
Abstract
Dengue virus (DENV) is an arthropod-borne virus, which belongs to the Flaviviridae family, and completes its life cycle in two hosts: humans and mosquitoes. For DENV maturation, the surface pre-membrane (prM) protein is cleaved to form a mature membrane protein (M) by furin, which is a cellular enzyme subsequently releasing the mature virus from the host dendritic cell. The objective of the current study was to inhibit mature DENV isotype 2 (DENV2) by RNA-interference in a Vero-81 cell line. Mature DENV2 was propagated in and isolated from U937 cells expressing dendritic cell-specific intracellular adhesion molecule-3-grabbing non-integrin. Maturation of DENV2 was confirmed by Western blot analysis, where virus stock lacking prM was considered mature. Inhibition studies were carried out by transfection of Vero-81 cells with six synthetic siRNAs along with a control siRNA. Reduction in cellular DENV2 was observed also by focus-reduction assay, immunofluorescence assay (IFA), and real-time quantitative polymerase chain reaction (RT-qPCR). Cells transfected with DENV2SsiRNA2, which was targeting the structural region M of mature DENV2, was able to reduce DENV2 titer by up to 85% in focus reduction assays. A significant reduction in mature DENV2 RNA load was observed by RT-qPCR, confirming the previous findings. IFA also revealed reduced levels of cellular DENV2. These results demonstrated that mature DENV2 can be effectively inhibited by synthetic siRNA targeting the structural region of the genome. Mature DENV2 can be successfully inhibited by siRNAs, and specifically high knock-down efficiency is observed by siRNAs against M region of mature DENV2. This study shows that M represents a potential target for RNAi based inhibitory approaches.
Introduction
D
Historically, RNAi was successfully able to limit viral replication, as demonstrated by several RNAi studies targeting major human viral pathogens such as hepatitis B virus, hepatitis C virus, DENV, West Nile virus, Japanese encephalitis virus, and influenza A virus, which led to a significant reduction in virus replication (10). In one of the recent studies, the siRNA and intracellular RNAi mechanism synergistically reduced disease severity associated with DF, proving that RNAi has potential as a potent antiviral therapeutic agent (22). The immature virions present in the ER contain E protein, associated with a pre-membrane (prM) protein in the form of a heterodimer. In each of the immature DENV virions, approximately 60 heterodimers are present, thus differentiating the immature from the mature DENV virions (14,30). Structural analysis of both mature and immature virions revealed that both have an icosahedral symmetry. However, the surface of a mature virion is smooth in comparison to the spiky surface of the immature form (14,30). The conversion of immature to mature DENV occurs in the trans-Golgi network, aided by the change in pH of the environment. As the pH drops in the trans-Golgi network, structural changes allow a cellular cleavage enzyme (furin) to cleave the prM, resulting in virus maturity (28).
Although most viruses have an efficient mechanism for viral assembly and maturation, DENV has a slightly inefficient maturation process (12). Electron microscopy illustrates significant differences in the surface structures of mature and immature DENVs. These differences at the conformational level stabilize the structural elements responsible for proper conversion of DENV morphology during various stages of the DENV life cycle (14). During viral infection in insect cell lines (C6/36) and mammalian cell line (Vero-81), many immature viruses are released because of the limited presence of the furin enzyme (25). Previous studies have shown that siRNAs targeting prM and E regions in the genome of both DENV2 and DENV3 effectively inhibited replication (21,27). A considerable reduction in DENV2 titer was observed in cells that were transfected with adeno-associated virus (AAV) vectors having short hairpin RNA (2).
The objective of the present study was to examine the propagation of DENV2 in a mammalian cell line that readily allows virus maturation of the U937 dendritic cell-specific intracellular adhesion molecule-3-grabbing non-integrin (DC-SIGN) cell line. These cells are rich in furin, which allows maximum DENV particles to mature, and permit the study of the effects of later inhibition of virus maturation via the RNA interference.
Material and Methods
Viruses and cell lines
DC-SIGN-U937 cells were maintained in RPMI complete media (RPMI 1640 supplemented with 1% L-glutamine, 1% penicillin/streptomycin/fungizone, 1% non-essential amino acids, 50 mM 2-mercaptoethanol, and 10% fetal bovine serum [FBS]; Gibco/Invitrogen) and kept in an incubator at 37°C with a constant supply of 5% CO2. Vero-81 cells were grown in Dulbecco's modified Eagle's medium (DMEM; Cellgro) in addition to 10% heat-inactivated FBS and Penstrep (Gibco; 100 IU/mL penicillin and 100 g/mL streptomycin). Vero-81 cells were incubated overnight at 37°C in a humid atmosphere, and CO2 was maintained at 5%. DENV2 stock (DENV2, strain S-16803) was obtained from Dr. De Silva's lab (University of North Carolina at Chapel Hill, NC).
