
Abstract
As a member of superantigens (SAgs) produced by Staphylococcus aureus, staphylococcal enterotoxin B (SEB) is a exotoxin superantigen that can regulate the activity of immunomodulatory and pro-inflammatory cell types. In addition, SEB plays a critical role in the pathogenesis of autoimmune disorders either by initiating the autoimmune process or by inducing a relapse in an individual in clinical remission from an autoimmune disorder. SEB can directly activate T lymphocytes, leading to the release of cytokines, superoxides, or other mediators of inflammation either directly or indirectly, because of its unique ability to cross-link human major histocompatibility complex (MHC) class II and T cell receptors (TCR), forming a trimolecular complex. This review discusses the potential effects of SEB in the pathogenesis of autoimmune diseases such as multiple sclerosis, systemic lupus erythematosus, and rheumatoid arthritis, and explores some updated therapeutic medications to neutralize SEB.
Introduction
A
Compared with normal antigen-induced immune response in which 0.001–0.0001% of the body's T cells are activated, SEB is capable of activating up to 20% of T cells because it has the unique ability to bridge the major histocompatibility complex (MHC) class II on the antigen presenting cells (APCs) and the T cell receptors (TCR) in both CD4 and CD8 T cells (22,37). This bridging effect causes the release of massive amounts of cytokines and chemokines, specifically interleukin 2 (IL-2), tumor necrosis factor α (TNF-α), interferon-γ (IFN-γ), chemokine (C-X-C motif) ligand 1 (CXCL1), and regulated upon activation normal T cell expressed and secreted (RANTES) (16,18,25,27).
Excessive amounts of these cytokines and chemokines result in a dysregulated immune response and cause autoimmune diseases (9,44,48). SEB plays an important role in these autoimmune diseases with unclear mechanisms. The purpose of this article is to summarize the possible mechanisms of SEB in the occurrence and development of several common autoimmune diseases such as multiple sclerosis (MS), systemic lupus erythematosus (SLE), and rheumatoid arthritis (RA), and to explore some updated therapeutic medications to neutralize SEB.
Physico-Chemical Properties of SEB
SEB is an exotoxin secreted by S. aureus, which has high thermal stability and is resistant to most proteolytic enzymes. Even at 78–80°C high temperatures or in acidic environments, SEB still has protein activity and is not completely destroyed within 30 min (23). These characteristics mean SEB retains its biological activities after being ingested through the digestive tract and exerts an exotoxinic effect resulting in local or systemic inflammatory responses. A study by Argudín et al. indicates that SEB can penetrate the intestinal wall, inhibit intestinal absorption of water and electrolytes, and trigger a local or systemic immune response (2).
Superantigen Activity of SEB
Impact of SEB on T cells, B cells, and APCs
Previous research has shown that T cells, B cells, and APCs in patients with autoimmune diseases display altered attributes and play a central role in disease progression and pathology. Pathogenic autoantibodies and cytokines secreted by these immune cells participate in the amplification and perpetuation of inflammatory and autoimmune responses. Excessive activation of immune cells provides the necessary inflammatory milieu for the continued expansion of pathogenic autoreactive clones, breaks immune tolerance, and, thereby, contributes to autoimmune diseases. It is known that SEB can affect immune cells such as T cells, B cells, and APCs, but in the chain reaction of immune cell activation caused by SEB is the key that leads to autoimmune diseases. Several possible mechanisms can be suggested as Figure 1 shows.
