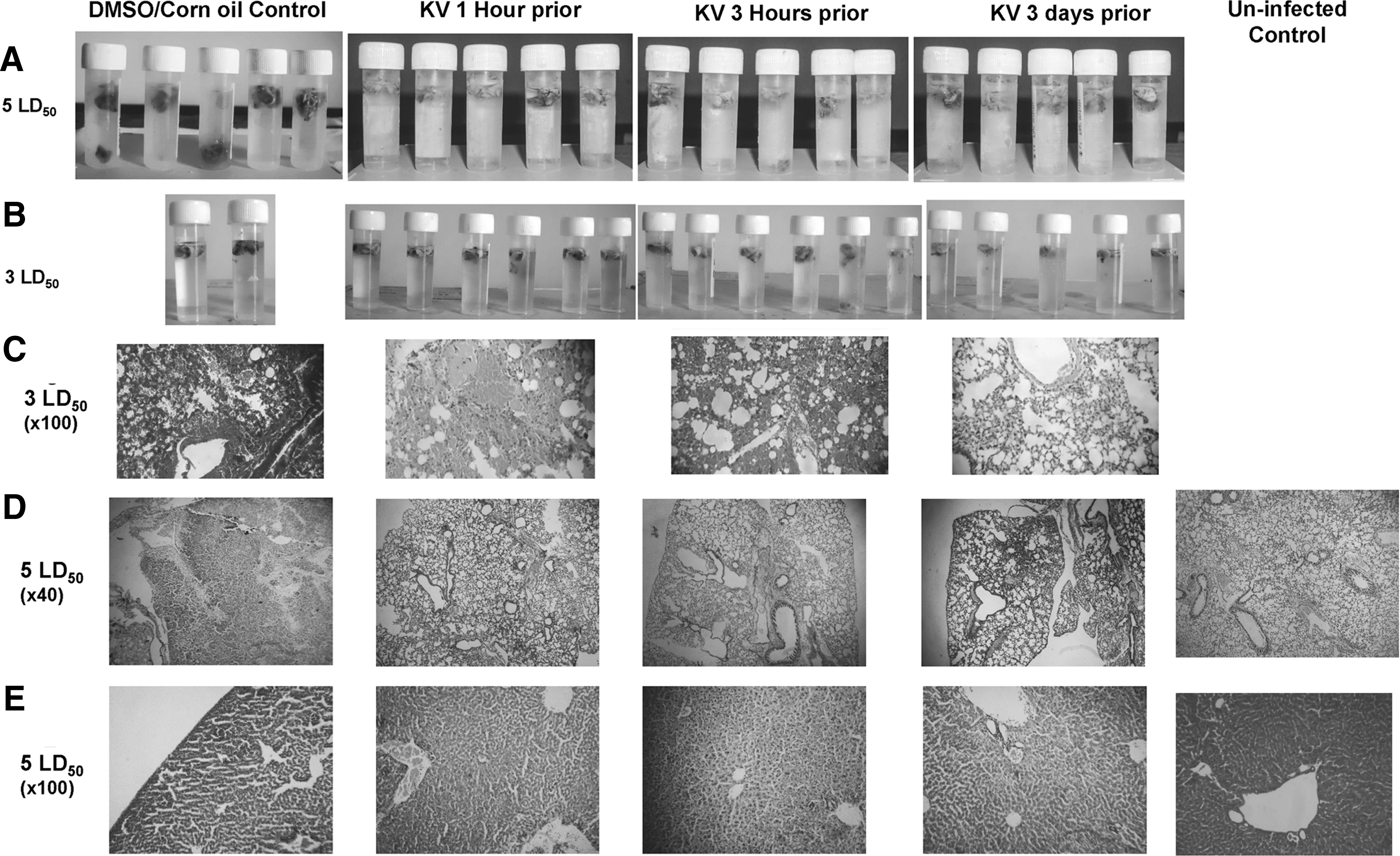
Gross pathology via hydrostatic lungs test (right panels) and histopathological changes (left panels) of mice infected with A/H3N2/Perth/16/09 (Pr/H3N2) and pretreated with or without KV. Gross pathology of the lungs of BALB/c mice pretreated with 400 mg/kg KV (1 h or 3 h) and later infected with 1LD50 Pr/H3N2. Control animals were mock-treated with corn oil solution of DMSO. At 14 days post infection, surviving mice were euthanized and lungs harvested, rinsed, and placed in phosphate-buffered saline (PBS) to determine the degree of aeration. For histology, lung sections were stained with hematoxylin and eosin (H&E).
