
Abstract
A role for Epstein-Barr Virus (EBV) infection in the etiology of autoimmune diseases, including rheumatoid arthritis (RA), has long been suggested. However, data about EBV burden in RA patients from Sardinian population, a genetic isolate with high prevalence of autoimmune diseases, have not yet been reported. One hundred thirty-five, Sardinian subjects (77 RA patients and 58 demographically matched healthy donors, HDs) were enrolled in a cross-sectional case–control study. EBV-DNA was quantified by quantitative real-time polymerase chain reaction in peripheral blood mononuclear cells (PBMCs). Prevalence and titers of anti-Early Antigen IgG (anti-EA-IgG) and anti-Epstein-Barr Nuclear Antigen 1 IgG (anti-EBNA-1 IgG) were determined by immunoenzimatic assay. EBV-DNA positivity was more frequent in RA PBMCs than in HD PBMCs (79.2% vs. 56.9% respectively, p=0.008). Similarly EBV relative load was increased in RA than in HD PBMCs [2.83 (6.5) vs. 0.53 (1) 2−ΔCt EBV-DNA, respectively, p=0.02]. Moreover, Sardinian RA patients were found to have increased prevalence of anti-EBNA-1 IgG (90% vs. only 69% of HD, p=0.006) and anti-EA IgG (37% compared with only 10.3% of HD, p=0.002). Subgroup analysis revealed that PBMCs from RA receiving Tocilizumab, an anti-interleukin-6 (IL-6) receptor monoclonal inhibitor, have significantly lower EBV viral loads in comparison to PBMCs from RA under other immunosuppressors (p=0.03). These data suggest an association between EBV infection and RA in the Sardinian population. The potential influence of IL-6 inhibition on EBV viral load in RA patients should be further explored in prospective trials.
Introduction
R
Among viruses, a role for the Epstein-Barr virus (EBV), a highly prevalent DNA virus of the herpes family, has long been supposed in RA pathogenesis (10).
EBV infects both B cells and epithelial cells, and can cause infectious mononucleosis in childhood and different malignancies in immunocompromised hosts. After a primary infection at the level of nasopharyngeal epithelial cells, EBV penetrate mature naïve and memory B cells in whom viral genome expression is restricted to maintain a latent asymptomatic lifelong infection. At this stage, a “classical” subset of latent viral antigens are produced, including the Epstein-Barr nuclear antigen-1 (EBNA-1). EBV may reactivate under different conditions, switching from the latent-phase to the lytic one; At this time, several EBV antigens are expressed, including the so called Early Antigen (EA).
Therefore, the presence of anti-EBNA antibodies is a marker of a prior infection, while the presence of antibodies to EA is detectable for only a transient period following primary infection or EBV reinfection/reactivation.
The presence of a potential link between EBV infection and RA is suggested by demonstration of higher EBV-viral load, increased titers of EBV-directed antibodies, and impaired EBV-specific T-cell cytolytic activity in RA patients compared with healthy subjects (4,6,23,27).
CD8+ memory cells are fundamental in controlling EBV-infected B-cell proliferation and EBV reactivation (15,24,28). Uncontrolled growth of EBV-infected B cells may result in an increased incidence of B-cells lymphoproliferative disorders in immunosuppressed patients (15,24). In RA patients, prospective data appear reassuring about the potential of EBV reactivation and tumor lymphomagenesis in subjects receiving methotrexate and antitumor necrosis factor alpha agents (5,21). There is a paucity of informations on the risk of EBV reactivation and EBV-related lymphoproliferative disorders in RA patients receiving new biotechnological drugs, including Tocilizumab (TCZ), an anti-interleukin-6 (IL-6) receptor monoclonal inhibitor.
No data about EBV prevalence in RA population in Sardinia (Italy) are available to date. These data could be of particular interest, considering that Sardinian population is characterized by a homogeneous genetic background with little gene inflow and a peculiar high frequency of autoimmune diseases (e.g., type 1 diabetes, multiple sclerosis, coeliac disease, and myastenia gravis) (7,8,25). Of note, a higher susceptibility to a second autoimmune disease has been demonstrated in Sardinian patients (25), clearly suggesting the presence of a common pathogenic mechanism(s) among autoimmune diseases (9).
