
Abstract
Antibody responses to vaccines can be influenced by various behavioral and psychosocial factors. Few reports exist on the impact of fighting on antibody response to vaccines. This study unexpectedly found that fighting could significantly enhance antibody production in male mice immunized with hepatitis B virus (HBV) vaccines. To confirm the finding, a mouse-fighting model was established in which it was observed that only intense fighting, not mild fighting, enhanced the antibody response to HBV surface antigen in male mice, and that the frequency of fighting and active attacks during fighting showed no obvious relationship with the antibody levels in the male mice that experienced fighting. In addition, fighting can cause significant upregulation of CD80 in CD11c+ cells in the spleen of male mice. These data suggest that fighting could influence the humoral immune response in individuals immunized with vaccines or infected with microbes.
Introduction
C
Accumulated evidence has revealed that psychosocial factors could impact on antibody response to vaccines bidirectionally (8). A meta-analysis of 13 studies involving 1,158 men and women revealed a significant negative association between psychological stress and antibody responses to influenza vaccination (24). In contrast, a study of 60 young healthy adults (31 men) showed that mental stress prior to vaccination enhanced antibody responses to influenza vaccines in women but not in men (5). Similarly, mental stress before meningococcal A + C vaccination was observed to enhance the antibody response to vaccines in healthy young men (6). A further study revealed that the stress-enhanced antibody response to both influenza vaccine and meningococcal A + C polysaccharide vaccine may be associated with heightened cardiovascular reactivity to stress (20). The association could be supported by the observations that acute exercise prior to influenza vaccination (5) or meningococcal A + C vaccination (6) in young healthy adults enhanced their antibody responses to the vaccines. The exercise-enhanced antibody response to the vaccines could be attributed to the hypothalamic–pituitary–adrenal axis and sympathetic nervous system, which are activated during exercise (2). Based upon the above findings, it might be postulated that fighting, an intense “exercise,” could influence the antibody response to vaccines, especially in males who are usually more aggressive than females are.
The initial purpose of the present study was to observe whether water in oil emulsion adjuvant could influence sexually dimorphic antibody responses to recombinant hepatitis B virus surface antigen (HBsAg) in mice. Unexpectedly, an obviously elevated antibody response was found to HBsAg vaccines in a group of male BALB/c mice that experienced severe fighting. To confirm the finding, an attempt was made to establish a mouse fighting model, and then the potential associations between fighting and the enhanced antibody response to HBsAg vaccines in male mice were examined.
Materials and Methods
Reagents and HBV vaccine formulation
Recombinant HBsAg was kindly provided by Walvax Biotechnology Co. Ltd. (China). ISA206 was purchased from Seppic Company (France). Horseradish peroxidase (HRP)-conjugated goat antimouse IgG was from Jackson ImmunoResearch Laboratories. HBV vaccine was prepared by mixing HBsAg (20 μg/mL) with ISA 206 adjuvant (water-in-oil-in-water emulsifier) at a 1:1 (v/v) ratio and stored at 4°C.
Mice
Specific-pathogen-free female and male (SPF) F1 (BALB/c ♀ × C57BL/6 ♂) mice and BALB/c mice, aged 6 weeks, were purchased from the Experimental Animal Center, Jilin University (Changchun, China). Female and male C57BL/6 mice, aged 3 weeks, were purchased from Beijing Weitonglihua Laboratory Animal Co. Ltd. All mice were maintained at 22 ± 2°C with a 12 h light/dark cycle and had free access to food and water in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals.
Animal experiments
Regular immunization procedure
Mice were immunized intramuscularly with HBsAg or HBV vaccine with 1 μg HBsAg per mouse on day 0 and day 14, and bled on day 21 and 28 for collecting sera. Each group was composed of six mice, either female or male. Behaviors of the mice were monitored and recorded on a daily basis.
Induction of fighting among immunized male BALB/c mice
Six male BALB/c mice aged 6 weeks were individually kept in different self-sustaining cages for 7 days, and then put together in one cage to arouse fighting. After meeting for several minutes, the mice began to fight each other. The fighting lasted for approximately1 h, after which the mice were immunized with HBsAg or HBV vaccine. To arouse more intense fighting, one male C57BL/6 mouse was introduced into the cage where the six immunized BALB/c male mice were kept. The introduction aroused fighting immediately between the male C57BL/6 mouse and each of the male BALB/c mice. The fighting was intermittent with an interval of about 2–5 min. The intermittent fighting lasted for around 12 h in some instances. On day 14 after the first immunization, the fighting male BALB/c mice were boosted with HBsAg and HBV vaccine, respectively. In parallel, the nonfighting BALB/c mice were immunized with HBsAg and HBV vaccines, respectively, on the same schedule. All the mice were bled on day 21 and day 28 in order to collect sera, and their behaviors were recorded during the whole experiment.