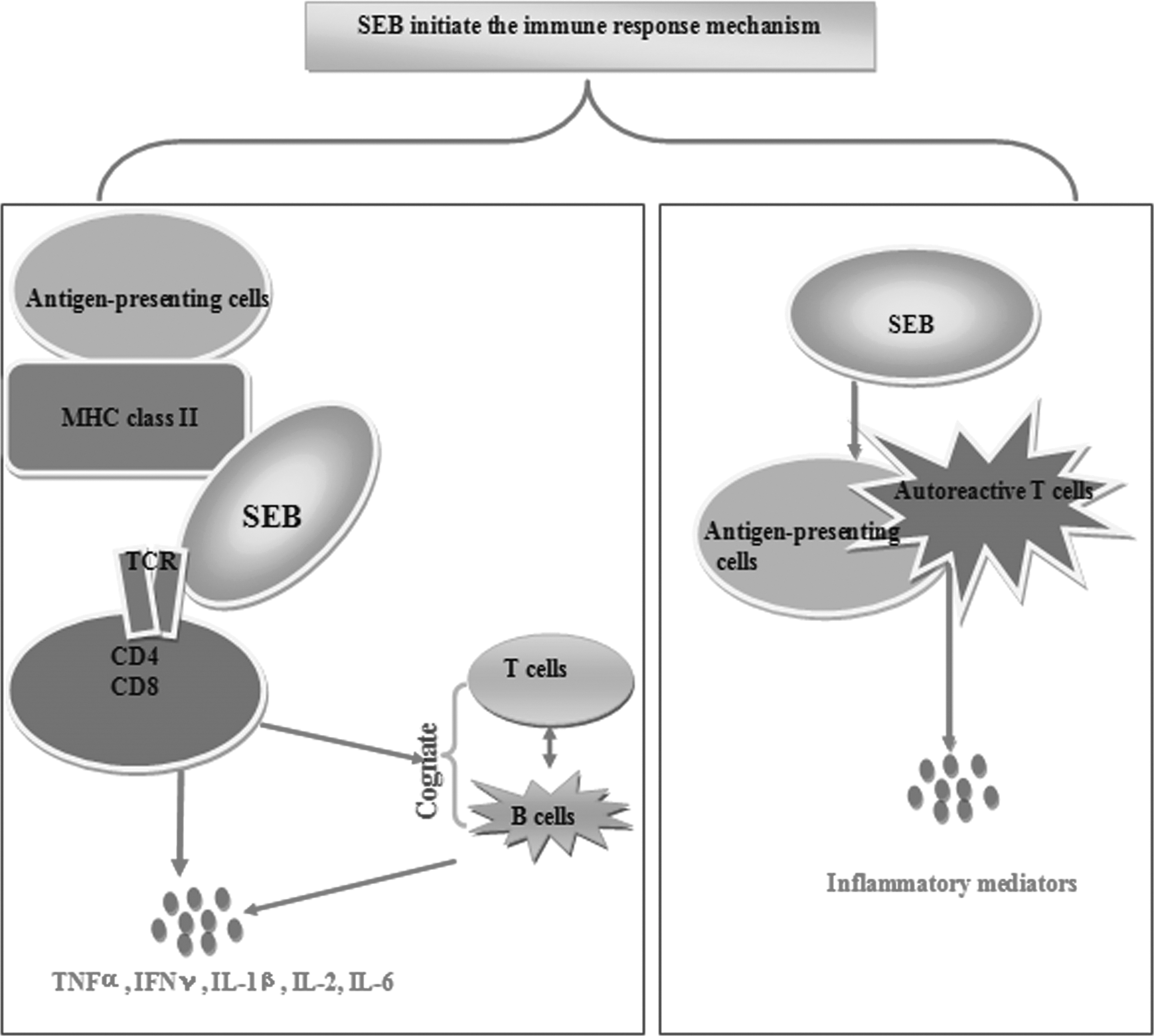
Impact of staphylococcal enterotoxin B (SEB) on T cells, B cells, and antigen presenting cells (APCs).
First, SEB can initiate the immune response by the “bridge” set up between the MHC II on the APCs and the T cell antigen receptor molecules CDR4 area, activating the T cells with the designated Vβ fragment, thereby bypassing the normal antigen processing and presenting mechanism. This bridging effect causes the release of massive amounts of cytokines and chemokines (16,18,25,27). These cytokines and chemokines not only cause the recruitment of additional inflammatory cells but also a relatively deficient activation of negative counter-regulatory feedback loops (39,52,54). More significantly, the mechanism of SEB activating T cells via selective, different Vβ fragments was confirmed to be associated with the migration of TCR spectra in many autoimmune diseases (36,55). Furthermore, T cells need not recognize a specific autoantigen in order to supply help to B cells, they will be activated in a nonclonal fashion by SEB, releasing cytokines in the vicinity of the autoreactive B cells leading to cognate T–B cell interaction and activation of B cells.