We previously demonstrated high prevalence and increased levels of anti-EBNA-1 IgG in Sardinian patients with multiple sclerosis (17). This observation prompted us to evaluate the hypothesis that EBV infection may be one among the missing pathogenic mechanisms common to different autoimmune disease in Sardinia.
Hence, we sought to measure EBV-DNA load and EBV-specific humoral immune response in blood of RA Sardinian patients with respect to healthy donors (HDs). Moreover, we evaluated cross-sectional differences in EBV burden in RA patients according to type of ongoing immunosuppressive treatment.
Materials and Methods
Ethics statement
The study was approved by the Ethics Committee of ASL-1 Sassari ASL (Prot. 1192/L, 2014). The patients and the volunteers gave written informed consent.
Subjects
Seventy-seven consecutive unselected Sardinian patients with RA diagnosed according to the American College of Rheumatology criteria (1) [median (IQR) age 56 (17) years, 63 females] referred to the outpatient clinic of Rheumatology Unit at the Department of Clinical and Experimental Medicine of the University of Sassari (Italy) between March and April 2014 were recruited in this cross-sectional case–control study. Fifty-eight healthy sex and age-matched subjects [median (IQR) age 59 (18) years, 42 females) from local blood donor bank were enrolled in the study as controls (HDs). All patients and controls came from various areas of Sardinia.
For each RA patient enrolled in the study data regarding disease duration, disease activity, functional state, and ongoing treatment were systematically registered. Disease activity was assessed using DAS-28 (Disease Activity Score-28 score) and functional class by HAQ (Health Assessment Questionnaire). Demographic, clinical, and laboratory characteristics of participants were summarized in Table 1.
Continuous data are expressed as median (IQR).
ACPA, anti-citrullinated protein antibody; anti-TNF, antitumor necrosis factor alpha; CRP, C-reactive protein; DAS-28, Disease Activity Score-28; DMARDs, disease modifying antirheumatic drugs; ESR, erythrocyte sedimentation rate; HAQ, Health Assessment Questionnaire; HDs, healthy donors; IQR, interquartile range; RA, rheumatoid arthritis; RF, rheumatoid factor.
Blood cell separation
Peripheral blood mononuclear cells (PBMCs) were isolated from heparinized peripheral blood samples by layering on Ficoll/Hipaque, as described elsewhere (18). Cell viability was assessed using the Trypan blue exclusion method. The cells were either frozen in deep cold, or processed immediately for further studies.
DNA extraction and Epstein-Barr virus real-time polymerase chain reaction
DNA was extracted from 106 cells, by DNAzol as described elsewhere (19). Selective amplification of the EBV DNA was obtained by a real-time polymerase chain reaction (PCR) assay, as previously published (18). The DNA amounts used for amplification were 1 μg/sample, and the PCR temperature was 63°C. For each sample, the Ct (cycle threshold) value of the gene of interest (GI) was normalized by comparison to the Ct of the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) invariate housekeeping gene, assuming that (i) a difference of 1 between sample Cts means that the sample with the lower Ct value had double the target sequence of the other sample, and (ii) the rate of Ct change versus the rate of target copy change is identical for the GI and the housekeeping gene. Data have been expressed according to the 2−ΔCt Method, according to the following formula: DNA BamHI W fragment relative amounts=2−ΔCt, where ΔCt=[CtGI−CtGAPDH].
Enzyme-linked immunosorbent assay
Presence and levels of IgG antibodies against EBV antigen-specific EBNA-1 and EA were evaluated using commercially available kit anti-EBV EBNA-1 and EA IgG enzyme-linked immunosorbent assay (ELISA) (Bio-Rad). The cutoff value was calculated from the mean OD450 value of the negative control plus 0.200, as recommended by the manufacturer, while standards and controls provided by the kit were used to quantitatively assess the results in arbitrary units (RU/mL). The EBV-ELISA KIT (BIORAD) for detection of EBNA-1-specific IgG was carried out with EBV nuclear antigen p72, while for EA IgG antibodies we used p54/p138 EAs.
Statistics
Differences between categorical variables between two groups and multiple groups were analyzed by Fisher exact test and by Fisher exact test with Monte Carlo approximation, respectively. Differences between quantitative variable were analyzed using the Mann–Whitney rank-sum test and the Kruskal–Wallis with Dunn's post hoc test, when comparing two or multiple groups, respectively. Receiver Operating Characteristic (ROC) curves were constructed to evaluate the potential of EBV-antibody testing in differentiating between RA and HD subjects. Bivariate analysis was performed to test association between quantitative variables.