Establishment of a fighting model using male C57BL/6 mice
The male C57BL/6 mice were aroused to fight with the following strategy (Fig. 3A). (a) The 10 C57BL/6 male mice, aged 3 weeks, were fed in a self-sustained cage for a month. (b) The 10 mice were randomly divided into two groups (group A and group B) and the mice were labeled A1–A5 in group A and B1–B5 in group B, respectively. (c) A1 and B1, A2 and B2, A3 and B3, A4 and B4, and A5 and B5 were put into five individual cages and were kept for 1 h when the two mice fought severely. Then, the paired mice were separated into single self-sustained cages where they were kept for 23 h. So, in 24 h, the mice experienced a fighting-inducing round consisting of pairing, fighting, and resting. (d) In the second fighting-inducing round, the mice were coupled A1 to B2, A2 to B3, A3 to B4, A4 to B5, and A5 to B1. (e) In the third fighting-inducing round, the mice were coupled A1 to B3, A2 to B4, and so forth. (f) In the first cycle, the mice were induced for fighting for five rounds. (g) After the cycle, the mice were kept individually for 9 days before being induced to fight for another cycle. The mouse fighting was recorded with video by which the fighting number for each mouse was calculated. To measure the fighting more accurately, an evaluating standard of fighting number was defined as one time when two mice chased and bit each other for more than 3 sec. If the average fighting number was more than 10 times, the fighting model was regarded as established.
Immunization of fighting model mice
Ten male C57BL/6 mice, aged 3 weeks old, were kept in a self-sustaining cage for 30 days. Afterwards, the mice were induced to fight for three rounds (as described above) and were immunized with HBsAg and HBV vaccine, respectively. On day 14 after the first immunization, the model mice were boosted. On day 21 and day 28, the mice were bled to collect sera to detect anti-HBsAg antibody (HBsAb).
HBsAb detection by indirect enzyme-linked immunosorbent assay
The HBsAb in the sera of the immunized mice was detected by indirect enzyme-linked immunosorbent assay (ELISA) (11).
Cell isolation and culture
Splenocytes (1 × 106 cells/mL) or peripheral blood mononuclear cells (PBMCs) isolated from fighting and nonfighting C57BL/6 mice were directly stained with fluorescent antibodies. Alternatively, splenocytes (1 × 106 cells/mL) in RPMI-1640 (Invitrogen Life Technologies, Carlsbad, CA), supplemented with 10% fetal calf serum (FCS; Hyclone, Logan, UT), penicillin (100 IU/mL), and streptomycin (100 μg/mL) were cultured with HBsAg (1 μg/mL) for 24 h.
Flow cytometry
To assess the activation of dendritic cells and B cells, the freshly isolated or in vitro stimulated PBMCs and splenocytes were stained with fluorescein isothiocyanate (FITC)-labeled anti CD11c and PE-labeled anti-CD 80 mAb or APC-labeled anti-80 mAb for 30 min. After washing, the stained cells were detected on a flow cytometer.
Statistical analysis
The data were analyzed using t-tests. The correlation between the average fighting number and antibody response was assessed by linear correlation analysis. Differences were considered statistically significant at p < 0.05.
