
Abstract
Hantaan virus (HTNV) infection can cause severe hemorrhagic fever with renal syndrome (HFRS) in humans. CD14, a pattern recognition receptor recognizing lipopolysaccharide, is highly expressed on monocytes and can be shed as soluble CD14 (sCD14) upon monocyte activation. To understand the role of sCD14 in HFRS, the sCD14 plasma concentrations from 45 HFRS patients were quantified, and the relationships between the plasma sCD14 level and the monocyte response status and clinical parameters were analyzed. The plasma sCD14 levels were significantly higher in the HFRS patients and they correlated with monocyte expansion and activation, which were characterized by increased blood monocyte counts, the proportion of CD14++CD16+ intermediate monocytes, as well as elevated plasma tumor necrosis factor-α (TNF-α) and soluble CD163 (sCD163) levels. Additionally, the high plasma sCD14 levels positively correlated with white blood cell counts and blood urea nitrogen levels and negatively correlated with platelet counts in the HFRS patients. Taken together, our data indicate that elevated plasma sCD14 levels are associated with the monocyte response status during HTNV infection in humans.
Introduction
H
Circulating monocytes play important roles in inflammatory and innate immune responses (2) and they have been characterized into three subsets, classic (CD14++CD16−), intermediate (CD14++CD16+), and nonclassical (CD14+CD16++) monocytes, based on their CD14 and CD16 expression (33,34). The major monocyte population is the CD14++CD16− monocytes, which account for 80–90% of these cells, while the latter two subsets account for ∼15% of the total blood monocytes. These monocyte subsets display different roles in immune defense to infection and tissue repair (30). Interestingly, dramatic monocyte subset changes are observed in viral diseases, such as in dengue fever and HIV, in which expanded CD14++CD16+ monocyte population levels are observed (30).
CD14, a glycosylphosphatidylinositol (GPI)-anchored membrane glycoprotein member, is a pattern recognition receptor that recognizes lipopolysaccharide (LPS) and it is highly expressed on monocytes and macrophages and weakly expressed on neutrophils (1). Monocyte activation-dependent shedding of CD14 yields the vast majority of soluble CD14 (sCD14) (4). Additionally, increased membrane-bound CD14 (mCD14) expression or direct CD14 secretion that bypasses the GPI linkage possibly contributes to the sCD14 variation (16). In recent years, many clinical studies focusing on sCD14 revealed that in infectious diseases, such as sepsis (14) and HIV (24), sCD14 levels were elevated and associated with disease severity or prognosis. However, the role of sCD14 in HFRS has not yet been investigated.
CD163 belongs to the scavenger receptor cysteine-rich superfamily and is exclusively expressed in the monocyte–macrophage lineage (32). Shedding of CD163 yields soluble CD163 (sCD163) in plasma. It was found that plasma sCD163 levels were elevated in HFRS patients and associated with severe acute kidney injury (32).
In this study, we measured the plasma sCD14 level in HFRS patients and analyzed the relationships between sCD14 levels and monocyte response status and clinical parameters. This analysis might help clarify the roles of sCD14 and monocytes in immune response to HTNV infection.
Materials and Methods
Sample preparation
Blood samples were harvested from 45 hospitalized patients with HFRS between 2013 and 2014 at the Tangdu Hospital of the Fourth Military Medical University (Xi'an, China) and 25 healthy donors (normal controls). The clinical diagnosis of HFRS was serologically confirmed by detecting specific IgM and IgG antibodies to HTNV. According to the diagnostic criteria from the Prevention and Treatment Strategy of HFRS promulgated by the Ministry of Health, People's Republic of China, the patients were classified into mild, moderate, severe, and critical, four clinical types (21).
(a) mild: mild renal failure without an obvious oliguric stage; (b) moderate: obvious symptoms of uremia, effusion (bulbar conjunctiva), hemorrhage (skin and mucous membrane), and renal failure with a typical oliguric stage; (c) severe: severe uremia, effusion (bulbar conjunctiva and either peritoneum or pleura), hemorrhage (skin and mucous membrane), and renal failure with oliguria (urine output, 50–500 mL/day) for ≤5 days or anuria (urine output, <50 mL/day) for ≤2 days; (d) critical: for those with ≥1 of the following symptoms: refractory shock, visceral hemorrhage, heart failure, pulmonary edema, brain edema, severe secondary infection, and severe renal failure with oliguria (urine output, 50–500 mL/day) for >5 days, anuria (urine output, <50 mL/day) for >2 days, or a blood urea nitrogen (BUN) level of >42.84 mM.