Statistical analysis and ROC curve analysis were performed by SPSS software (release 20.0). A p-value of less than 0.05 was considered statistically significant.
Results
Higher prevalence of humoral response against EBV-antigens in Sardinian RA patients than in HD
Anti-EBNA-1 IgGs were found in sera of 61 out of 68 (90%) RA patients compared with 40 out of 58 (69%) HD (AUC=0.58; p=0.006) (Fig. 1A). IgG response to EA was observed in 25 out of 68 (37%) RA patients compared with only 6 out of 58 HD (10%) (AUC=0.69; p=0.001) (Fig. 1C). There were no differences in anti-EBNA-1 IgG levels between EBV-positive RA patients and HD (Fig. 1B), whereas anti-EA IgG levels were significantly higher in HD than in RA patients [60.9 (38.5) vs. 20.2 (26.3) RU/mL, respectively; p=0.007] (Fig. 1D).
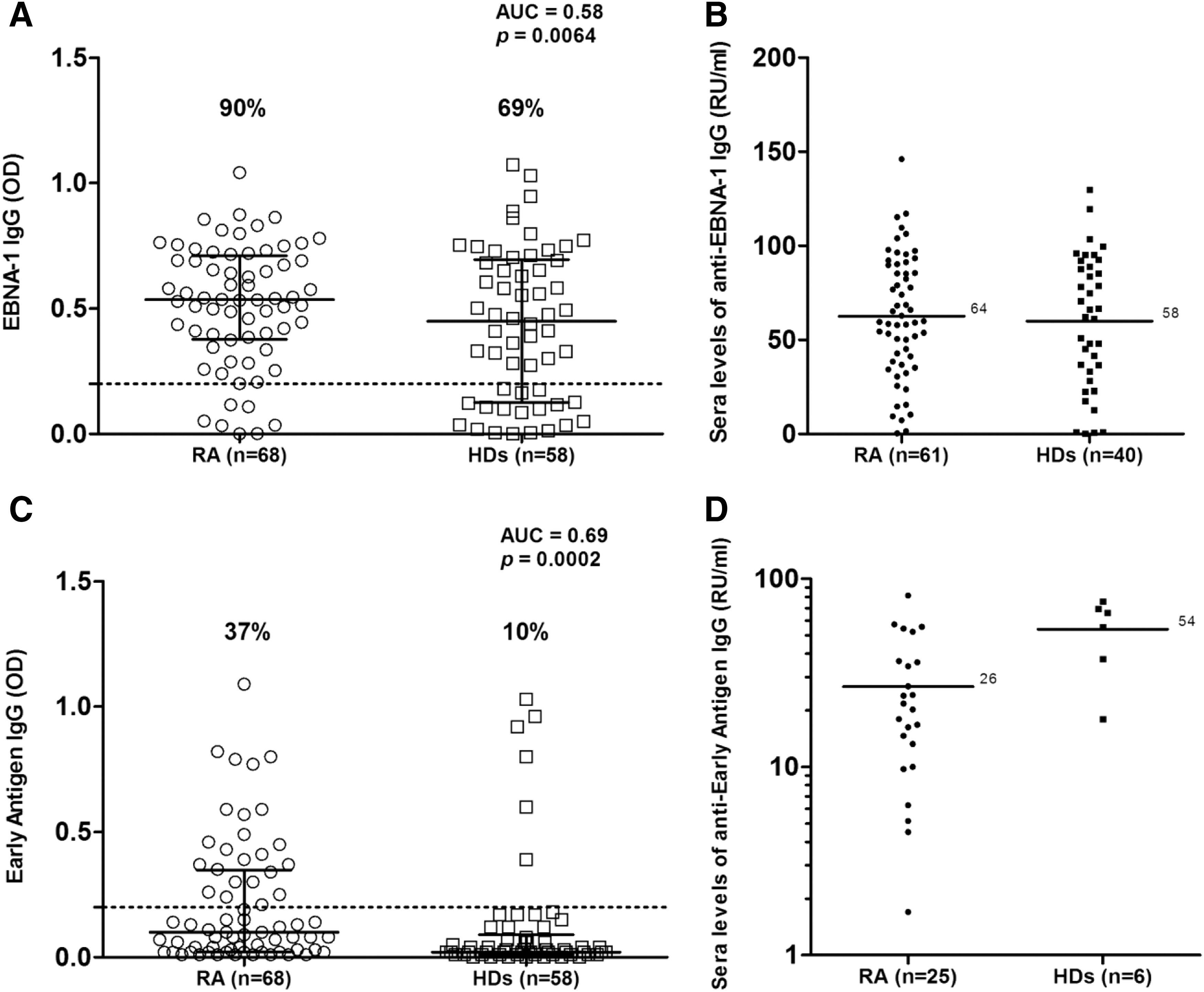
We found no significant association between prevalence and magnitude of humoral response against EBV antigens in RA patients and type of immunosuppressive treatment. Bivariate analysis did not demonstrate correlation between EBV-directed antibodies levels and DAS-28, HAQ, CRP, and erythrocyte sedimentation rate (ESR) values.