It has been suggested that this situation is analogous to a graft-versus-host (GvH) reaction, with SEB serving as a bridge between the T and B cells (15,16). Subsequently, autoantibodies produced by autoreactive B cells may lead to immune complex formation, complement deposition, and eventually result in tissue damage. In addition, SEB may also activate APCs leading to the processing of autoantigens and the presentation of peptides potentially derived from cryptic epitopes, to autoreactive T cells with the subsequent release of multiple inflammatory mediators by both the T cell and the activated macrophage (42).
Impact of SEB on T Helper Cells
The T cell is one of the most important lymphocytes of the immune system, playing a major role in the cellular immune responses, especially the T helper cells (Th). Naive CD4+ T cells differentiate into Th1, Th2, Th17, and T regulatory cells (Tregs) after being activated. Th1 cells mainly produce IFN-γ, TNF-α, and IL-2, and promote cell-mediated immunity. Th2 cells secrete IL-4, IL-5, IL-6, and IL-10. Th17 cells secrete IL-17, IL-21, and IL-22, and induce the release of chemokines. All these cytokines and chemokines are generally involved in humoral immunity (16,18,25,27,30).
Immune balance controlled by Th1 and Th2 cells is critical for the protection of the host from pathogenic invasion, while its imbalance has been demonstrated to play a key role in the induction and development of autoimmune diseases (1,10,53). Previous studies have shown that SEB is the factor that induced both Th1- and Th2-associated cytokines. Plaza et al. confirmed that in vivo administration of SEB induces the acute release of the Th1-type cytokines such as IL-2, IFN-γ, and IL-12 at around 48–72 h (38). A study by Chen et al. also indicated that SEB activated CD4+ T cells to release a high level of Th1-type cytokines rather than Th2-type cytokines (7). Laouini et al. have previously shown that exposure to SEB induces a Th2 cytokine IL-4 expression but not Th1 cytokine IFN-γ expression in the skin of BALB/c mice (21).
Which type of cytokines SEB activates remains controversial. Romani et al. reported SEB treatment led to the early appearance of high levels of IFN-γ, IL-4, and IL-10. However, production of IL-4 and IL-10 returned to normal or slightly increased levels by days 14 to 28, in contrast to the persistence of significant levels of circulating IFN-γ (43). A study by De Heer et al. also validated the murine model for SLE in mice with chronic GvH disease. The first SEB injection increased the release of IL-2, the Th1-type cytokines, but after the second SEB injection, IL-2 was reduced to background values, whereas much more IL-10 and IFN-γ were produced at this stage (13). Studies by Brandt et al. reported that the concentration of SAgs determines the type of immune response elicited, namely, Th1 versus Th2. Low concentrations of SAgs polarize to a Th2-type response and high concentrations to a Th1-type response (4). It seems there is a dose-related effect of SEB adjusting the immune balance between Th cells.
Impact of SEB on Tregs
In both mice and human subjects, CD4+CD25+ Tregs constitute 5–15% of peripheral CD4+ T cells. Accumulating evidence indicates that CD4+CD25+ Tregs are essential for the maintenance of self-tolerance (28). Tregs can inhibit activation and proliferation of CD4+ or CD8+ T cells. They can also inhibit the initial T cell proliferation reaction through direct contact with target cells. The inability of human Tregs to suppress strongly activated CD4+CD25– Tregs in inflammatory environments has major implications in autoimmune diseases (11). A study by Florquin et al. led to the postulation that repeated injection of low doses of SEB induces the development of CD4+ Tregs capable of suppressing the primary response of SEB-specific T cells (14). This hypothesis was also supported by the observation that, in BALB/c mice made unresponsive to SEB, 10–15% of SEB-specific TCR Vβ8 bearing CD4+ T cells overexpressed CD152 (CTLA-4), a molecule described recently as preferentially expressed by Tregs (33,49). Cardona et al. also indicated SEB can act as an environmental factor that effectively inhibits the activity of Tregs (6).