With promising data in the previous study and the vindication that KV is a better chemopreventive agent when pretreated for a number of days in several chemopreventive models (21,23), another pretreatment regimen was included for 3 days, in addition to the aforementioned groups in the previous experiment. However, the dose of the influenza virus instilled was raised to three times the mean lethal dose (3LD50). A higher dose of the virus imposed a higher level of morbidity and mortality (Fig. 4A–C). Compared with the infected but not treated mice with subsequent significant 72% mortality, treated mice at different time points progressively increased in weight (Fig. 4A and B). Again, KV given 3 days prior conferred complete protection, while administration at 1 and 3 h before viral intranasal challenge resulted in an 80% protective index (Fig. 4C and F). In similitude, infective virions were also detected in the lung homogenate of the treated rats (Fig. 4D). However, consolidation of the lungs in the treated mice was minimal (Figs. 4E and 5B), and this could also be histologically traceable to the improved immunopathology characterized with reduced edematous alveolitis, necrotizing bronchiolitis, and patchy infiltration of inflammatory cells (Fig. 5C). Alveolar septa thickening and desquamation of bronchus epithelium were evident in the mock-treated mice (Fig. 5C).
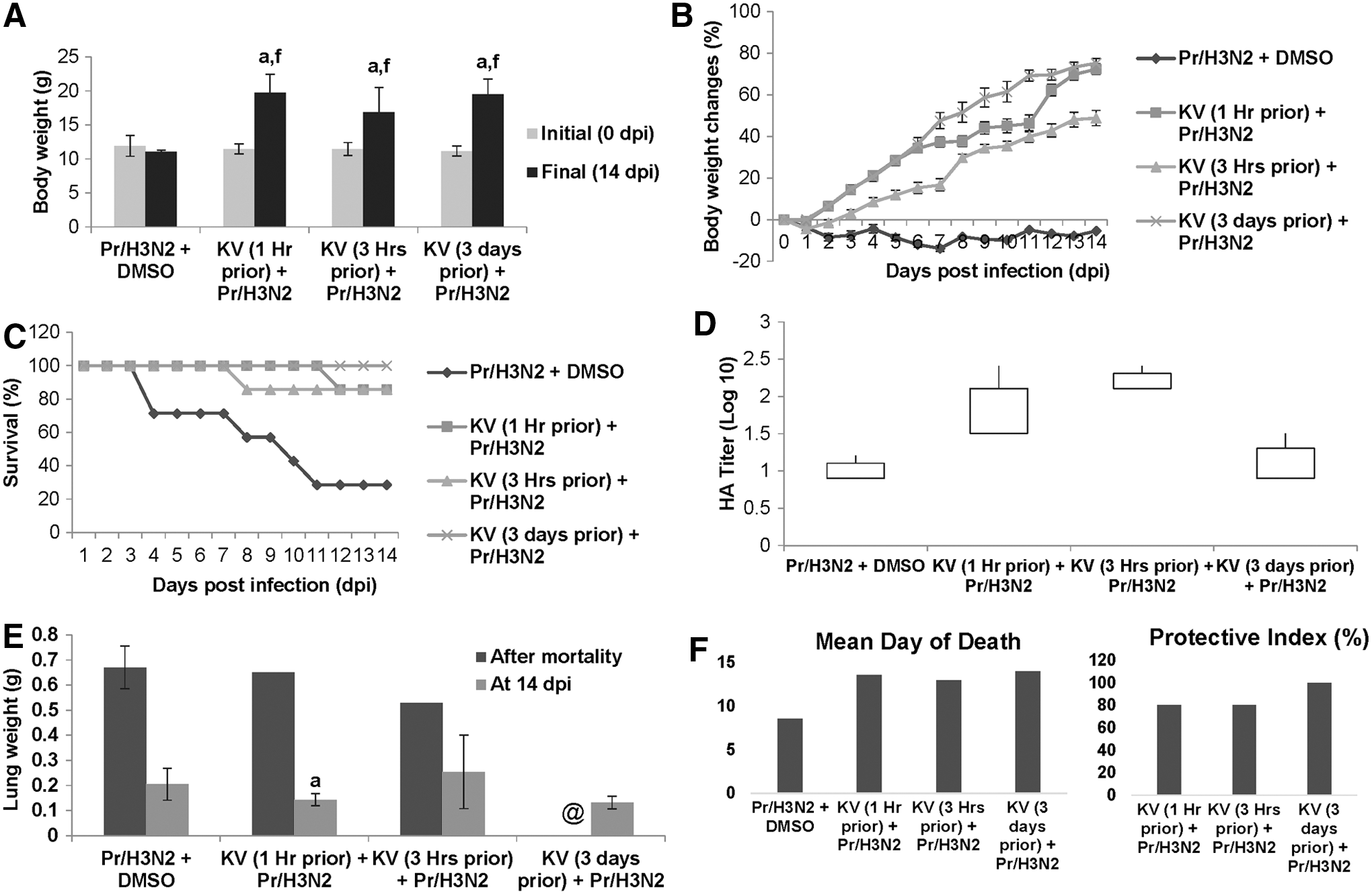
KV improves survival of mice infected with H3N2 influenza virus. BALB/c mice received KV (400 mg/kg) orally 1 h, 3 h, or 3 days before A/Perth/16/09 virus (3LD50) instillation and monitored daily for body weight changes (
Gross pathology via hydrostatic lungs test (
In an attempt to delineate the underlying mechanism of protection, a preliminary study was carried out to assess biochemically some biomarkers of oxidative stress and inflammation with a similar design but for the dose of influenza virus and study duration, which were pegged at 5LD50 and 6 days, respectively. Since it was observed in the previous study that KV may not mitigate viral replication but may impair mortality by improving morbidity via suppression of immunopathology, this particular study focused more on adducing biochemical explanations for the observed protection. Mock-treated mice increasingly lost weight to reach about 22% of the initial body weight by 6 days post-challenge (Fig. 6A and B). On the contrary, even at a remarkably higher dose, BALB/c mice pretreated with KV at 1 h, 3 h, or 3 days before Pr/H3N2 (5LD50) instillation showed a significant reduction in morbidity when compared with the untreated mice (Fig. 6A and B). Consolidation of the lungs in the KV-treated groups was likewise significantly reduced (Figs. 6C and 5A).