According to the clinical observation, the illness could be divided into the acute phase and convalescent phase. The patients with other kidney diseases, diabetes, hematological diseases, cardiovascular diseases, viral hepatitis, autoimmune diseases, and other liver diseases were excluded in this study. Plasmas were separated from these blood samples by centrifuge and frozen at −80°C before application, and peripheral blood mononuclear cells (PBMCs) were obtained using density gradient centrifugation at the same time. The local Institutional Review Board approved this study, and all of the subjects signed an informed consent form before their blood was collected.
Enzyme-linked immunosorbent assay
The plasma sCD163 level was quantified, as in our previous study (32). The plasma sCD14 and TNF-α concentrations were quantified with sandwich enzyme-linked immunosorbent assay kits (R&D Systems) according to their protocols. The optical density values were measured at 450 nm with a microplate reader, and the sCD14 and TNF-α concentrations were estimated from their standard curves.
Flow cytometry
Flow cytometry was used to measure the percentages of the three monocyte subsets from PBMC preparations using CD14 (anti-CD14-FITC, clone 61D3; eBioscience) and CD16 mAbs (anti-CD16-PE, clone CB16; eBioscience). Matched isotype controls were used as negative controls. Monocytes were gated based on their forward and side scatter characteristics and were further divided into CD14++CD16−, CD14++CD16+, and CD14+CD16++ monocytes (34). The percentage of each subset was obtained after monocyte gating. The data were analyzed with the FlowJo 7.6.1 software (Oregon).
Statistical analysis
The Mann–Whitney U test was used to compare the plasma sCD14 levels between two independent groups. The Kruskal–Wallis test was used for analysis of differences among multiple groups. The nonparametric Spearman correlation test was used for correlation analysis of the sCD14 levels with clinical parameters or the monocyte response status. Continuous variables were reported as medians with corresponding interquartile ranges (IQR). p-Values (two-tailed) below 0.05 were considered statistically significant.
Results
The clinical information of the subjects
A total of 100 plasma samples from 45 HFRS patients and 25 healthy donors were examined. The information of patient samples is summarized in Table 1.
Thirty patients have samples collected during acute and convalescent phases, 7 patients have samples collected during the acute phase, and 8 patients have samples collected during the convalescent phase.
Dynamic changes of the plasma sCD14 levels in the HFRS patients
The plasma sCD14 levels in the HFRS patients were elevated compared with the normal controls (2,327 [1,734–3,026] ng/mL vs. 1,478 [1,171–1,640] ng/mL, p<0.0001) (Fig. 1A). Considering the different phases, the plasma sCD14 levels were 2.0-fold higher in the acute phase compared with the normal controls (2,927 [2,430–3,388] ng/mL vs. 1,478 [1,171–1,640] ng/mL, p<0.0001). These levels decreased during the convalescent phase (1,894 [1,525–2,287] ng/mL, p<0.0001), but they were still higher than the normal control levels (p=0.0002) (Fig. 1B). The acute and convalescent phase plasma sCD14 levels also showed a declining tendency in the 30 observed patients (Fig. 1C). Between the different severity groups, there were no significant differences in the plasma sCD14 levels (data not shown).