Impact of SEB on Epithelial Cells and Acidophilic Granulocytes
Recent studies indicated that epithelial cells have a wide range of biological activity. They can express HLA-DR and CD86 and can therefore be used as APCs for antigen processing. In addition, epithelial cells can release multiple cytokines (IL-4, IL-6, IL-8, and TNF-α), chemokines (eotaxin, RANTES), and adhesion molecules (ICAM-1 and VCAM-1). Damm et al. verified that SEB can stimulate the nasal mucosa epithelial cells to express IL-5 and granulocyte-macrophage colony stimulating factor (GM-CSF) significantly. These two types of molecules are known as strong chemokines and antiapoptotic factors of acidophilic granulocytes (12,56). Krakaur et al. also discovered that SEB can induce high levels of CC chemokines such as monocyte chemotactic protein-1 (MCP-1) and macrophage inflammatory protein-1 (MIP-1) in order to promote aggregation and activation of acidophilic granulocytes (20).
SEB Affects Autoimmune Diseases
SEB in MS
MS is a chronic disease of the central nervous system, and is widely accepted as a complex autoimmune disease that involves both host genetic and environmental factors. Recently, there has been increased evidence to support the theory that S. aureus may carry toxins that can act as SAgs that activate large numbers of CD4+ cells and have been implicated in various autoimmune diseases, including MS, Wegener's granulomatosis, and RA (24,50). While the role of SEB in the etiology of MS is currently unknown, it has been demonstrated that it is involved in the reactivation of a mouse model for MS known as experimental autoimmune encephalomyelitis (EAE). Brocke (5) and Schiffenbauer et al. (45) determined that SEB exacerbated the course of EAE because it could activate Vβ8+ T cells, which have been shown to be important in the initiation of EAE in PL/J mice. However, Racke et al. found SEB treatment of (PLxSJL) F1 mice energized MBP-specific T cells using Vβ8, and subsequent in vitro activation with SEB resulted in a marked decrease in proliferation to SEB and inability to transfer EAE (41). Therefore, they concluded SEB could protect PL/J mice from the development of EAE. Consistent with this observation, Soos et al. also reported that the treatment of PL/J mice with SEB was able to protect PL/J mice from the development of EAE (46). The two conflicting results are worthy of further study.
SEB in SLE
SLE is a chronic autoimmune disease that affects multiple organ systems and is characterized by polyclonal B cell activation and high levels of autoantibody production. Multiple inherited risk factors, combined with random environmental factors, are thought to be responsible for the abnormal immune response. SLE can be triggered by a number of factors, including hormonal imbalance at puberty, environmental factors, certain medication, and viral infections. Tumang et al. found chronic SEB administration to nonautoimmune-prone mice resulted in polyclonal hypergammaglobulinemia and increased titers of autoantibodies (51). This led to the hypothesis that SEB may also influence the course of SLE. A study by Chowdhary et al. supported this hypothesis. They found that chronic exposure to small amounts of SEB resulted in a multisystem autoimmune inflammatory disease in a HLA-DQ8 transgenic mouse model, with features similar to human SLE. Chronic exposure to SEB causes significantly elevated levels of anti-dsDNA and anti-Sm antibodies, T lymphocyte infiltration in the liver and kidney, significant expansion of total CD4+Foxp3+T cells, as well as TCR Vβ8+Foxp3+ Tregs in the spleen of the donor mice. They concludeded chronic exposure to extremely small amounts of SEB could be an etiological factor for SLE and can result in a CD4+ T cell–dependent, IFN-dependent, Th1-mediated multisystem immunologic derangement (8).
In contrast, Kim et al. found a preventative effect of SEB administration on the induction of SLE. They reported that SEB reduced lupus nephritis of MRL/lpr mice. Rather than causing the expansion of Vβ8+ T cells, SEB administration resulted in the reduction of Vβ8+, CD4–CDS– “double- negative” (DN) T cells (19). It seems that the role and mechanism of SEB in the pathogenesis of SLE remains controversial.