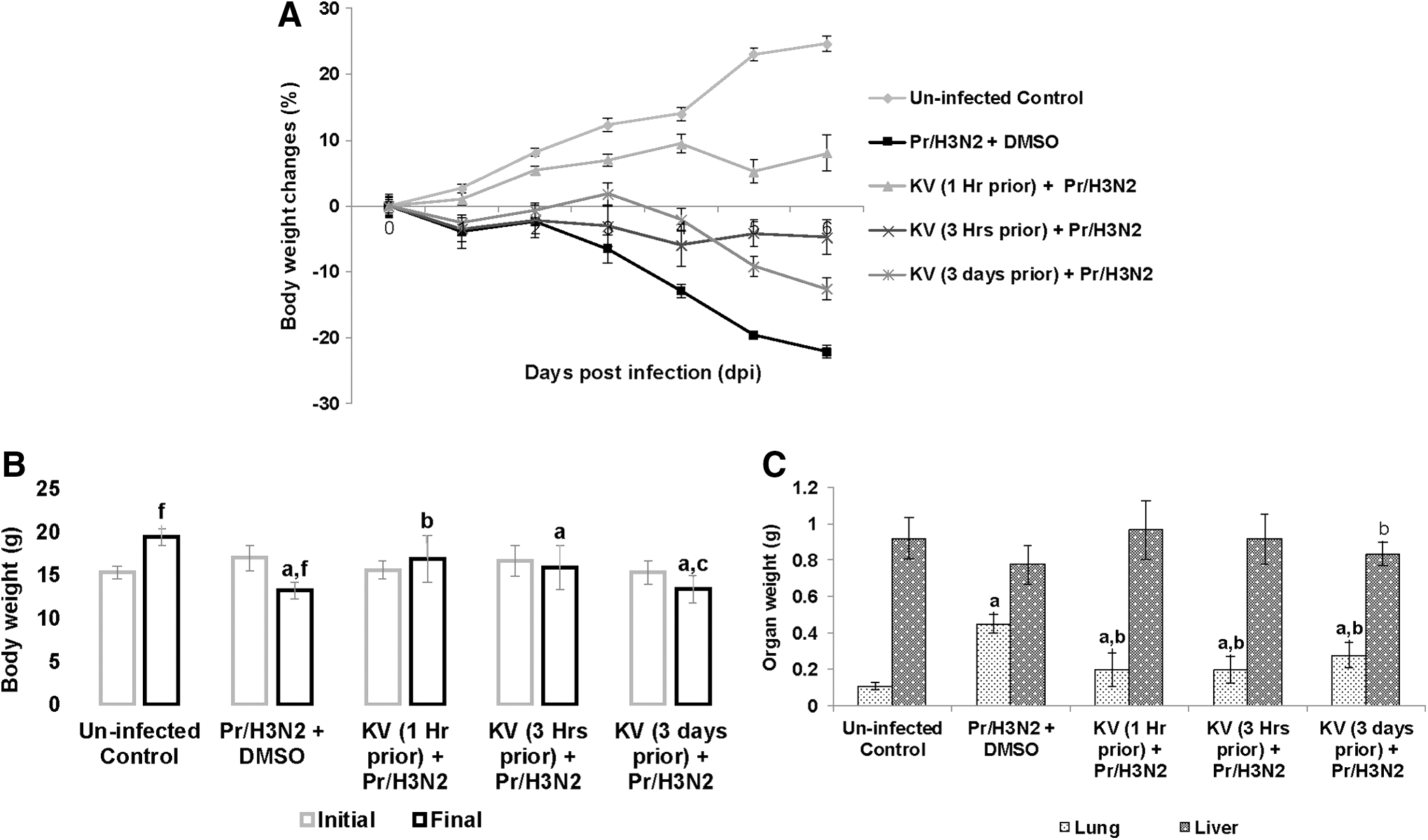
Daily body weight changes (
Pathological examinations showed that KV, at various administration time points, reduced pulmonary inflammation exclusively, increased lung aeration, and improved parenchyma integrity with patchy cellular infiltration (Fig. 5D). Interestingly, influenza virus–induced inflammation was felt at a distant extrapulmonary tissue. The liver tissues from the virus-challenged mice were pale with reduced weight, even though the difference was not statistically significant when compared with the KV-treated mice, except in the group of mice pretreated for 3 days (Fig. 6C). However, liver from BALB/c mice challenged with Pr/H3N2 appeared architecturally disrupted, with inflammatory cells occupying the portal vacuoles, while those from the pretreatment groups appeared structurally and functionally normal (Fig. 5E).