Higher prevalence and amount of EBV-DNA viral load in PBMCs from Sardinian RA patients than in PBMCs from HD
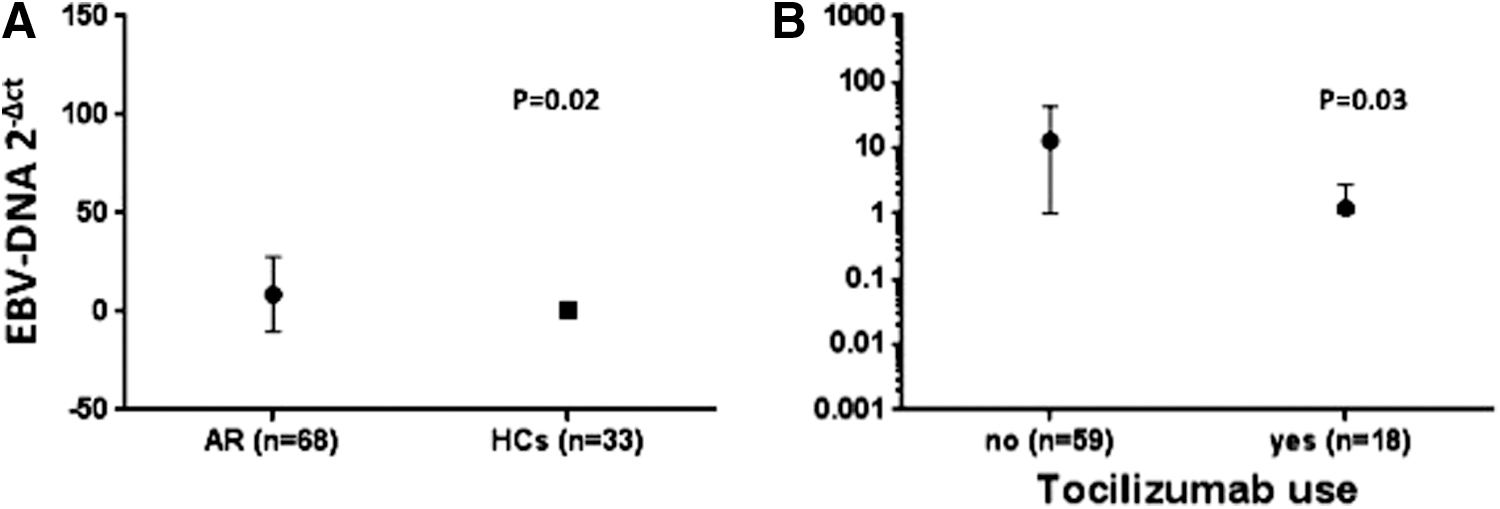
We found EBV-DNA in PBMCs from 61/77 (79.2%) RA patients and in PBMCs from 33/58 (56.9%) HD (p=0.008). Of note, EBV viral load was increased in PBMCs from RA patients with respect to PBMCs from HD [2.83 (6.5) vs. 0.53 (1) 2−ΔCt, respectively; p=0.02] (Fig. 2A).
Epstein-Barr Virus (EBV) load distribution in RA patients according to immunosuppressive treatment.
Lower EBV-DNA viral load in PBMCs of RA patients under Tocilizumab treatment
Subgroup analysis demonstrated significant differences (p=0.007, Kruskal–Wallis test) in EBV viral load across RA groups stratified according to type of immunosuppressive therapy (Fig. 2B). Of interest, lower EBV-DNA levels were demonstrated in RA patients receiving TCZ with respect to RA patients under other immunosuppressive therapy 1.73 vs. 11.48 2−ΔCt, respectively; p=0.03, Mann–Whitney test (Fig. 2B).