SEB in RA
RA is an autoimmune disease characterized by marked inflammation and destruction of several joints. SEB has been suggested to have a critical role in the pathogenesis of RA by activating T and B cells in the combination with Vβ-TCR and activating polyclonal B cells as a bridge between T and B cells at the same time. Histological studies of synovium have shown that RA patients had increased levels of serum IgM SEB antibody compared with normal subjects, and synovial cells could serve as APCs for SEB and induce T cell activation (35,48). Studies of collagen-induced arthritis (CIA), the experimental disease model of human RA, also confirmed the critical role of SEB in activating specific autoreactive Vβ8+ T cell clones produce Th1 cytokines, IL-2, and IFN-γ, which have been implicated as important mediators of inflammation and joint destruction in RA (34). Thus, based on the above studies in mice and in vitro, it appears that SEB can influence the course of RA and may have a significant influence on the ability of synovial cells to activate T cells.
Possible Mechanism of SEB in Autoimmune Diseases
Although the studies mentioned above are comprehensive, it is not difficult to find evidence of SEB as either initiating or preventing the development of autoimmune diseases. Previous studies of SEB's performance in immune cells, especially Th and Tregs, also demonstrate the dual immunomodulatory activity of SEB acted in the immune system (4,6,13,14,21,33,43,49). Based on the above research, it is speculated that SEB acts as a bidirectional immunoregulatory factor in autoimmune diseases, as shown in Figure 2.
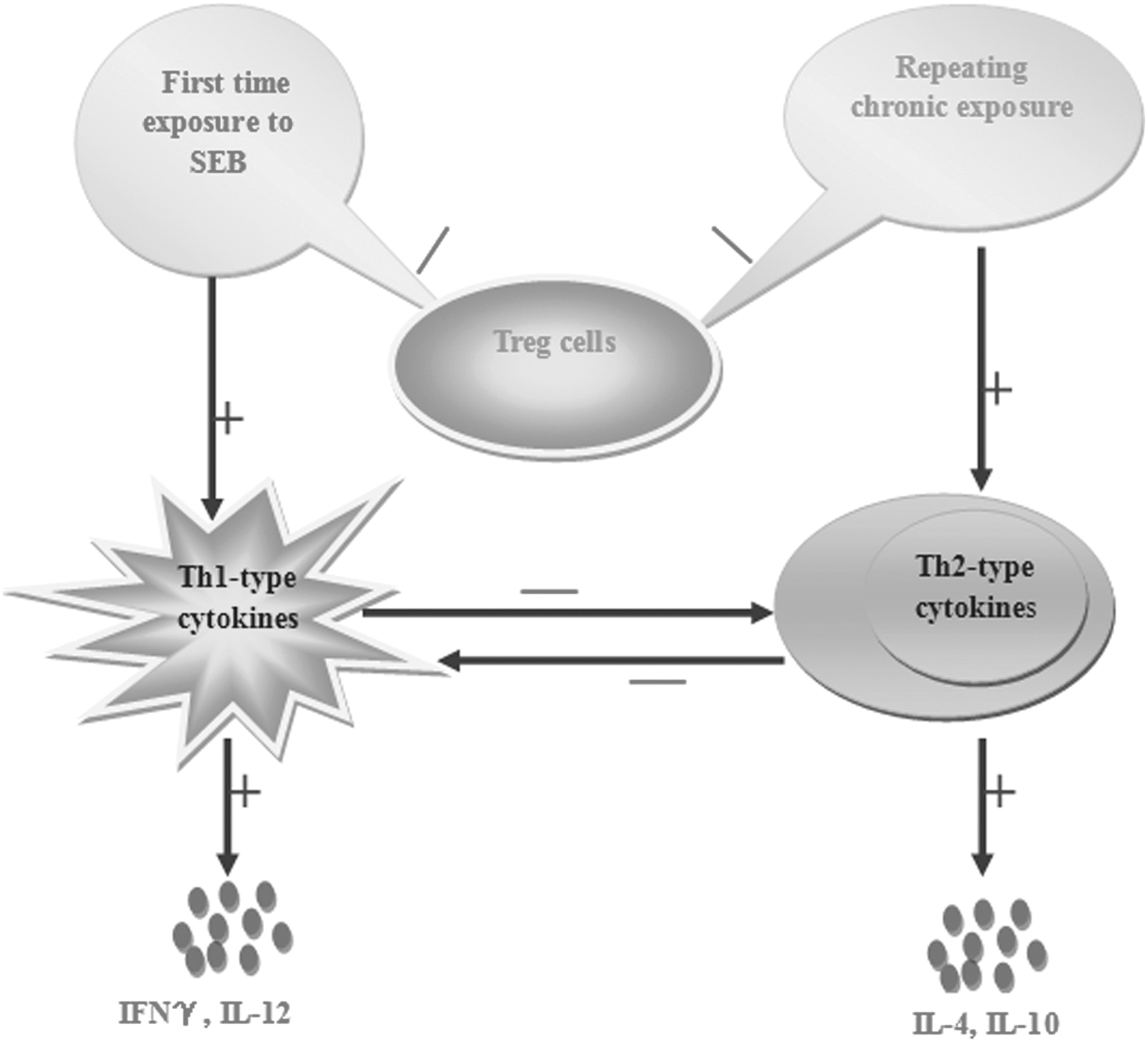
Possible mechanism of SEB in autoimmune diseases.