On study day 6, the infection of BALB/c mice with 5LD50 A/H3N2/Perth/16/09 significantly resulted in sustained myeloperoxidase activity, increased generation of nitric oxide (Fig. 7), and, unsurprisingly, soaring MDA levels (Fig. 8A) in both lungs and liver when compared with uninfected control mice. However, the GSH level was unaffected in the lungs but was significantly reduced in the liver (Fig. 8B). Oral administration of KV ameliorated the increase in the activity of myeloperoxidase (Fig. 7B), as well as levels of nitric oxide (Fig. 7A) and MDA (Fig. 8A) in the lungs, with marked amelioration observed in the group pretreated for 3 days. In the liver, pretreatment with KV reversed the induced myeloperoxidase activity at various time points (Fig. 7B). Though the return to normality with respect to nitric oxide generation in the group pretreated for 3 days was not attained (Fig. 7A), this was adequately compensated for by a concomitant significant increase in reduced glutathione levels (Fig. 8B). The levels of MDA, a biomarker of oxidative lipid peroxidation, were restored to normality with both single-dose pretreatments, while the reduction in the MDA level was not significant in the 3-day KV-pretreated mice (Fig. 8A).
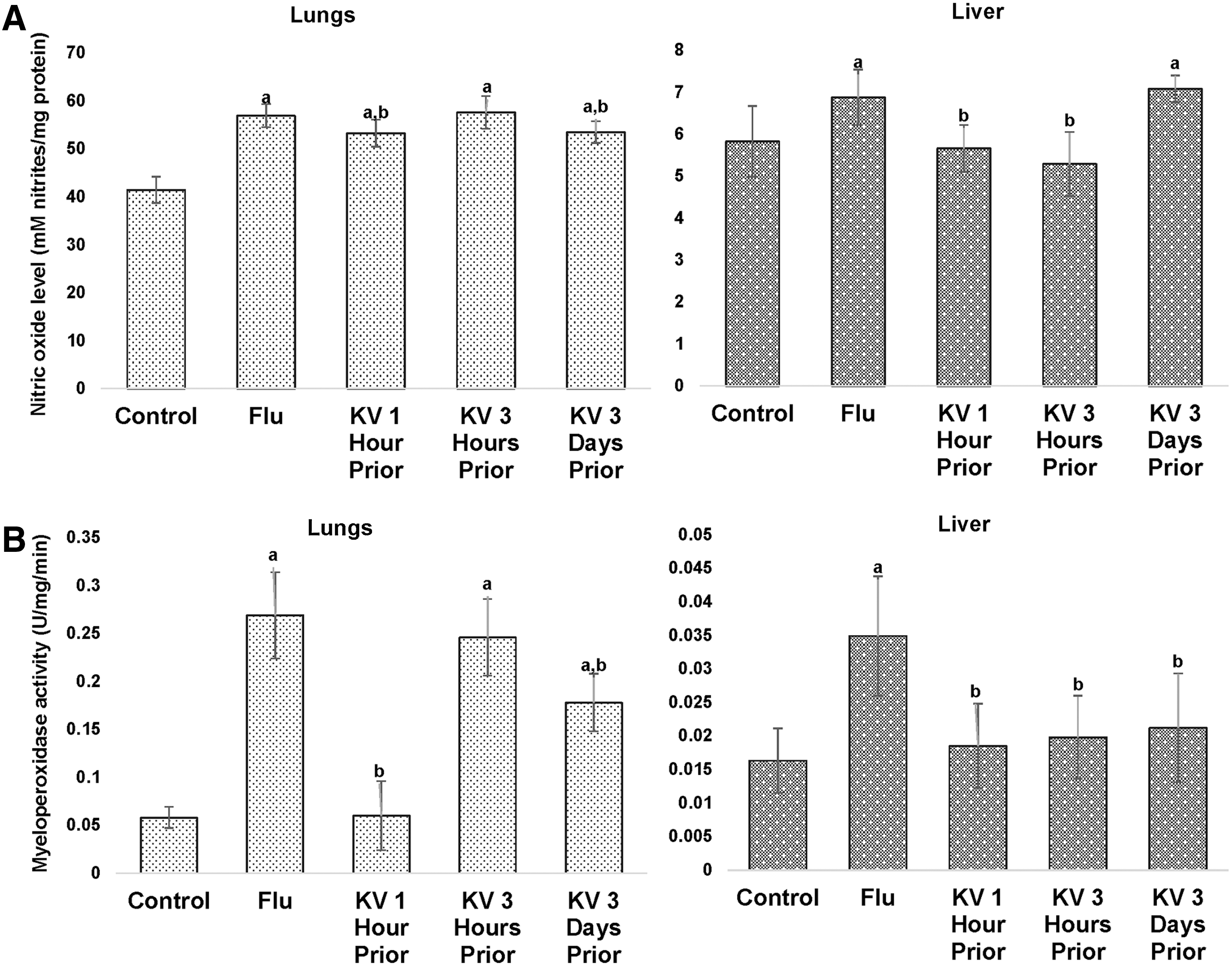
A/H3N2/Perth/16/09 influenza virus induced nitric oxide generation (
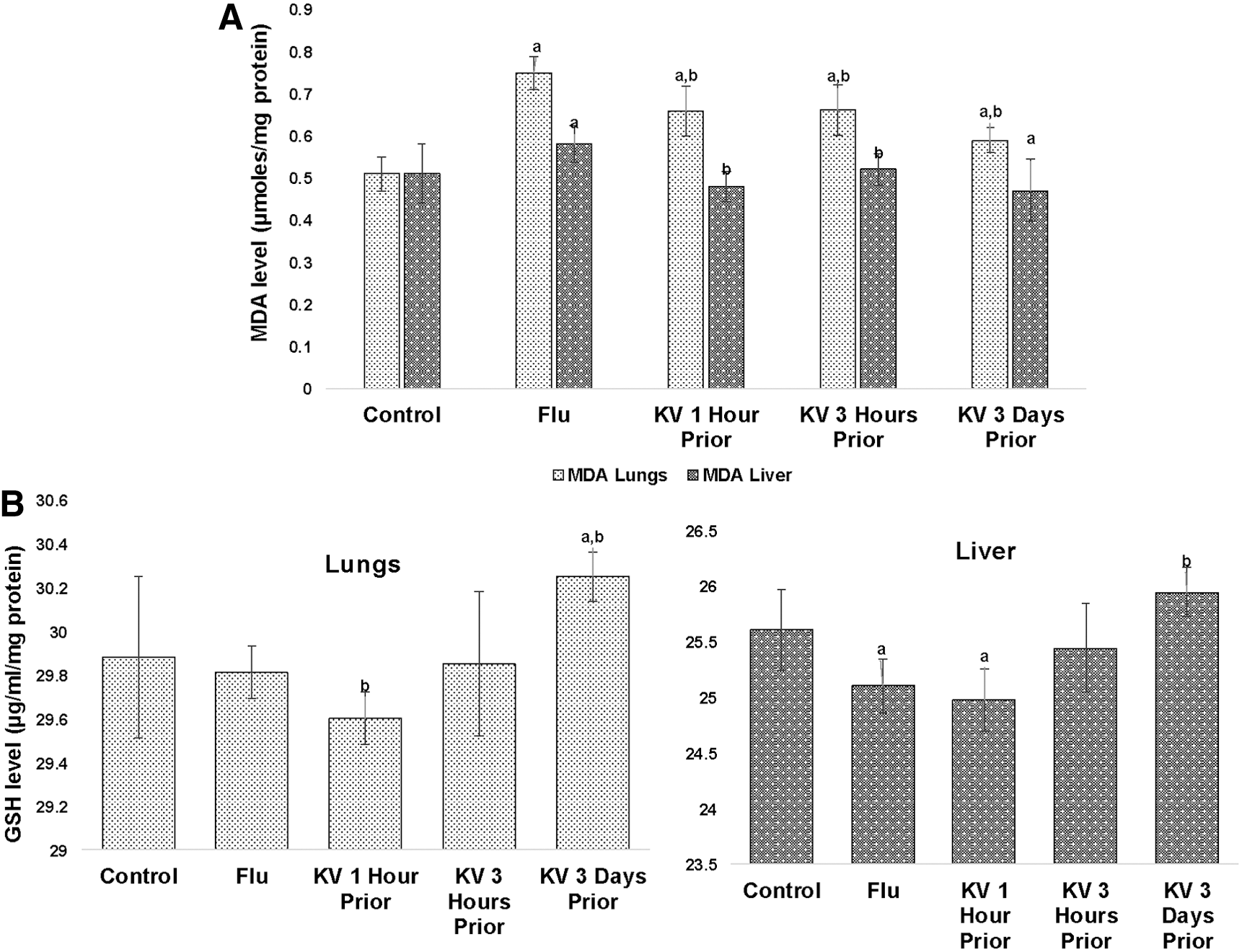
A/H3N2/Perth/16/09 induced malondialdehyde (MDA) formation (
Further investigation revealed that KV may not be able to block the competency and affinity of viral particle for cell surface receptors, a vital step necessary for viral entry into host cells, as it fails to interfere with the virus and/or virus-receptor association (Fig. 9).