Discussion
Uncontrolled EBV infection and replication may trigger autoimmunity in genetic susceptible individuals and may be crucial in multiple sclerosis or RA (23). In this study we investigated for the first time the association between RA and the EBV virus in a large cohort of unselected Sardinian RA patients and HD.
Of note, we found a higher prevalence of antibodies directed to antigens of both latent and lytic EBV phases in Sardinian RA patients compared to HD, which is consistent with previous report (2,22,29).
In agreement with previous reports (3,5), also Sardinian RA patients show increased frequency of EBV-positive PBMCs and raised EBV viral load (about a 6.5-fold change) with respect to HD.
Collectively taken, these data further support the hypothesis that EBV-infection, among the environmental factors, may have a role in the pathogenesis of RA.
Significantly higher rates of anti-EA reactivity in our RA sera may implicate EBV reactivation possibly linked to immunosuppressive therapy. However, a similar higher proportion of anti-EA antibodies positivity respect to controls has been demonstrated in patients with systemic sclerosis and primary anti-phospholipid syndrome free of immunosuppressive therapy (13). Therefore, it is conceivable that EA reactivity reflects an alteration in the relationship between the viral behavior and the altered RA immunological state independent from immunosuppressive treatment.
Of note, although all patients were on immunosuppressive therapy, no cases of overt EBV reactivation were registered in our series. This observation is in line with data obtained in two large prospective cohort of RA patients receiving anti-TNF and methotrexate (5,21), and these are reassuring about the risk of EBV reactivation in EBV-positive RA patients receiving such an immunosuppressive therapy.
We reported for the first time data about EBV-burden and EBV-seroprevalence in RA patients under TCZ, a recombinant humanized monoclonal antibody against the IL-6 receptor: at this regard we demonstrated a significantly lower EBV burden in PBMCs of RA patients receiving TCZ with respect to PBMCs of RA patients receiving other immunosuppressive treatments.
TCZ acts competing for IL-6 receptors and leads to inhibition of the inflammatory response by blocking signal transduction through gp130 receptor. Moreover, IL-6 is a key factor involved in proliferation of B cell and appears to induce EBV replication in B cells (12,26) and to negatively modulate T-cell control of EBV-infected B cells (20). Therefore IL-6 may predispose to development of EBV-related B-lymphoproliferative disorders (BLPD). At this regard, an interventional phase 1–2 study has demonstrated promising results for IL-6 inhibition in the treatment of EBV-related BLPD (27). In this trial 12 recipients of transplanted organs who had BLPD received a monoclonal anti-IL-6 antibody: 8/12 patients entered a condition of clinical remission. Interestingly, the EBV genome was no longer detectable 2–4 months after treatment in the blood of six patients that were EBV-DNA positive before treatment. Moreover, most of the patients in clinical remission were able to develop full antibody responses to EBV, including anti-EBNA antibodies (14).
Of note, Pitzalis and coworkers demonstrated that a large proportion of B cells and anti-citrullinated protein antibody (ACPA)-producing plasma cells in synovial tissue from RA patients were infected with EBV, suggesting a role for the virus in the selection of autoreactive B cell (11). This observation suggest that chronic infection of synovial immune cells by EBV may be involved in autoantibody production and disease pathogenesis
Therefore, it can be supposed that TCZ may exert a favorable effect in the control of RA activity, not only hampering systemic inflammation but also reducing B-cell EBV load and inhibiting the selection of EBV positive ACPA producing plasma cells.
In conclusion, Sardinian RA patients show increased frequency of EBV-directed antibodies and EBV overload when compared with healthy population. In addition, EBV viral load appears to be significantly lower in RA patients receiving TCZ, suggesting a role for IL-6 inhibition in controlling EBV burden. Prospective data in larger cohorts are needed to confirm these data and to evaluate the potential influence of TCZ therapy on EBV replication.
Footnotes
Acknowledgments
This investigation was supported by grants from the Sardinian Region L.R.7 2010 and Fondazione Banco di Sardegna year 2014 to L.A.S.
Author Disclosure Statement
No competing financial interests exist.