Results
Phenomenon of fighting-enhanced antibody response in male mice to HBV vaccine
To test whether ISA206 adjuvant could influence sexually dimorphic antibody response to vaccines, female and male F1 (BALB/c ♀ × C57BL/6 ♂) mice were immunized with HBsAg or HBV vaccine (HBsAg plus ISA206 adjuvant) twice on day 0 and day 14, and bled on day 21 (Fig. 1A) to collect sera. The anti-HBsAg antibodies (HBsAb) in the sera were detected by ELISA. The results showed that the female F1 mice produced significantly higher levels of HBsAb to HBsAg (p = 0.0085) or HBV vaccine (p = 0.0474) than the male F1 mice did. ISA206 adjuvant enhanced immunogenicity of HBsAg in both the female and the male mice (p = 0.0198 and p = 0.0011, respectively; Fig. 1B), and could not reverse the sexually dimorphic antibody responses to HBsAg. To confirm the observation, female and male BALB/c mice were immunized with the HBsAg or HBV vaccine and were bled to detect HBsAb following the above-mentioned procedure. As shown in Figure 1C, HBsAg without ISA206 adjuvant induced a more vigorous HBsAb response in the female BALB/c mice compared with the males (p = 0.0339). HBsAg with ISA206 adjuvant induced similar level of HBsAb in the female and the male BALB/c mice. Unexpectedly, significantly higher levels of HBsAb antibodies to HBsAg (p = 0.0374), and especially to HBV vaccine (p < 0.0001), were detected in the sera from the groups of male mice. Unlike F1 male mice, the BALB/c mice were noticed to have experienced fighting because that their tails and ears were severely bitten. Therefore, the data suggested that fighting could somehow enhance the HBsAb antibody response to HBsAg and HBV vaccine in male BALB/c mice.
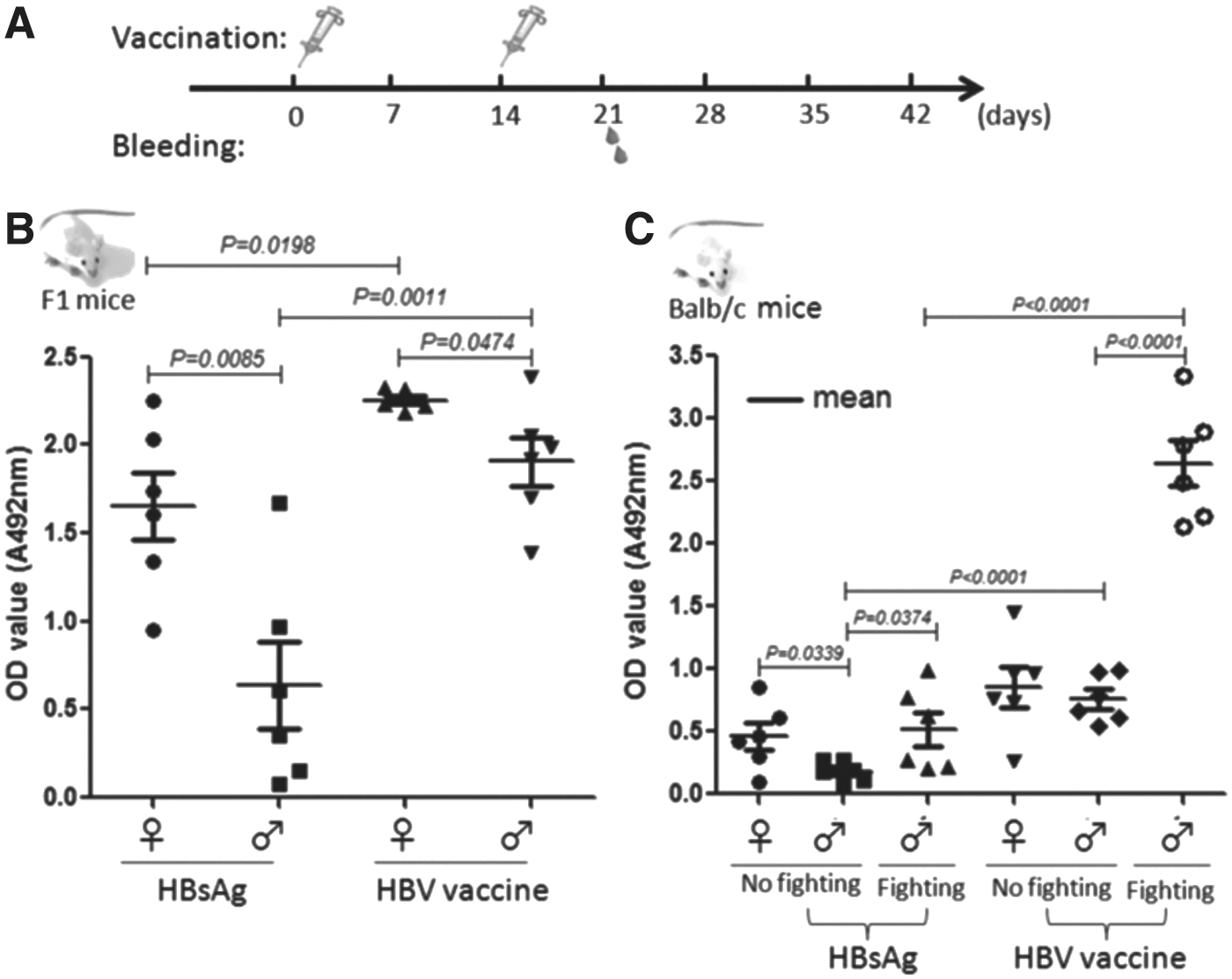