In vitro H3N2/Perth/16/09 hemagglutinating preventing capability of KV. (
Discussion
The evolving IAV pandemic strain continues to outsmart the annually predicted vaccines, and seems to defy the drug of choice, neuraminidase inhibitors, especially when used after clinical manifestation of the disease symptoms. A recent paradigm shift includes the development of new anti-influenza compounds with novel antiviral mechanisms (5,25). Influenza virus induces a cytokine storm and the host's intracellular redox imbalance to ensure continuous replication and survival. The implicated oxidative stress in the redo-immunopathology of influenza virus further enhances the multiplication of influenza virus. Thus, worsening the pathology and supplementation with various antioxidants was found to restore intracellular redox status, ameliorate the pathology, and improve the clinical outcome of the infection. Hence, taking charge of the cell's host response by restoring the usurped redox impairment presents a novel anti-influenza target (36,37,47).
This study investigated the protective effect of KV, a biflavonoid complex from Garcinia kola seeds, against mortality and morbidity in BALB/c mice and the biochemical mechanism responsible for protection. Garcinia kola seed is a freely consumed masticatory seed known among other kolas for its peculiar deterring bitter taste, valued and attributed to longevity traditionally in social ceremonies. Many scientific studies have established that the seeds and its extract, KV, possess antioxidant properties, immunomodulatory potentials, and antimicrobial and chemopreventive abilities, even in experimental inflammatory disease models, thus partly validating its ethnomedical claims as antiparasitic and for liver disease (2,3,19,21,22,38).
The present findings indicate that KV at 400 mg/kg extends survival of Pr/H3N2-infected BALB/c mice and improves morbidity, despite the detection of the virus in the lung homogenate of KV-treated mice. Flavonoid and polyphenols are well known for their protein-binding capacities and, as such, envelope proteins of some pathogens, including viruses, are good target of these bioactive phytochemicals (26). It was, however, revealed in an in vitro HI assay that KV weakly interfere with the attachment of infective virus to sialic acid containing receptor. This may suggest that KV's strength as an antiviral may not be via direct contact with the virus or interference with steps involved in homing the virus to the target cell. This type of nonspecific interaction has been reported to be one of the anti-influenza mechanisms of epigallo-catechin-gallate (32). Yoo et al. (2012) also reported a comparable observation with ginseng polysaccharide treatment in mice, enhancing the survival rate with little impact on viral clearance (52). However, in prolonging survival, KV may have downplayed other contributory factors yielding to morbidity and mortality, such as the overzealous host's response dependent pulmonary immunopathology and loss of function as well as multiple organ failure (13,31).