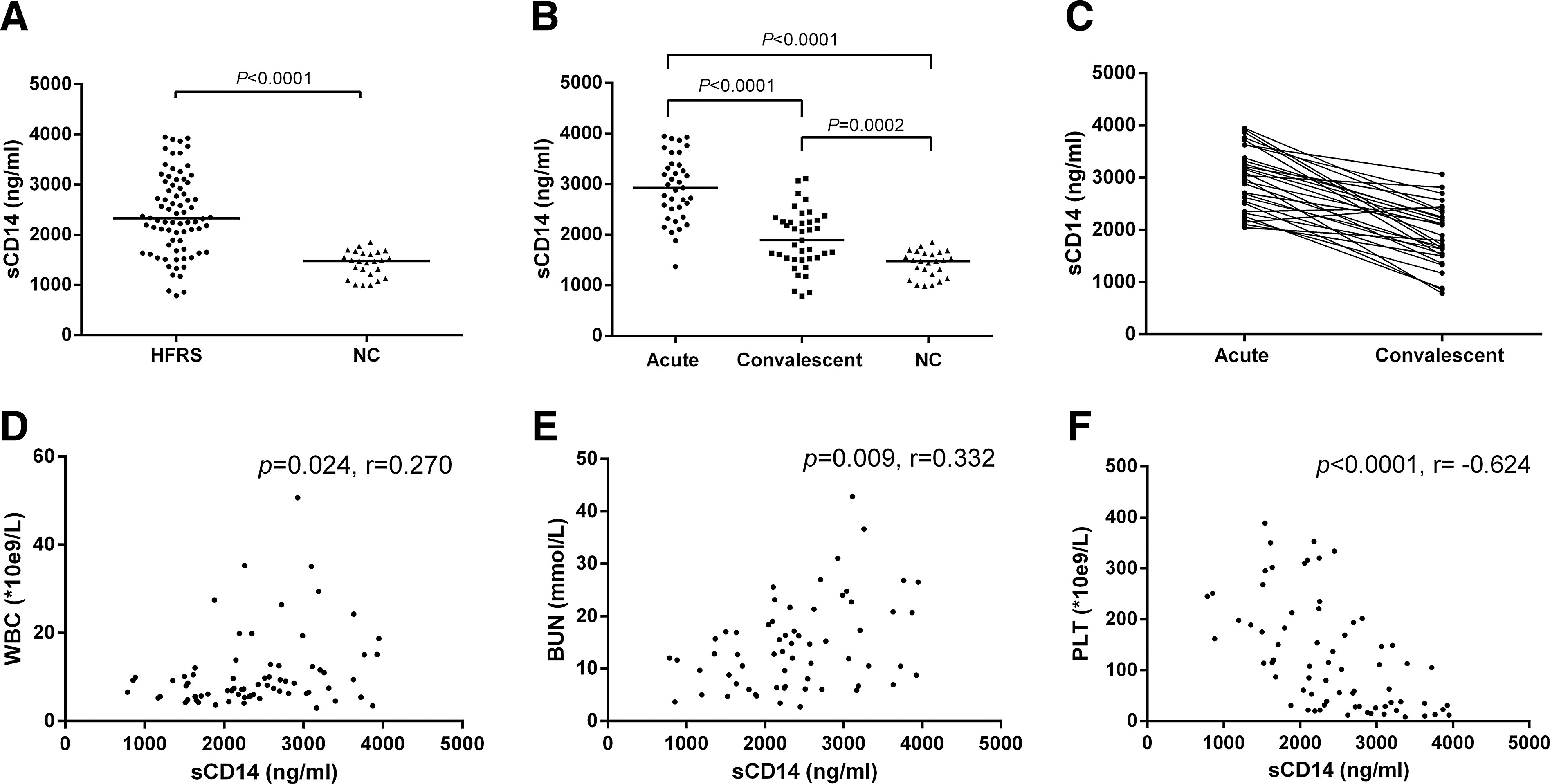
Dynamic plasma soluble CD14 (sCD14) changes in hemorrhagic fever with renal syndrome (HFRS) patients and its correlation with the observed clinical parameters.
The plasma sCD14 levels correlated with the clinical parameters
The plasma sCD14 levels positively correlated with white blood cell (WBC) counts (r=0.270, p=0.024, Fig. 1D) and blood urea nitrogen (BUN) levels (r=0.332, p=0.009, Fig. 1E) and negatively correlated with the platelet (PLT) counts (r = −0.624, p<0.0001, Fig. 1F) in the patients.