Virus propagation and isolation
Seven days post-infection, media were harvested from cells and clarified by centrifugation at 10,000 g for 30 min. The virus-containing supernatant was supplemented with 20% FBS and stored at −80°C. For studies requiring purified and concentrated virus, the supernatant was centrifuged at 76,221 g for 5 h to pellet down the virus under a 20% sucrose/phosphate-buffered saline (PBS; w/v) cushion. The pelleted virus was resuspended in PBS, loaded onto a 15–60% (v/v) continuous iodixanol gradient, and centrifuged at 29,331 g for 154 min. Fractions containing the purified virus were diluted in PBS and centrifuged at 76,221 g for 5 h to pellet the virus and remove the iodixanol. Purified virus stocks were stored at −80°C for later use in various siRNA study assays.
siRNA designing
siRNAs were designed for targeting DENV2 by applying the Dharmacon algorithm (Dharmacon siRNA Design Center). This algorithm was used to produce effective anti-DENV2 siRNAs, which were later used in various assays to check the inhibition capabilities of selected siRNAs in the context of anti-dengue therapeutic development. While designing siRNAs, important parameters such as length, 3′ overhangs, GC content, and base pair mismatches were considered to obtain a maximum efficiency of the siRNAs (5,20). To have a stable siRNA, duplex GC content was maintained between 30% and 60%. To enhance the efficiency of RNase III, care was taken to ensure that there were four or more identical consecutive nucleotides. siRNAs were designed to facilitate the entry of guide strand in the RISC thermodynamically. It was also confirmed that these siRNAs were not targeting any other organism. Therefore, each siRNA was tested for off-target effects. Finally, siRNA were also checked and cleared of any stretches of nucleotide that can potentially activate immunity.
After a detailed analysis, the Dharmacon algorithm produced many sequences based on several factors and probability to produce effective siRNAs (data not shown). Six non-overlapping sequences that were at the top of the scoring list were selected (Table 1). Further alignment with the complete genomes of reference strains was performed via TCoffee online software (
Transfection (siRNA) of cell culture
The reverse transfection technique was applied to transfect Vero-81 cells seeded in 48-well plates. 10 μM of siRNA taken from the siRNA stock and 1 μL of RNAiMAX (Invitrogen) were mixed in 100 μL of opti-DMEM under standard room-temperature conditions and added to the respective wells of the 48-well plates. Subsequently, 100 μL of DMEM supplemented with 2% FBS containing 25,000 Vero81 cells were added to each well. Cells were incubated for 2 days at 37°C in a CO2 incubator to be later challenged by DENV2.
Western blot analysis
Western blot was performed by initial electrophoresis (Bio-Rad Mini protein3) of the samples on a 12% separating gel. After washing, the membrane was blocked by blocking buffer (2% skim milk in Tris-buffered saline with 0.05% Tween). Primary staining was performed with an anti-DENV2-envelope antibody 8A1 (1:1,000 dilution; gifted by De Silva's lab). After the primary antibody, the membrane was stained with secondary anti-human prM monoclonal antibodies (mAbs; mixture of several prM mAbs at 1:1,000 dilution for maturation studies; gifted by De Silva's lab) for 1 h at room temperature with a very slow speed of rotation on an orbital shaker. The membranes were then washed three times with washing buffer at an interval of 4 min. Final staining was performed by horseradish peroxidase (HRP)-conjugated goat anti-mouse secondary antibody (gifted by De Silva's lab), followed by washing three times and finally developed on an X-ray film.
Focus assay
Vero-81 cells transfected with siRNA in 48-well plates were used to perform focus reduction assays by adding DENV2, diluted in 100 μL/well medium, to obtain an appropriate number of foci in each of the wells (50–60 foci/well). Afterwards, 100 μL of diluent DMEM was added to each well to cover the entire surface area of the well. The cells were then incubated at 37°C on a shaker for 1 h. After 1 h of incubation, DMEM was removed, and the cells were overlaid with 1 M 1% methylcellulose in Opti-MEM (Gibco), 2% FBS (Cellgro), and Penstrep (Gibco). The cells were then incubated at 37°C in 5% CO2 incubators. On day 3 post-infection, methylcellulose-containing overlay medium was removed, and the cells were fixed with 80% methanol in 1× PBS. After air-drying the plates, each of the wells was blocked by blocking buffer (5% instant milk in PBS). The cells were washed again with anti-flavivirus mAb (4G2) (gifted by De Silva's lab) diluted 1:60 in blocking buffer, with a 1 h incubation, on a shaker at 37°C. The cells were washed twice with PBS, incubated with HRP-conjugated goat anti-mouse Ab (Sigma), and diluted 1:500 in blocking buffer at 37°C for 1 h on a shaker. In the final step, the cells were washed again, and 150 μL/well of TrueBlue HRP substrate (KPL) was added. Foci counts were performed on a light box. These counts were then used for virus titer calculations by standard methods (7).