The host's response to influenza virus begins the moment it infects pulmonary epithelial cells and macrophages with phagocytosis and immediate secretion of inflammatory cytokines. While this is gearing toward innate and adaptive immunity via stimulation of more chemokines (type 1 interferons) and inflammatory cells that will orchestrate viral clearance, exacerbated production of pro-inflammatory mediators, including eotaxin, TNFα, IL-1, and IFN-γ, stimulates excessive infiltration and activations of macrophages and neutrophils to inflammatory focus, resulting in increased activities that ultimately culminate in unabated nitric oxide generation and immunopathology (13,39). The present data provide promising evidence that pretreatment with KV prior to viral challenge attenuated nitric oxide production and aggressively suppressed myeloperoxidase activity, a neutrophil enzyme capable of generating nitric oxide and derived nitrating oxidants via inducible nitric oxide (iNOs) expression. Nitric oxide has been identified as a major contributor to influenza virus–induced oxidative damage, mediator of macrophage toxicity contributing to immunopathology and a positive regulator of nuclear factor κB (NfκB) (13,36,37,52). At the molecular level, KV has been identified as an inhibitor of iNOs and NfκB, a transcription factor essential for iNOs expression in both macrophages and neutrophils. NfκB is a cytokine-sensitive transcriptional factor that has also been implicated in unabated IAV replication and many pathological conditions (11,36,37). This corroborates the long-standing notion of tipped inhibitors of iNOs and NfκB as recommended treatment for inflammatory diseases (11).
Influenza virus induces the host's intracellular redox imbalance to ensure continuous replication and survival by depleting the endogenous reduced GSH. GSH preserved intracellular redox homeostasis in a reductive state preventing damaging assaults to biological macromolecules from free radicals and oxidative stress (36,37,43). Treatment with exogenous GSH has been linked to suppression of viral replication through the ability to delay the expression of influenza virus late proteins (7,35). It was observed that KV pretreatment prevailed on the viral-induced oxidative damage to lipids in both the lungs and liver of infected mice. KV primed GSH levels in the pretreated mice, especially when administered for 3 days, both in the liver and the lungs. This may suggest the involvement of multi-organ preservation as a protective mechanism of KV. While the single-dose treatment of KV at different time points were unable to restore the GSH level, they were effective enough to check damage to biomolecules. This could be a function of viral dose, as KV had elicited a robust protection in the earlier studies when mice were treated with 1LD50 and 3LD50.
Collectively, the findings from this preliminary investigation demonstrated that administration of KV significantly prevents morbidity, pulmonary immunopathology, and mortality in BALB/c mice via robust attenuation of unabated activation of inflammatory cells, preservation of redox homeostasis, and multiple organ conservation. This study suggests that KV may be effective for delaying the development of clinical symptoms of influenza virus, and this may be through a mechanism unrelated to those deployed by the existing anti-influenza drugs but closely associated to its antioxidant and immunomodulatory properties.
Footnotes
Acknowledgments
Data analysis and writing of this paper was supported by the Medical Education Partnership Initiative in Nigeria (MEPIN) project funded by Fogarty International Center, the Office of AIDS Research, and the National Human Genome Research Institute of the National Institute of Health, the Health Resources and Services Administration (HRSA), and the Office of the U.S. Global AIDS Coordinator under Award Number R24TW008878. Technology transfer to IOA and technical assistance on fundamental virological techniques by Dr. Clement Meseko of Department of Virology, University of Ibadan, is gratefully appreciated. The content is solely the responsibility of the authors and does not necessarily represent the official views of the funding organizations.
Author Disclosure Statement
No competing financial interests exist.