The plasma sCD14 levels correlated with the monocyte response status
The elevated plasma sCD14 levels positively correlated with the increased monocyte counts (r=0.527, p=0.010, Fig. 2A left), TNF-α (r=0.462, p=0.002, Fig. 2A middle), and sCD163 levels (r=0.342, p=0.028, Fig. 2A right) that were observed in the patients.
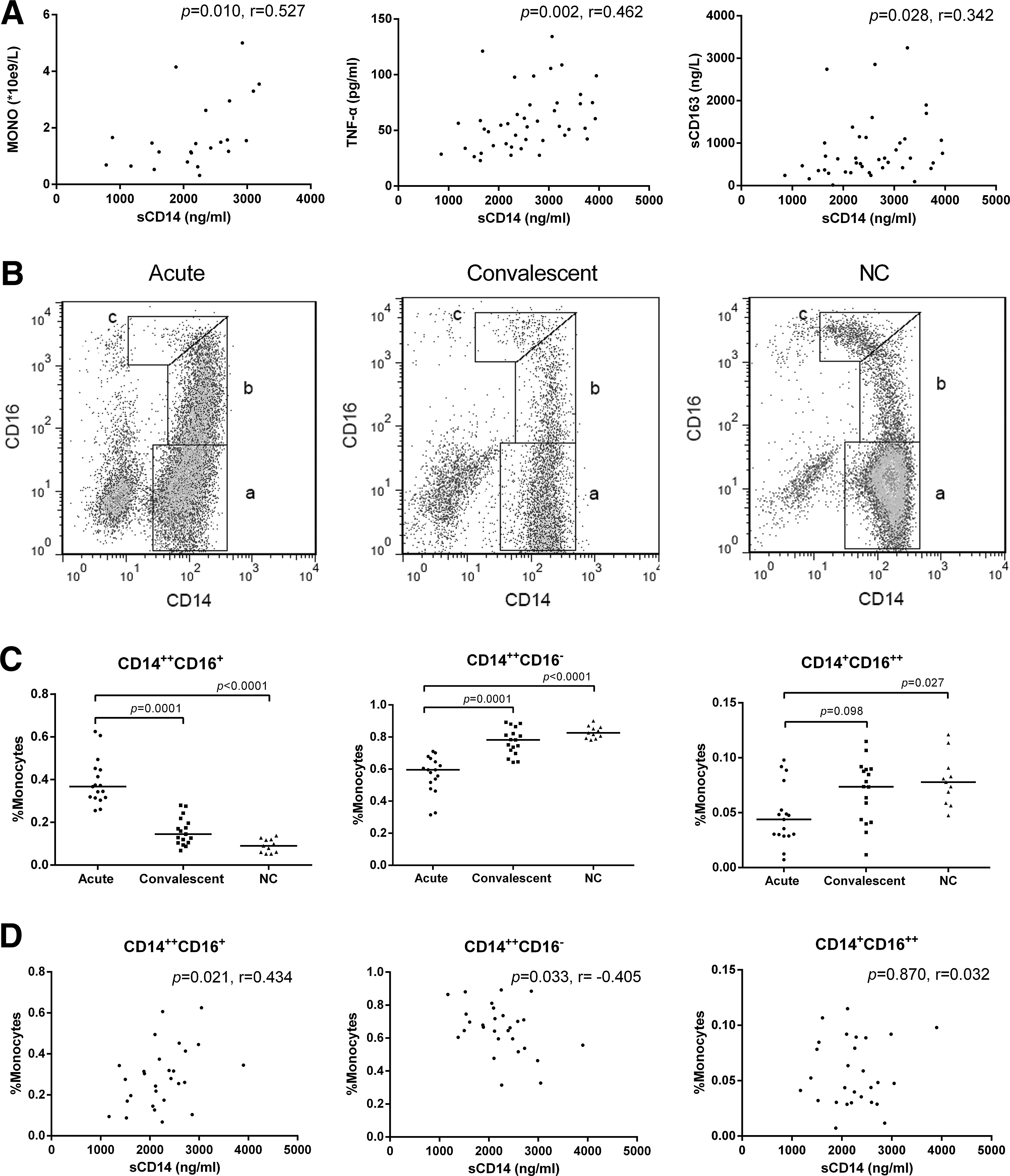
The plasma sCD14 level correlation with the monocyte response status during HNTV infections.