Real-time quantitative polymerase chain reaction
Viral RNA was extracted using a standard Qiagen kit (QI Aamp Viral RNA Kit, cat. #52904). DENV2 RNA was subjected to real-time quantitative polymerase chain reaction (RT-qPCR; CFX96TM detection system) using an iScriptTM RT-PCR kit with SYBR Green according to available protocols (1). Three replicate experiments were conducted for each of the samples, and a 18S rRNA control RNA was also used.
Immunofluorescence assay
From an area of 1.5 cm×1.5 cm (6-well plate), Vero-81 cells were washed with 1 mL medium and detached with a cell scraper. The scraped cells were then centrifuged at 2,500 g for 3 min. A pellet was obtained, and 10 μL of pellets were resuspended in 500 μL of 1× PBS, which was spread in each well of poly-L-lysine treated slides (coated with 0.0005% poly-L-lysine diluted in 1× PBS). After air-drying, the slides were fixed in 100% acetone. Pre-staining was done with diluted anti-prM 2H2-488 antibody (1:200; gifted by De Silva's lab) in block solution. Final washing was performed by adding 35 μL of 1× PBS to each well with 30 min of incubation in the dark. A single drop of mounting fluid (Aqua Poly/Mount 18606; Polysciences Inc.) was applied on each spot in each well. Lastly, each well was covered with a mounting slide (Corning 22 mm×50 mm covers all 10 wells) and observed under a fluorescent microscope. The images were obtained by using and Olympus IX81 motorized inverted microscope, and auto-focusing parameters were set via Metamorph software. Finally, DENV quantification was performed using CellProfiler software.
Results
Effective siRNA designing and synthesis
siRNAs were designed against three selected regions in the DENV2 genome, targeting the 5′-untranslated region (5′UTR), the 3′UTR, and the structural region, respectively. Considering all parameters, 20 sequences from the DENV2(S-16803) genome were selected for further evaluation. Following a series of stringent assessment procedures, six siRNA sequences were selected that fulfilled all the required parameters for effective siRNA design (Table 1). These six siRNAs were later synthesized and used in inhibition studies in a Vero81 cell line.
Western blotting analysis
DENV2 were propagated in U937 DC-SIGN cells. The expression of both E and prM proteins was analyzed for determination of virus maturity (shown in Fig. 1). Lanes 1–3 are DENV2 stocks propagated on C6/36 cell lines. These lanes exhibit an E protein band in combination with a prM band, demonstrating that the majority of DENV2 particles are immature. However, lanes 4–6, which are lacking a prM band, contain DENV2 stocks propagated on U937 DC-SIGN. This shows that approximately all DENV2 particles propagated in U937 DC SIGN are fully mature due to efficient cleavage of the prM protein by the cellular enzyme furin, which is present in abundance in U937 DC-SIGN cells.
