
Abstract
Chronic hepatitis B virus (HBV) infection causes approximately one-third of all the cases of liver cirrhosis and more than three-quarters of hepatocellular carcinoma (HCC) worldwide. There are eight different genotypes (A–H) of HBV, among which B and C are the major types of HBV in China. There is a positive correlation between viral load and level of viral splicing variants and the high risk of HCC. The aim of this study was to investigate the splicing variants of HBV circulating in HCC patients. Twenty-four carcinoma and adjacent liver tissues collected from HCC patients were studied. Using reverse transcription-polymerase chain reaction (RT-PCR) and sequencing, we identified a new type of natural splice variant with nucleotides 2448-489 and 910-2120 deleted, and we named it ASN. We also found that a higher viral load and splicing variant level existed in liver carcinoma tissues compared to paracarcinoma tissues. In the investigation of our splicing variant, we found its enhancing effect on HBV replication in vitro. Although splicing variants are not essential for the replication of HBV, they may have an important influence.
Introduction
L
HBV has a 3.2 kb genome, which is partially a double-stranded and partially a relaxed circular genome. All viral RNA transcripts are transcribed by polymerase II from cccDNA, and these RNA transcripts can be classified as either the unspliced or spliced pattern. The unspliced RNAs consist of the pregenomic RNA, the precore mRNA and the subgenomic RNA (1). The pregenomic RNA undergoes alternative splicing, and the reversely transcribed defective genomes can be packaged and released (16). When the viruses mature, they get out of the cell by way of budding or exocytosis. Although the virus itself does not cause cytopathy, the immune response to it leads to cell lesion and expedite disease progression. Till date, 14 different types of spliced defective HBV genomes have been identified and isolated from the sera and liver tissues in HBV-infected patients (2,8,14). Different splicing variants have different introns removed, and the types of splicing variants vary among different HBV genotypes. As for the HBV patients in Asia, especially in China, HBV type B and C are dominant, but facts of their splicing variants remain to be revealed. HBV genotypes have been recognized to influence the course of liver disease (20). It has been reported that the high viral load and HBV genotypes B and C are associated with the high risk of HCC, and the level of HBV splicing increases before the diagnosis of HCC (3,4). Although these splicing variants are not essential for the replication of HBV, they may have an important influence on HBV replication and viral persistence (18,19,21). A protein HBSP encoded by a splicing variant has been reported to be associated with viral replication and liver fibrosis (23) and also may induce cell apoptosis without a cell-cycle block (22). P-S fusion protein also encoded by a splicing variant is involved in viral maturation (11). However, the role played by these splicing variants remains to be clarified.
In this study, we genotyped HBV DNA samples and found a new type of natural splicing variant ASN, which contains 740 base pairs with 2448-489 and 910-2120 removed compared to the wild type. We quantitated viral load and the level of splicing variant in liver carcinoma tissues and paracarcinoma tissues. Besides, we elucidated in vitro the enhancing effect of splicing variants on the replication of HBV.
Materials and Methods
Patients and DNA and RNA extractions
Twenty-four patients admitted into the East Hepatobiliary Hospital in Shanghai with the diagnoses confirmed based on clinical manifestation and pathological and serological examinations were included in this study. The cancer classification assessment was based on the NCCN (National Comprehensive Cancer Network) guideline. Twenty-four pairs of HCC and adjacent liver tissue samples were excised during surgical operation with the consent of the patients following the Chinese State Ethic Regulation. DNA was extracted by the DNA Extraction Kit (Tiangen, Inc.). RNA was extracted with Total RNA Extraction Kit (SLNco, Cinoasia). Reverse transcription was performed using the ReverTra Ace® qPCR RT Kit (FSQ-101; Toyobo, Inc.) following the manufacturer's protocol.
Genotyping
A pair of primers (P1 and P2) was designed for S gene within the conserved region to identify the genotype of HBV (Table 1). Polymerase chain reactions (PCRs) were performed in a 25 μL volume containing 12.5 μL 2× Taq PCR Master Mix (Tiangen, Inc.), 9.5 μL ddH2O, 1 μL P1 and 1 μL P2 (10 μM), and 1 μL DNA template. The mixtures were amplified by 30 cycles with denaturation at 94°C for 3 min, annealing at 55°C for 30 sec, elongation at 72°C for 40 sec. PCR products were analyzed by 1.5% agarose gel electrophoresis and stained with ethidium bromide. PCR amplified DNA was further sequenced by the Sanger method (ABI 3730xL). Sequences of PCR products were blasted and compared using GenBank for confirming their genotypes (7,13,15,25).
HBV, hepatitis B virus.