The dynamic changes in the three monocyte subset populations and their correlations with the plasma sCD14 levels were investigated. Representative gating populations of the three blood monocyte subsets, which include the classical (CD14++CD16−), intermediate (CD14++CD16+), and nonclassical (CD14+CD16++) monocytes, are shown in Figure 2B (a, b, and c, respectively), and comparisons between the monocyte subset proportions during the acute and convalescent phases and the normal controls are indicated in Figure 2C.
The CD14++CD16+ monocyte proportion was significantly increased in the acute phase patients (median=36.7%, IQR=31.5–44.9%) compared with the proportion in the convalescent phase patients (median=14.5%, IQR=10.4–20.7%, p=0.0001) and normal controls (median=9.0%, IQR=6.0–12.0%, p<0.0001, Fig. 2C left). In contrast, the CD14++CD16− and CD14+CD16++ monocyte proportions were decreased in the acute phase patients (median=59.5%, IQR=49.7–65.6% and median=4.4%, IQR=2.9–6.6%) compared with the proportions in the convalescent phase patients (median=78.2%, IQR=70.8–84.3%, p=0.0001 and median=7.4%, IQR=4.2–9.0%, p>0.05) and the normal controls (median=82.6%, IQR=79.2–86.1%, p<0.0001 and median=7.8%, IQR=5.9–9.1%, p=0.027) (Fig. 2C middle and right). No significant differences between the three monocyte populations were observed between the convalescent phase patients and the normal controls (Fig. 2C).
A correlation analysis of the sCD14 levels and the three monocyte subset proportions showed that the plasma sCD14 level positively correlated with the CD14++CD16+ monocyte proportion (r=0.434, p=0.021, Fig. 2D left), negatively correlated with the CD14++CD16− monocyte proportion (r = −0.405, p=0.033, Fig. 2D middle), and did not correlate with the CD14+CD16++ monocyte proportion (p>0.05, Fig. 2D right).
Discussion
Our previous studies demonstrated that both the CD4+ (21) and CD8+ (20,29) T-cell responses to HTNV infection in humans are associated with immunoprotection. However, there are few reports regarding the role of monocyte responses to HTNV infection.
In this study, we demonstrated for the first time that the plasma sCD14 levels were substantially elevated in HFRS patients. Our results showed that elevated plasma sCD14 levels positively correlated with increased monocyte counts as well as increased TNF-α and sCD163 levels in HFRS patients. Since monocyte activation promotes sCD14 production (4), TNF-α and sCD163 have been identified as sensitive markers for monocyte activation (2,32), the results indicated a correlation between increased plasma sCD14 levels and monocyte expansion and activation during HTNV infection. Additionally, blocking matrix metalloproteinase-9 (MMP-9) expression significantly reduced sCD14 concentrations (12). As CD14++CD16+ monocytes express high MMP-9 levels (6), this monocyte subset may predominantly contribute to the observed elevated plasma sCD14 levels.
To better clarify the correlation between the sCD14 levels and the monocyte subsets in HFRS, we further analyzed the dynamic changes of three monocyte subsets in HFRS patients. A dramatic expansion of the CD14++CD16+ monocyte proportion was observed, which resulted in a decrease in the CD14++CD16− monocyte and CD14+CD16++ monocyte proportions. Noticeably, the cell numbers of all the three monocyte subsets in the HFRS patients increased to different extents, as calculated by the monocyte counts and their proportions (data not shown).
It is obvious that the decline of CD14++CD16− monocyte and CD14+CD16++ monocyte proportions was due to the dramatic expansion of the CD14++CD16+ monocytes. The plasma sCD14 levels positively correlated with the CD14++CD16+ monocyte proportion, indicating that the elevated plasma sCD14 levels may be attributed to the CD14++CD16+ monocyte expansion. CD14++CD16− monocytes can differentiate into CD14++CD16+ monocytes under some conditions (22,31), and activated monocytes can biochemically alter their mCD14 expression levels and promote liberation of sCD14 (4,8).