When first exposed to SEB, the immune response of Th cells is activated during the early course of the autoimmune disease, while Treg cells lose their immunosuppressive activity. This process leads to the initiation of immune responses and the proliferation of Th1-type cytokines such as IFN-γ and IL-12, which inhibit the production of Th2-type cytokines to eliminate antigens. That is why high levels of Th1-type cytokines can be seen in the early stage of autoimmune diseases. Then, after a period of time, when the concentration of SEB drops to a lower concentration or SEB is exhausted, Th2-type cytokines occupy the dominant position of Th1-type cytokines instead, and Treg cells exert their immunosuppressive activity. On the other hand, when repeating chronic exposure to low doses of SEB or Th2-type immune response in the body already exists before exposure to SEB, an inflammatory response will be intensified and a proliferation of Th2-type cytokines will be secreted and play a dominant role in autoimmune diseases. Dynamic changes of SEB cause a repeated and prolonged course of autoimmune diseases.
Strategies to Neutralize SEB
SEB can exist in water and contaminated food or as an aerosol, the inhalation of which may cause an extreme stress reaction of the immune system and endotoxin shock (31). Currently, the conventional therapy for SEB poisoning is intravenous immunoglobulin. Considering that immunoglobulin from healthy volunteers has a complex antibody component and the curative effect is not stable, some research institutes have tried to develop antibodies that directly target SEB (17). In addition, the mutation vaccine of SEB, which has just proceeded to Phase I clinical trials, is also considered a good treatment strategy (32). However, since the immunoprotective effect of the vaccine dictates that vaccination must take place more than 2 weeks in advance, it is only used for the prevention of SEB, not for the therapeutic effect of SEB-induced toxic reaction that has already appeared.
Studies by Maródi (26) and Mitra et al. (29) confirmed that signal transducer and activator of transcription (STAT) proteins such as STAT1, STAT3, and STAT5B are key components in host defense against various viruses, bacteria, and fungi. Mutations of these STATs may disrupt various cytokine pathways that control mucocutaneous immunity, particularly against S. aureus (26,29). The study by Qiu et al. found that many of the traditional Chinese medicine monomers and volatile oils such as thymol, eugenol, liquorice A chalcone, basil essential oil, and peppermint essential oil can reduce the expression of SEB in a dose-dependent manner (40). Although these methods have played a role in the treatment of SEB, complex ingredient and pharmacodynamic evaluation is required in clinical trials, which has limited their application. Development of fast and efficient treatment for SEB is the focus of the authors' future work.
Conclusion
Evaluating the important roles of SEB in regulating immunocytes has begun to shed light on the mechanism of bacterial infections in autoimmune diseases. SEB has been reported to be involved in autoimmunity diseases such as MS, RA, and SLE in humans and laboratory animals by stimulating activation, expansion, anergy, or deletion of T cells expressing the appropriate Vβ segment of the TCR and releasing different Th-type cytokines under different circumstances. However, other studies have indicated that SEB plays a protective role in the disease process of autoimmune diseases. These studies suggest that SEB may act as a bidirectional regulating factor in autoimmune diseases in a staged and dose-related manner, and that the mechanism requires further research.
Footnotes
Acknowledgments
This work was financed by the Doctoral Scientific Research Foundation of Anhui University of Chinese Medicine (2013RC002).
Author Disclosure Statement
No competing financial interests exist.