Identification of splicing variants
To identify the splicing variants, two pairs of primers P3 and P6 and P4 and P5 were applied to amplify the cDNA of HBV (Table 1). For each cDNA sample, PCRs were performed with these two pairs of primers using the same PCR protocol as for genotyping. After sequencing the PCR products, the sequences were blasted and compared using the GenBank database.
Quantitative polymerase chain reaction for quantification of viral load and splicing variant level
HBV viral load was quantified using the diagnostic kit for quantification of HBV DNA (Kehua, Inc.), with a linear range of 500–108 IU/mL. The quantitative real-time PCRs were performed in a 20 μL reaction volume in triplicate following the manufacturer's protocol.
To find whether these 48 cDNA samples contain splicing variants, two pairs of primers P4 and P5 and P3 and P6 were used to amplify these samples. The levels of HBV splicing variants of the cDNA samples that contained splicing variants were relatively quantified by SYBR GREEN quantitative real-time PCR (Toyobo, Inc.) compared to internal control glyceraldehyde-3-phosphate dehydrogenase (GAPDH). PCRs were performed in triplicate in a 20 μL reaction volume containing 10 μL 2× SuperReal PreMix plus, 6 μL ddH2O, 0.8 μL P3, and 0.8 μL P6 (10 μM) or primers for internal reference GAPDH, 0.4 μL 50× ROX reference dye, and 2 μL cDNA template. Control was set by the cDNA sample, which contained the least level of splicing variants. Results were calculated as times of splicing variant DNA in the samples compared with the control by 2−ΔΔCt.
Plasmid construction
Full-length DNA of the splicing variant ASN and the most common splicing variant AS1 were cloned into pcDNA3.1 plasmid, respectively, by NovoRec PCR step directional cloning kit (SinoBio, Inc.). The reaction system contained a 50 ng vector, 66.5 ng DNA fragment, 2 μL 10× buffer, and 1 μL recombinase and ddH2O was added up to a total 20 μL. Reaction mixture was incubated at 37°C in water for 30 min and transformed into DH5α, and positive clones were further selected. The pcDNA3.1 plasmid containing the HBV full length was kindly provided by Professor Zhenghong Yuan (Biomedical Research Institute of Fudan University).
Cell cultures and transfection
The HepG2215 cells (HepG2 cell transfected with HBV DNA that can stably produce HBV virions) and HepG2 cells without intrinsic splicing variants were cultivated in Dulbecco's modified Eagle's medium (Gibco) with 10% fetal calf serum at 37°C in a 5% CO2 incubator. The three plasmids containing the most common splicing variant AS1, the new splicing variant ASN, and the full length of HBV1.3 were transfected into the HepG2215 cells and cotransfected into HepG2 cells with the full-length HBV1.3 plasmid, respectively. Different ratios of the three plasmid DNAs were used (Fig. 4A–C). To make sure that the total amount of plasmid DNA used for each transfection was identical, the empty pcDNA3.1 plasmid was used to adjust. HBV1.3 plasmid was used as a positive control.
Assay for HBV replicative efficiency
Three days after transfecting AS1, ASN, and HBV1.3 plasmids into HepG2215 and HepG2 cells, cells were collected and the cellular DNA was extracted by the DNA Extraction Kit (Tiangen, Inc.). The replication efficiencies of HBV were examined by the real-time PCR diagnostic kit (Kehua, Inc.). Each sample was assayed in two separate experiments at least twice and in duplicate. The quantitative polymerase chain reaction (qPCR) protocol was as mentioned above.
Detection of HBsAg and HBeAg
Three days after transfecting AS1 and ASN plasmids into HepG2215 and HepG2 cells, the cell culture supernatants were collected. Different ratios of plasmid DNAs were used (Fig. 5A, B). The levels of HBsAg and HBeAg in the supernatants were measured by the ELISA kit (Kehua, Inc.) following the manufacturer's protocol.
Statistical analysis
Statistical analysis was performed using GraphPad Prism5. The data were evaluated using the F-test and t-test. p-Value < 0.05 was considered to be a statistically significant difference.
Results
HBV genotypes of HCC patients
Using the specific primer, we genotyped 48 HBV DNA samples, among which 26 samples were HBV B type at the rate of 54.2% and 22 samples were HBV C type at the rate of 45.8%. These results are in accordance with the previous studies that B and C are the most common genotypes in the mainland of China.