Therefore, the CD14++CD16+ monocytes are likely to be an activated form of the CD14++CD16− monocytes that express mCD16 and shed or directly secrete CD14 into circulation at the same time after uptake of virion or following other stimuli. If this is the case, then the process probably contributes to the increased observed CD14++CD16+ monocyte proportion with the corresponding reduction in the CD14++CD16− monocyte proportion during HTNV infection. The precise mechanism by which the increased plasma sCD14 levels and CD14++CD16+ monocyte subset proportion occurred needs to be investigated further in the future.
sCD14 benefits the gram-negative bacteria-infected host by neutralizing the biological activity of LPS based upon its ability to inhibit LPS/lipopolysaccharide binding-protein binding to mCD14 (13). Additionally, functional epitope mapping of sCD14 by phage display suggests a possible role of sCD14 epitopes as natural antimicrobial peptides (1). The correlation of plasma sCD14 levels with the WBC implied a potential relationship between sCD14 and leukocytes. In vitro, sCD14 binds directly to B cells to induce their growth and differentiation, as well as promote their survival. Additionally, sCD14 is involved in IgM homeostasis (9,26). This indicates that sCD14 plays a potential role in B-cell activation before helper T cells are involved. CD14++CD16+ monocytes seem to be predisposed to antigen presentation and processing and show the highest capacity to induce CD4+ T-cell proliferation (31). In this regard, both the increased plasma sCD14 and CD14++CD16+ monocyte levels in HFRS patients may contribute to early adaptive immune responses to HTNV infection.
TLR7 and TLR8 mediate ssRNA recognition in macrophages and activate them to produce interferon (IFN)-γ, IL-6, and TNF-α (10). Interestingly, Baumann et al. and Lee et al. showed that sCD14 can bind to both ssRNA and dsRNA and promote their selective uptake as coreceptors for endosomal TLR7 and TLR9 and lysosomal TLR3 activation (3,18). Therefore, sustained high sCD14 and expanded CD14++CD16+ monocyte levels in HFRS may play a TLR-dependent antiviral role. Additionally, Jiang et al. demonstrated that HTNV infection of endothelial vein cells (mCD14-negative cells) induces enhanced production of IFN-β, IL-6, and TNF-α through TLR4 expression upregulation (11). As a TLR4 coreceptor (13), sCD14 may contribute to this response.
sCD14 can trigger IL-6 and IL8 secretion in astrocytes and microglial cells as a direct TLR2 agonist (5) and it can induce receptor clustering for monocyte signaling through binding to TLR2-derived peptides (23). CD14++CD16+ monocytes show the highest TLR2 expression (22); thus, the cross talk between sCD14 and CD14++CD16+ monocytes may act as a crucial pathophysiological mechanism that links inflammation.
The elevated plasma sCD14 levels positively correlated with increased BUN levels, which reflects kidney dysfunction to some extent, and with increased plasma sCD163, which is associated with severe acute kidney injury in HFRS (32). In parallel, CD14++CD16+ monocytes express high angiotensin-converting enzyme and reactive oxygen species levels, which subsequently enhance local oxidative stress that can cause renal injury (27,31). Therefore, the expansion of plasma sCD14 and CD14++CD16+ monocytes that are induced by HTNV infection may play a role in renal injury during HFRS.
HFRS patients have increased platelet activation (7), and blood monocytes interact with activated platelets and ingest them to suppress their own apoptosis (17). This may contribute to thrombocytopenia and sustained CD14++CD16+ monocyte levels. Our study indicated that PLT was associated with CD14++CD16+ monocytes (data not shown) and plasma sCD14 levels; thus, there may be a mechanism linking platelets, CD14++CD16+ monocytes, and sCD14 levels, which needs further exploration.
In conclusion, elevated plasma sCD14 levels associated with the monocyte response status and sCD14 may cooperate with CD14++CD16+ monocytes to participate in inflammatory immune responses during HTNV infection. These results may provide insights into the understanding of the antiviral protection and pathogenesis of HFRS through the monocyte response to HTNV infection.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (No. 81401297) and the National Basic Research Program of China (No. 2012CB518905).
Author Disclosure Statement
No competing financial interests exist.