Anti-HBsAg antibody (HBsAb) levels in the sera of hepatitis B virus (HBV) vaccine-immunized F1 and BALB/c mice. (
Effect of mild fighting on antibody response in male BALB/c mice to vaccine
To verify the impact of fighting on antibody production, six male BALB/c mice were individually kept for 7 days, and then put together in one cage to arouse fighting (Fig. 2A), which was lasted for about 1 h. The mice were subsequently immunized with HBsAg and HBV vaccine, respectively. After the first immunization, a male C57BL/6 mouse was introduced to the six immunized BALB/c male mice and kept in the same cage to induce fighting. On day 14 after the first immunization, the fighting-induced male BALB/c mice were boosted. In parallel, nonfighting BALB/c mice were set up as controls. All the mice were bled on day 21 and day 28 after the first immunization to detect the HBsAb antibody response. The result showed that female BALB/c mice also produced significantly higher levels of HBsAb antibody than male BALB/c mice did on both day 21 and day 28 (p = 0.0214 and p = 0.0314, respectively), and that fighting tended to enhance the HBsAb response in male BALB/c mice (Fig. 2B). Noticeably, it was observed that the male C57BL/6 mouse intruder did not induce lasted intense fighting between the male C57BL/6 mouse and each of the BALB/c male mice, and that the fighting happened fewer than 10 times within 1 h during the experiment. Together, the results imply that the intensity of fighting is mild and not strong enough to impact the antibody response to HBV vaccine.
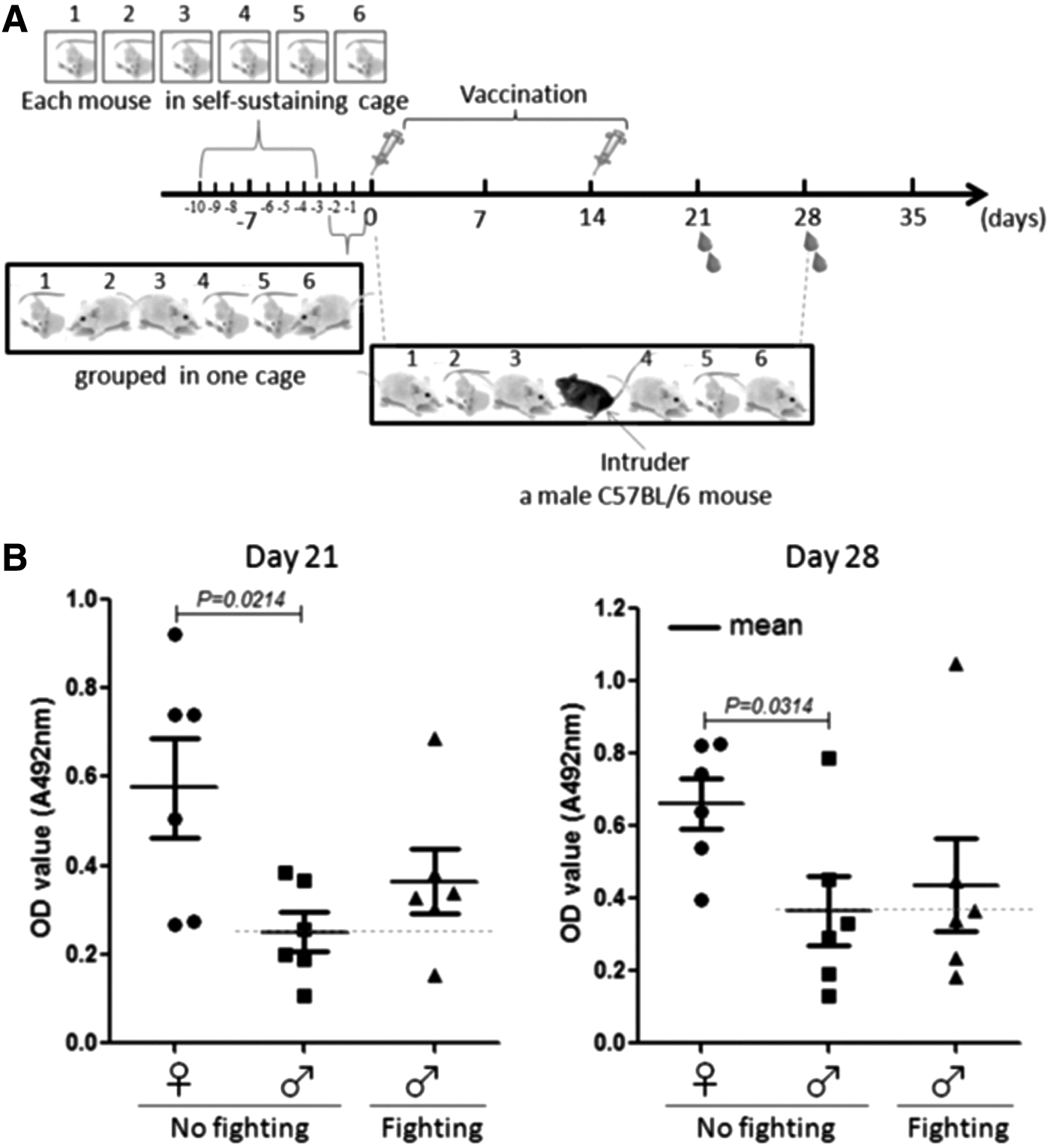
HBsAb levels in the sera of female and fighting and nonfighting male BALB/c mice. The BALB/c mice were artificially induced to fight and were immunized with HBV vaccines based on the procedures described in Materials and Methods. (