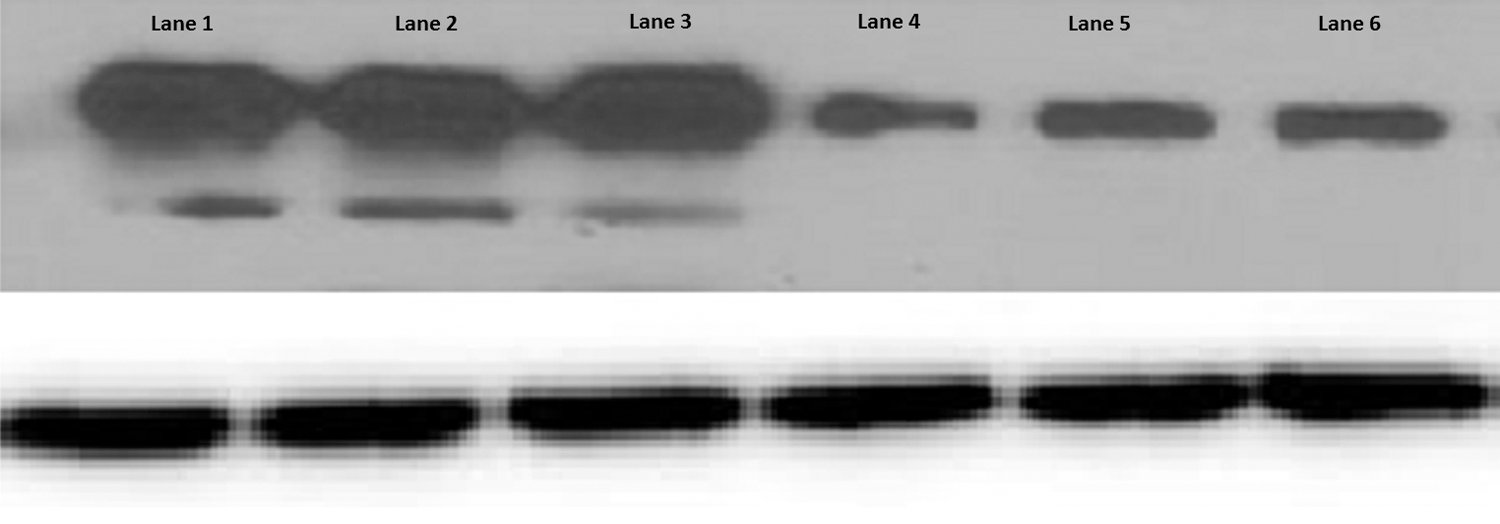
Western blot analysis of mature and immature dengue virus isotype 2 (DENV2). Top image: Western blot was performed by using an anti-envelope (E) protein 8A1 antibody and anti-pre-membrane (prM) mixture of monoclonal antibodies (mAbs). The first three lanes (1 –3) show DENV 2 grown on C6/36 cell line having both E and prM bands, while the remaining three lanes (4 –6) exhibit DENV2 grown on U937 dendritic cell-specific intracellular adhesion molecule-3-grabbing non-integrin (DC-SIGN) cells lacking a prM band. Bottom image: DENV2 E-protein as a loading control. The experiment was repeated twice showing similar results.
Focus assay
Vero-81 cells transfected with each of the six produced siRNA sequences were challenged with DENV2 at a dilution sufficient to obtain the required number of foci (in a range of 50–60 per well). On day 4 post-infection, focus-reduction assay (FRNT) was conducted in which most of the siRNAs were found to be ineffective, since the respective wells exhibited high viral titers. Only DENV2SsiRNA2 was able to reduce DENV2 titer significantly, with more than a 80% decline in viral titer detected. DENV2SsiRNA1 and DENV2UTR3′siRNA2 also demonstrated a reduction in DENV2 titers by 50% and 35%, respectively (Fig. 2). These results emphasize that siRNAs targeting the structural region of DENV2 can effectively inhibit the maturation of DENV2 in Vero-81 cells, while siRNA from the UTR regions were not as effective. Focus count revealed that siRNAs targeting the structural regions of the DENV2 genome can consequently halt DENV2 replication in Vero-81 cells.
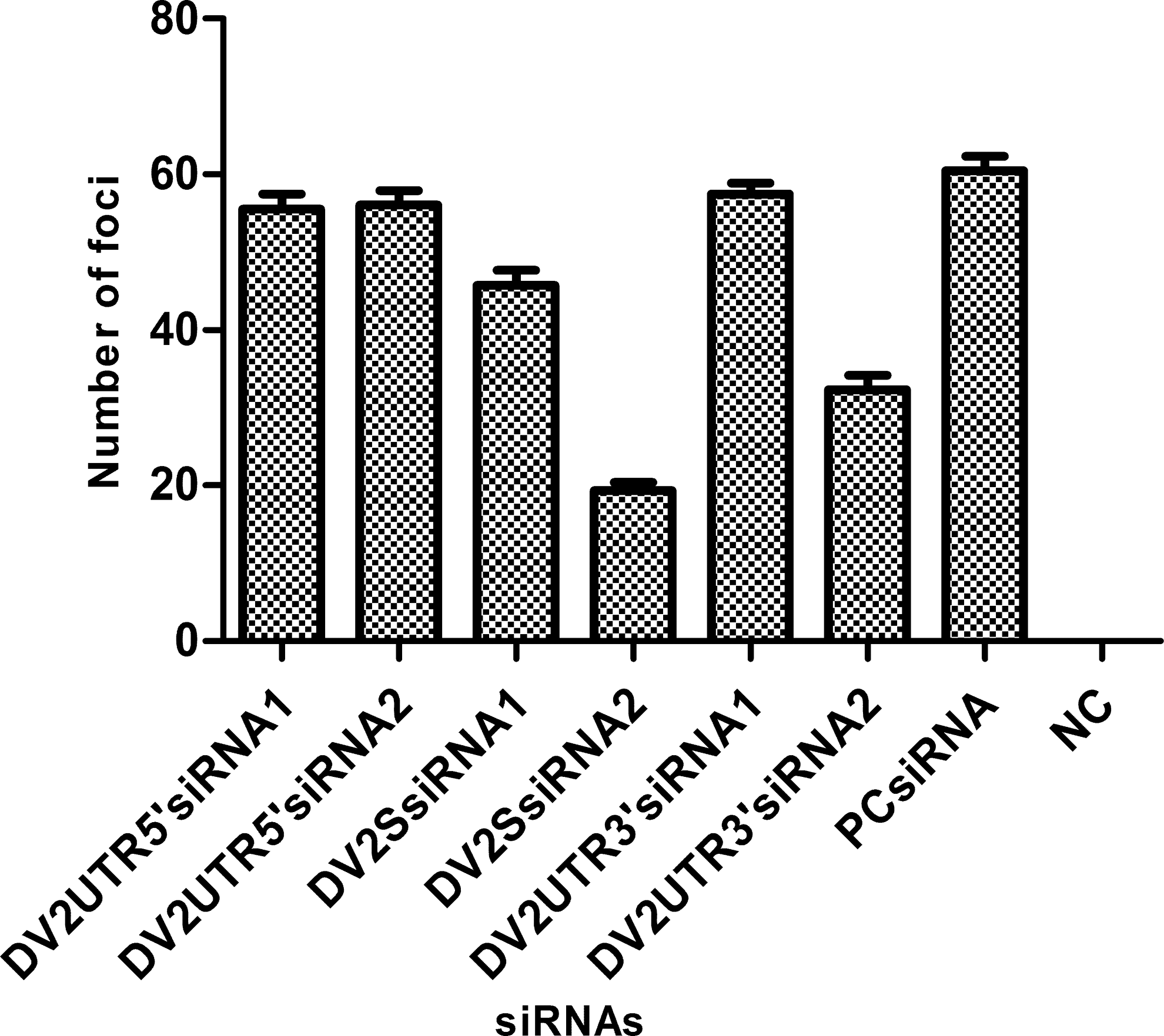