Effect of intense fighting on antibody responses in male C57BL/6 mice to HBV vaccine
To induce more intense fighting, C57BL/6 mice, more aggressive than BALB/c mice (7), were used to establish a fighting model (Fig. 3A) as described in Materials and Methods. To assess the fighting more accurately, the fighting number and the active attack in which the mice were initiatively provoked to fight were recorded. In the model, all of the paired mice fought each other more than 10 times in an hour, which was considered as intense fighting. The fighting-induced male C57BL/6 mice were immunized with HBV vaccines twice. In parallel, both of the male C57BL/6 mice that did not fight and female C57BL/6 mice that did not fight were used as controls. On day 21 and day 28 after the first immunization, the mice were tested for HBsAb production. As shown in Figure 3B, the HBsAb levels in the fighting-induced male C57BL/6 mice were significantly elevated on day 21 (p = 0.0218), not on day 28, compared with nonfighting male C57BL/6 mice. However, no correlation between the fighting number and the level of HBV-specific antibody were found, although the active attack times trended to be reversely related to the antibody level in A5 and B3 mice (Fig. 3C). The lack of a correlation was confirmed by further linear correlation analysis (Fig. 3D). These results indicate that fighting elevates antibody levels transiently in male C57BL/6 mice, and does not show any relationship between fighting number and active attack number.
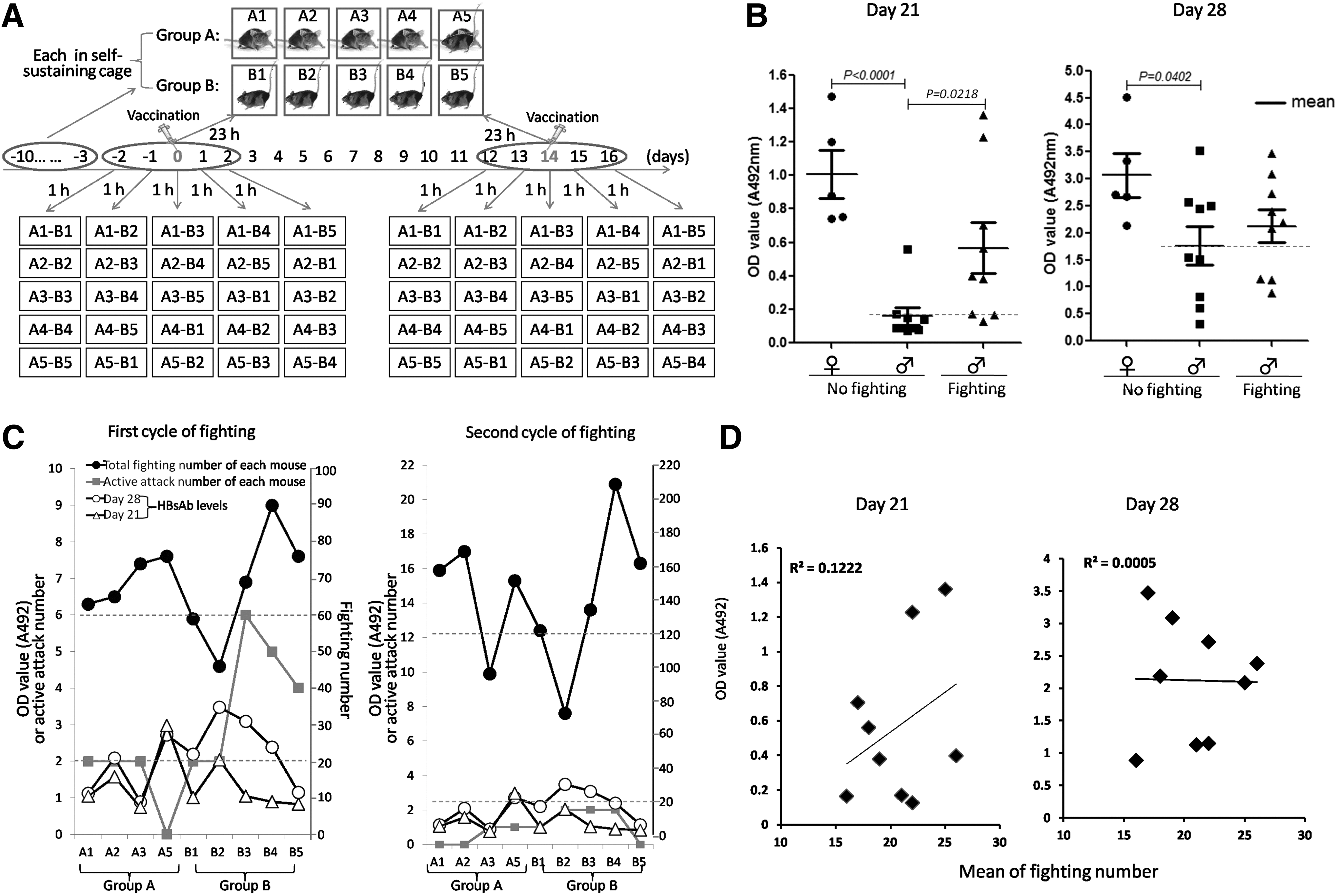
Establishment of the fighting model and analysis of the relationship between fighting and antibody production in male C57BL/6 mice. (