Focus assay of siRNA-transfected Vero-81 cells. The focus assay was conducted on Vero-81 cells transfected with the six selected anti-dengue synthetic siRNAs (displayed in Table 1). Focus assays revealed that siRNAs from the 5′UTR (DENV2UTR5′siRNA1 and DENV2UTR5′siRNA2) had no effect on virus titers, and steady virus replication was observed as shown by bars 1 and 2. DENV2SsiRNA2, which targets the structural region of DENV2 genome, was able to achieve maximum reduction in DENV2 titer, while a slight decrease was also observed with DENV2SsiRNA1 (bars 2–3). DENV2UTR3′siRNA2, designed from the 3′UTR managed to limit DENV2 replication to some extent. Experiments were repeated with similar results, thus reducing the chances of error.
Immunofluorescence assay
Immunofluorescence assay (IFA) is one of the standard methods for checking viral titers in various cell lines. In the present study, seven different siRNA were transfected in Vero-81 cells seeded in 6-well plates. Each siRNA sample was replicated to achieve maximum accuracy, and IFA was performed after an interval of 48 h post-infection. After performing IFA, cells were scraped from individual wells and later stained and visualized under a fluorescent microscope (Fig. 3). Positive control cells having siRNA not related to the DENV2 genome exhibited the maximum DENV2 titer, while negative control cells having no DENV2 were virus free (Fig. 3). In Figure 3, it can be validated that the majority of the siRNAs were ineffective in reducing mature DENV2 titers. However, the lowest viral titer (about 90–95% reduction) was observed in cells transfected with DENV2SsiRNA2. Although DENV2SsiRNA1 and DENV2UTR3′siRNA2 showed a slight reduction in virus replication, a marked decline was not observed.
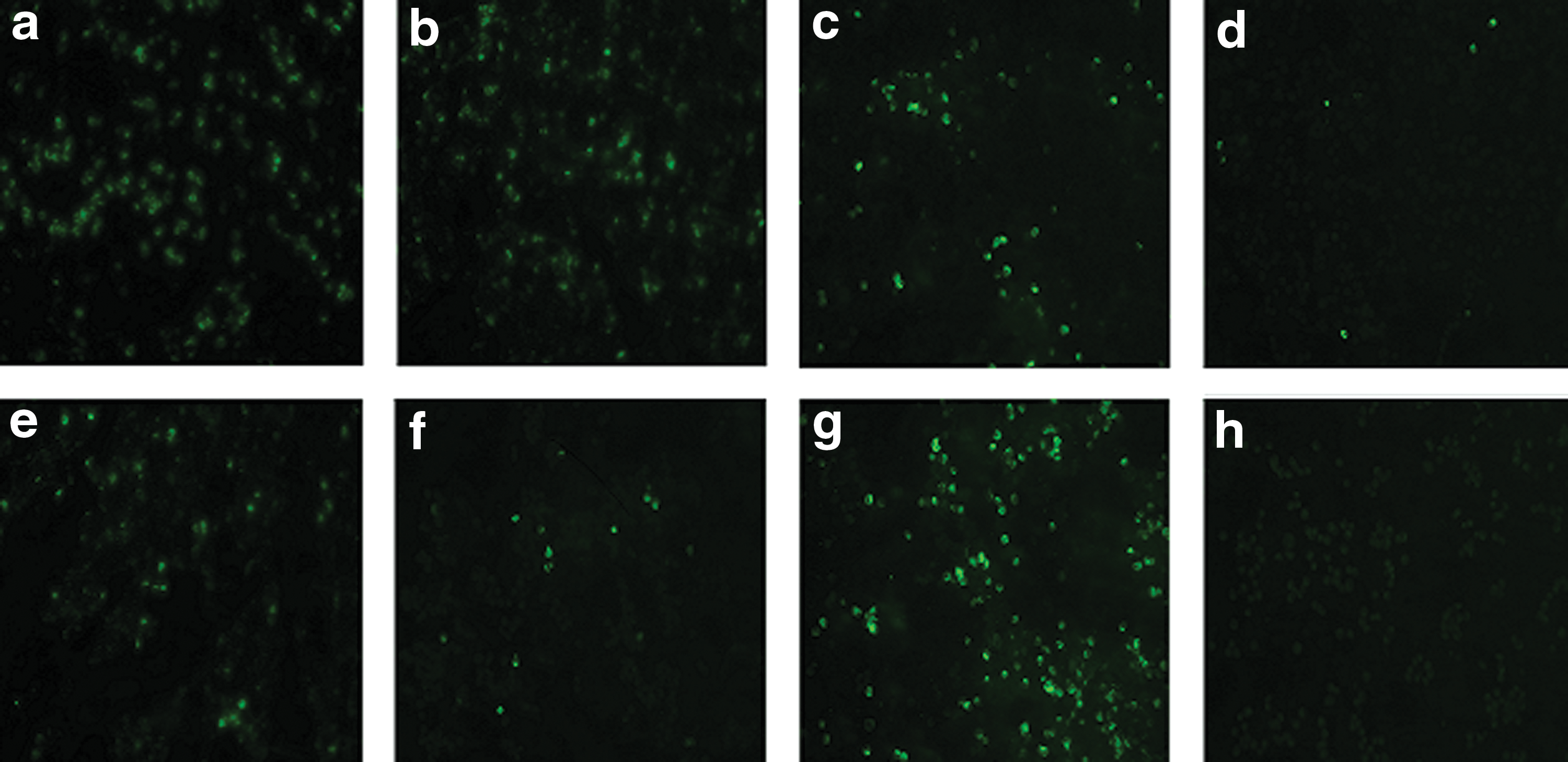
Immunofluorescence assay (IFA) images of siRNA-treated Vero-81 cells. IFA was done 48 h post-infection with anti-E antibody (2H2-488) for monitoring cellular levels of DENV2 E-protein. (
RT-qPCR of DENV2 RNA
In the next step, mature DENV2 RNA was quantified by RT-qPCR. RNA levels were constant except for DENV2SsiRNA2, DENV2SsiRNA1, and DENV2UTR3siRNA2. Similar to the results of the IFA assay (Fig. 3), in the qPCR of cells transfected with DENV2SsiRNA2, the DENV2 RNA copy number was significantly reduced (by more than 90%), whereas DENV2UTR3siRNA2 was able to drop DENV2 RNA by up to 40%, respectively (Fig. 4). These observations suggest that siRNA targeting the structural regions of mature DENV2 can efficiently hamper viral replication.