Mobilization of immune cells from spleen in mice experienced fighting
To find how fighting could influence the antibody response, male mice were separately fed in self-sustained cages for a week, followed by inducing fighting once a day for three days. The nonfighting male and female mice were used as controls. On day 3, all mice were sacrificed to collect blood and the spleen (Fig. 4A). By using flow cytometry, CD80 and CD86 expression of dendritic cells (CD11c+) in the spleens and PBMCs were detected. The results demonstrated that the ratios of CD80+CD11c+ and CD86+CD11C+ cells in PBMCs tended to be higher in the male mice experiencing the fighting than that in male mice without fighting, and that the ratios of CD80+CD11c+ and CD86+CD11C+ cells in the spleens were similar among the mice in the three groups (Fig. 4B). Furthermore, the splenocytes collected from the mice in the three groups were cultured with HBsAg for 24 h and 48 h, respectively, followed by staining with florescent-labeled antibodies against CD80, CD11c, and CD86. The flow cytometry analysis showed that the ratios of CD80+CD11c+ cells were significantly increased in the spleens of the male mice experienced fighting compared with that in the spleens of the nonfighting male mice (p = 0.0290 for 24 h cultured cells and p = 0.0144 for 48 h cultured cells, respectively), and not significantly different from that in female mice (Fig. 4C, left). The ratios of CD86+CD11+ cells were also higher in fighting male mice than that in female mice and nonfighting male mice (p = 0.0663 and p = 0.0547, respectively; Fig. 4C, right). These results indicate that fighting does indeed affect the activation of dendritic cells, which may be reason for various antibody responses.
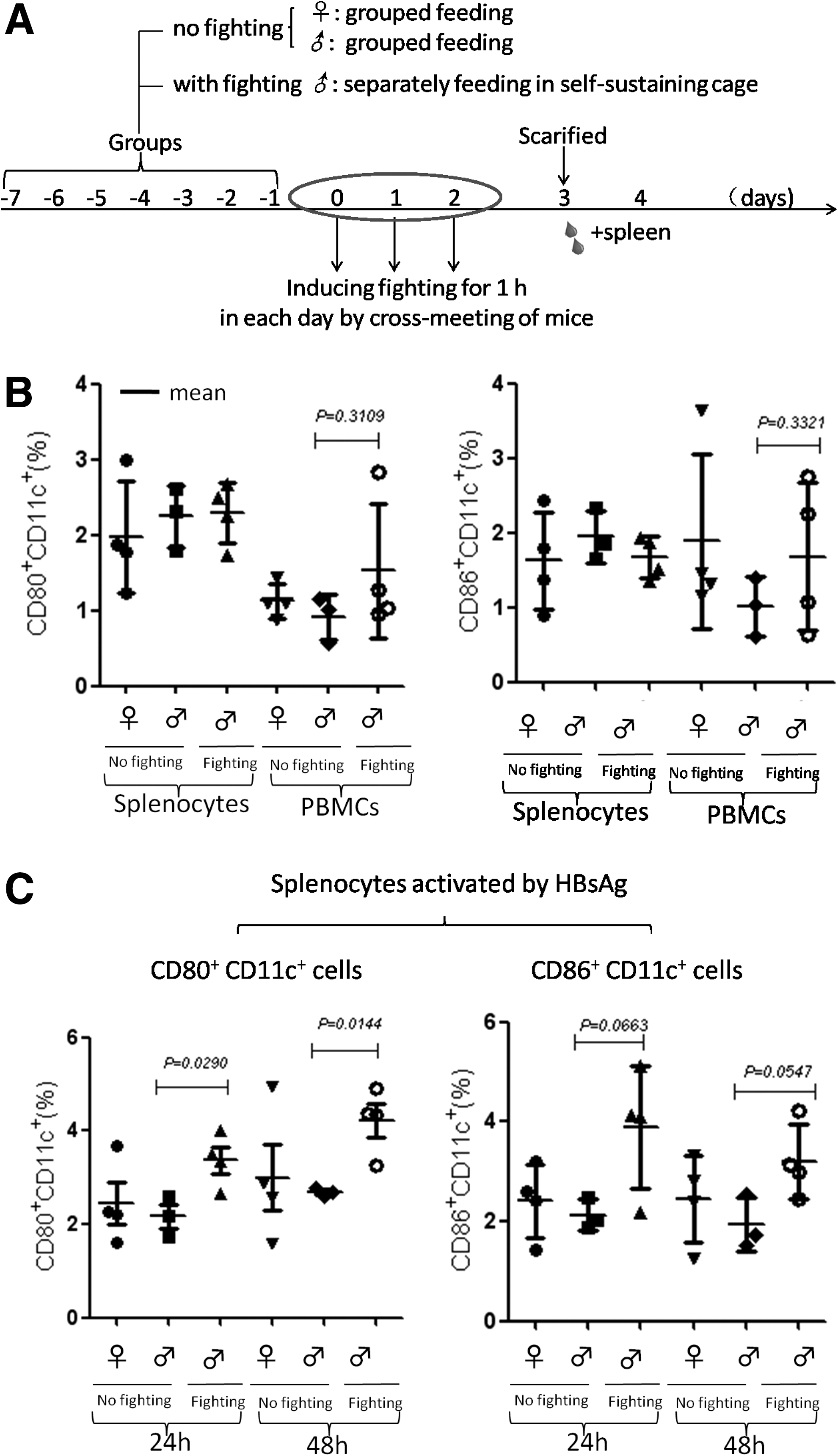
Impact of fighting on the activation of immune cells in mice. (
Discussion
Typically, females have a higher immune response to infections and vaccinations than males do. This study also found that female mice produced significantly higher levels of anti-HBsAg antibody to HBV vaccine (HBsAg plus ISA206 adjuvant) than male mice did. Unexpectedly, it was observed that male BALB/c mice that experienced spontaneous intense fighting produced HBsAb at a similar level as female BALB/c mice. To confirm the observation, male mice were induced to fight and then their HBsAb response to HBV vaccines was tested. Initially, male BALB/c mice were selected for fighting induction by separating them in self-sustained cages for a while and then mixing them together. However, the fighting was too mild. Possibly, the less intense fighting was insufficient to promote the antibody response to vaccines in the male mice. To induce more intense fighting, C57BL/6 mice—more aggressive than BALB/c mice—were used to establish the fighting model (14). Three-week-old male C57BL/6 mice were fed in self-sustained cages for a month before arousing them to fight in a prearranged scheme as shown in Figure 3A. In the schemed conditions, the male C57BL/6 mice experienced 10 h intense fighting in 10 days, and the fighting was quantitatively evaluated with average fighting number. By using the fighting model, it was confirmed that fighting did promote an antibody response to vaccine in male mice. The finding is different from previous reports that showed that intense exercise, which surely occurred in fighting, led to immune suppression not promotion (18). The incompatibility may arise from the difference between the intense exercise and fighting. The fighting individuals not only experienced intense exercise, but also psychological stress that may impact the immune responses of the male mice. From the point of view of evolution, psychological stress, imposed by mammalians, is environmental stressors constituting evolutionary pressures. Adaptive capability to manage the pressure enhances the survival of an individual and therefore is selected. Reasonably, fighting could be expected to be able to improve the immune response to microbes as well as vaccines that mimic the microbes, since fighting, more likely to occur in males, usually wounds the involved individuals, especially their skin. The wounded skin is very likely to become contaminated by pathogenic microbes that cause infections and induce immune responses. Presumably, if fighting is linked to the increased immune response to infections, the capability of an enhancing immune response when fighting would therefore be adaptive and selected to increase evolutionary fitness (24). The presumption could be rebutted by the observations that perceived stress reduced the antibody response in caregivers inoculated with influenza vaccines (25), life event stress was likely to induce an inadequate antibody response to HBV vaccine (2), and that acute stress in healthy graduates inhibited the secondary antibody response to hepatitis B vaccinations (15). In contrast, the presumption could be partially supported by the observations that divorced men had a higher antibody titer against both Epstein–Barr virus and herpes simplex virus (14), and that men who had undergone acute stress exhibited an enhanced antibody response to meningococcal A vaccine (6). Accordingly, both intense exercise and psychological stress during fighting should contribute to an enhanced antibody response to vaccine in male mice.