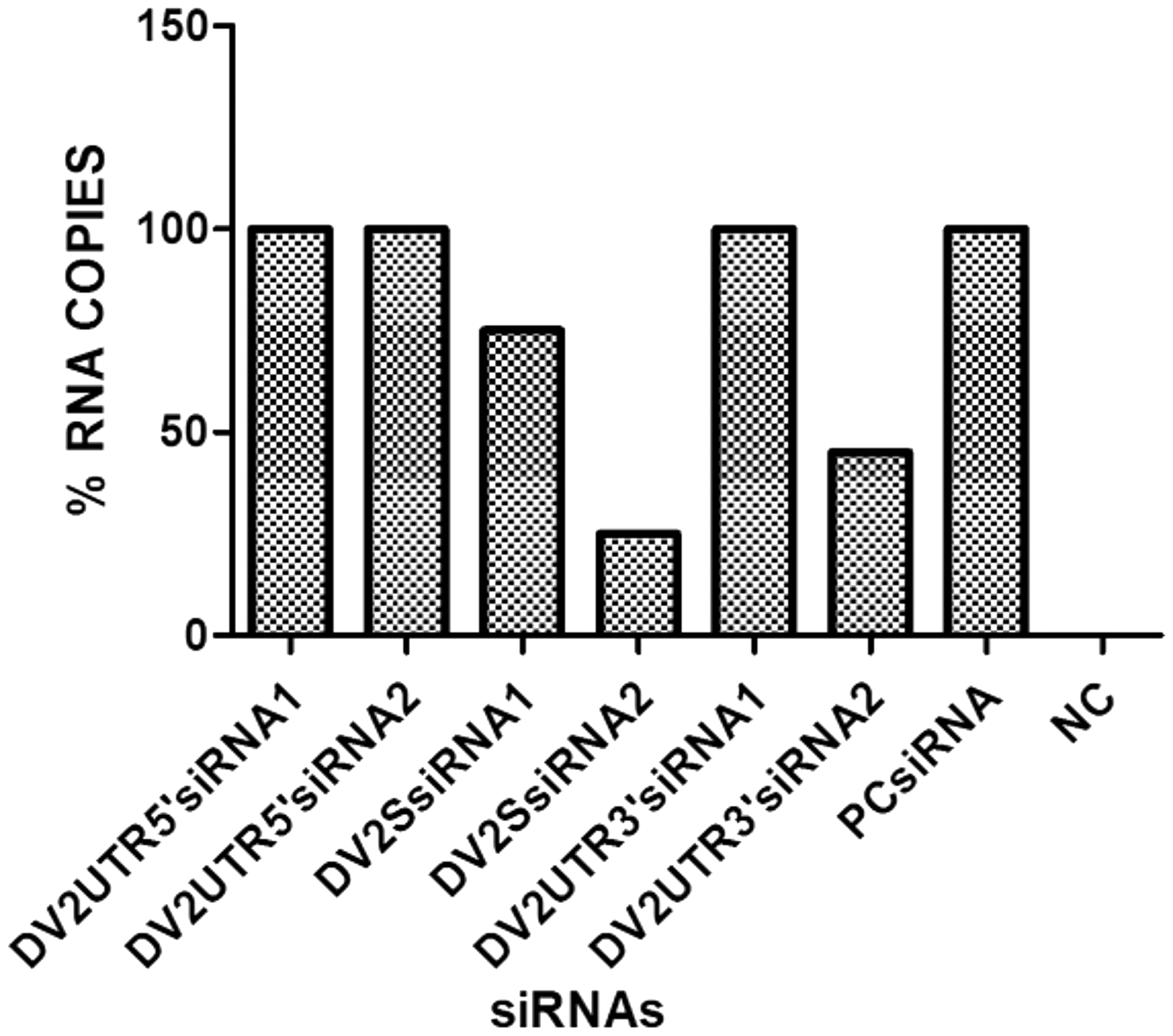
Real-time quantitative polymerase chain reaction (RT-qPCR) for checking DENV2 RNA levels. Quantitative analysis of DENV2 RNA levels was performed by RT-qPCR. RNA from the cells was extracted 48 h post-infection for each individual sample. From left to right, bars 1 and 2 show no significant decrease in DENV2 RNA levels. Bars 3 and 4 show a steady decrease in DENV2 RNA, with DENV2SsiRNA2 showing least DENV2 RNA. Bar 5 is uniform without any prominent change in DENV2 RNA, while bar 6 shows almost a 60% reduction in DENV2 RNA. The last bar represents PCsiRNA (which is a plant origin siRNA and used as a negative control).