This study also explored why fighting enhanced the antibody response to vaccine in male mice. Previous work showed that an antibody response to antigens such as Candida albicans in humans was specifically influenced by sex steroid hormones, being enhanced by progesterone and estradiol at low levels but depressed by testosterone (16,13), implying that fighting might influence the production of sex hormones, consequently leading to an enhanced antibody response to vaccines. To confirm this, serum testosterone and estradiol were detected in male BALB/c mice immunized with the HBV vaccine, but not statistical significant difference was found at the levels of two sex hormones between the fighting mice and the mice without fighting experience, suggesting that sex hormones might not be involved in the fighting-enhanced antibody response to vaccines. Because that exercise and psychological stress can activate the hypothalamic–pituitary–adrenal (corticosterone) response and the sympathetic nervous system (norepinephrine and epinephrine) response, resulting in a range of rapid physiological and behavioral responses(10,12), next the study tested whether fighting, a behavior complicated with intense exercise and psychological stress, might affect antibody production through the stress hormones. It was observed that administration with hydrocortisone or norepinephrine by intravenous injection to the tail significantly decreased rather than increased the levels of HBsAb to HBV vaccine in both the male and the female BALB/c mice. The observation was in accordance with reports showing that elevated cortisol was negatively related to the antibody response to influenza vaccination in humans (21), and that decreased levels of specific antibody to ovalbumin were negatively correlated with increased levels of plasma norepinephrine, epinephrine, and corticosterone in rats experiencing emotional stress (26). Obviously, the fighting-enhanced antibody response to HBV vaccine in the male mice could not be related to the increased production of stress hormones. In addition to the sex hormones and stress hormones, the study examined how fighting could impact immune cells in male mice. It was found that fighting tended to increase the ratios of CD80+CD11c+ and CD86+CD11C+ cells in the PBMCs of male mice and that CD80 in CD11c+ cells was significantly upregulated in HBsAg co-cultured spleen cells from the fighting-induced mice. Thus, the observations imply that fighting can somehow activate dendritic cells to enhance their capacity to present antigen, such as HBsAg, and therefore promote an antibody response to the antigens.
Footnotes
Acknowledgments
The authors are grateful to Yanmei Wang and Peiyin Zhang for their technical support. This study was supported by the National Nature Scientific Foundation of China (81471888 and 31270982).
Author Disclosure Statement
No competing financial interests exist.