Discussion
The current study aimed to find a means of inhibition of mature DENV2 in Vero-81 cells. Earlier RNA interference studies concerning DENV have been conducted on immature DENVs grown on C6/36 cells. Reports from recent studies suggest that the majority of DENV produced in C6/36 are immature due to inefficient cleavage of the prM to M (11,26). During in vitro conditions, DENV tend to produce more mature viral particles in DC cells because of the presence of adequate furin enzyme needed for DENV maturation (16). In the current study, DENV2 was grown on U937 DC-SIGN cells, which resulted in a maximum yield of mature DENV2. Assays using anti-E and anti-prM antibodies confirmed the presence of both proteins, where the presence of prM band was considered as an indication that DENV2 particles were not fully mature, while the absence of prM was indicative of a mature DENV2 (Fig. 1). The mature DENV2 stock that was obtained was later used for RNA interference studies. According to Richter et al., both mature and immature DENVs are equally infectious in nature, but mostly mature DENVs are released from monocytes and DCs during the life cycle of DENV (13). Therefore, to address this issue, inhibition studies were conducted on mature DENV2 in order to maximize the efficacy of this anti-dengue therapeutic strategy.
The RNAi technology is currently being explored as an effective tool of modern molecular biology aimed at silencing of a single or multiple genes in order to curtail events such as the development of cancer or replication of the virus in host cells (24). Endogenous mRNA degradation via siRNA can effectively achieve gene silencing. Many pathogens, including viruses, can be targeted via RNA interference inside the host cells, thus limiting the ability of the pathogens to multiply and cause severe infections. Several RNA-genome viruses have their genome exposed at some time during the process of replication and protein synthesis. This naked genome (RNA) can serve as a potential target for siRNA-based inhibition (15). In the current study, synthetic siRNAs were designed from three regions of the DENV2 genome: the structural region, the 5′UTR region, and the 3′UTR region. Untranslated regions are important components of the DENV genome. The 5′UTR of DENV is a short structure with 100 nucleotides, having a 5′cap, while the 3′cap UTR is relatively larger with 450 nucleotides and has several conserved structures (9). The 5′UTR has two major domains that are involved in viral RNA synthesis and replication (8). The 3′UTR is crucial in DENV replication (17, 23). A few studies suggest that deletions in the 3′UTR can result in defective virus progeny (23). The significance of 5′UTR and 3′UTR makes these regions desirable targets for RNA interference. Consequently, siRNAs were designed from these UTRs to inhibit mature DENV2 in Vero-81 cells. Structural proteins are important for DENV maturation, as well as for transmission, and therefore the structural region was also targeted for inhibiting mature DENV2. Among the designed siRNA, DENV2UTR3′siRNA2 was able to decrease DENV2 titers in the range of 50–60%, and though it proved to be effective, it did not completely inhibit the maturation of DENV2. The results demonstrated that DENV2SsiRNA2 targeting the structural region was able to inhibit mature DENV2 considerably, with up to a 90% decrease in mature DENV2 titers seen in cells transfected with DENV2SsiRNA2.
There can be multiple reasons for the lower effectiveness of siRNAs targeting the UTRs. According to some existing studies, there are several conserved hairpin structures located in the UTRs that can hinder the cleavage efficiency of siRNAs due to unavailability of proper binding sites for the guide strand in RISC (29). Ching-Fang et al. also suggested in 2013 that thermodynamically siRNAs designed from coding regions were more efficient in comparison with siRNAs targeting UTRs (6). A recent study has shown that when AG129 mice were challenged with DENV2, the group of mice that underwent intravenous anti-dengue siRNA inoculation survived. These results demonstrate that RNAi can be an important candidate for treatment of DENV in the future (22). The ability to inhibit DENV2 effectively in the current study can be further evaluated in animal model in future studies.
Conclusion
This article uncovers the role of RNA interference in the inhibition of maturation of DENV2 in mammalian cells. One of the six siRNAs has shown promising potential for future inhibitory studies. Recent advances in drug development have helped in targeted delivery, as well as in sustained release of siRNAs. These advances in drug delivery utilizing liposome and nanoparticle technology have significantly increased the importance of RNA interference. The use of mature DENV makes this study vital in reference to DENV replication in the host (human) cells. DENV2SsiRNA2 can be an important candidate for anti-dengue drug development. Consequently, siRNA technology is a potent therapeutic method in the quest for achieving a drug for the treatment of DENV. This study will pave the way for future studies on DENV maturity and factors associated with controlling DENV infections in human hosts.
Footnotes
Acknowledgments
Financial support for the conduct of this study was provided by National University of Sciences and Technology (NUST), H-12, Islamabad, Pakistan, and the Higher Education Commission of Pakistan.
Author Disclosure Statement
No competing financial interests exist.