Identification of a new splicing variant ASN
Among 48 cDNA samples, we found two different types of splicing variants in 14 samples (Table 2). One was the most common splicing variant named as AS1 with nucleotides 2448-489 deleted, while the other was a new splicing variant that has not been reported yet (Figs. 1 and 2). We named it as ASN. The transcript of the new splicing variant is double spliced, one donor site is at nucleotide position 910 and the corresponding acceptor site is at nucleotide position 2120. The second donor site is at nucleotide position 2448, and the corresponding acceptor site is at nucleotide position 489. The full length of ASN genome is 740 base pairs. The deleted parts correspond to the regions that encode part of the C, S, and P proteins. Besides, the whole transcript region encoding X protein was deleted.
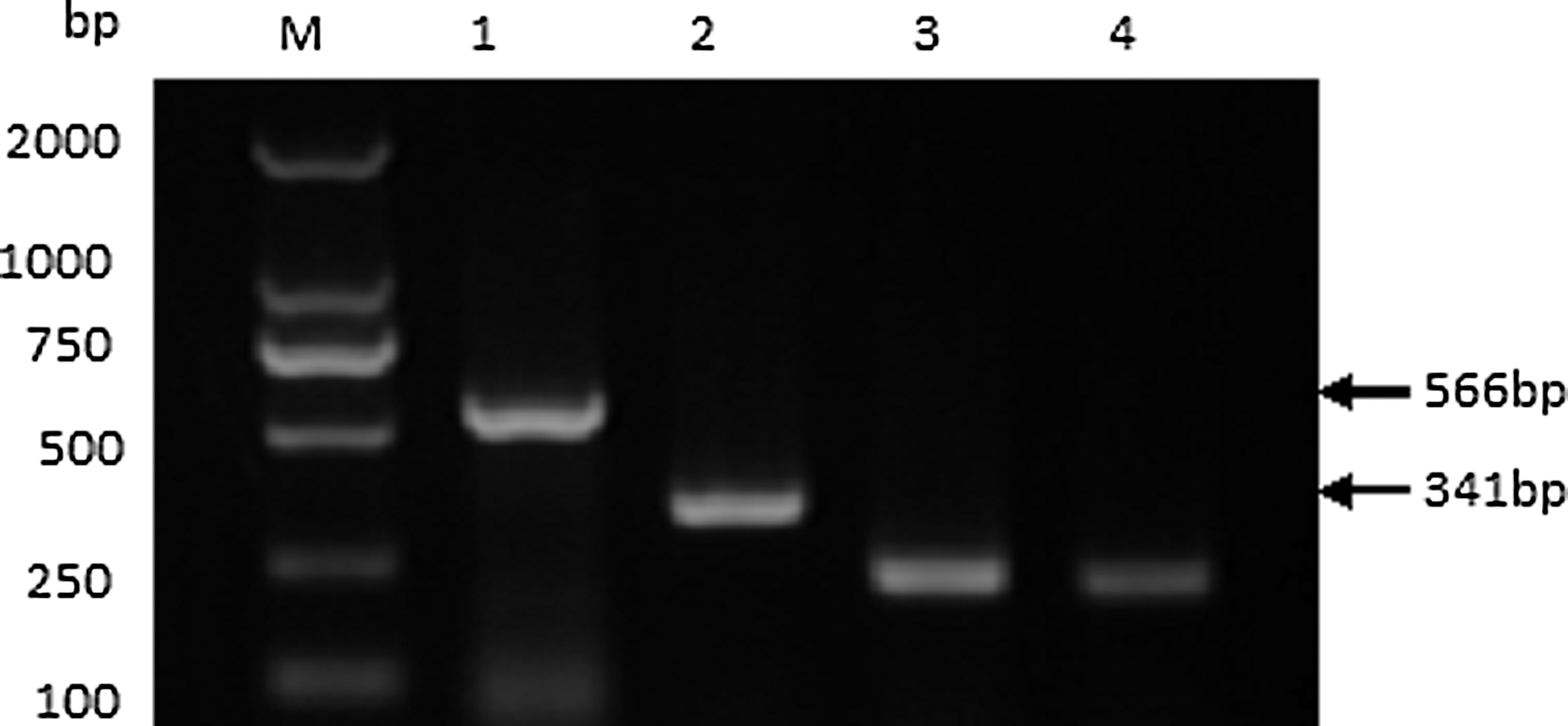
Electrophoretic analysis of polymerase chain reaction (PCR) products from the specific cDNA sample that contained the new splicing variant ASN. Lane M contains markers 2000. Lane 1 is the product using the primer P4 and P5, lane 2 is the product using the primer P3 and P6, lane 3 is the product of inner control GAPDH using the primer GF and GR, and lane 4 is the product of positive control (the sample contained virion definitely). The sizes of marker DNAs and PCR products are indicated. GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
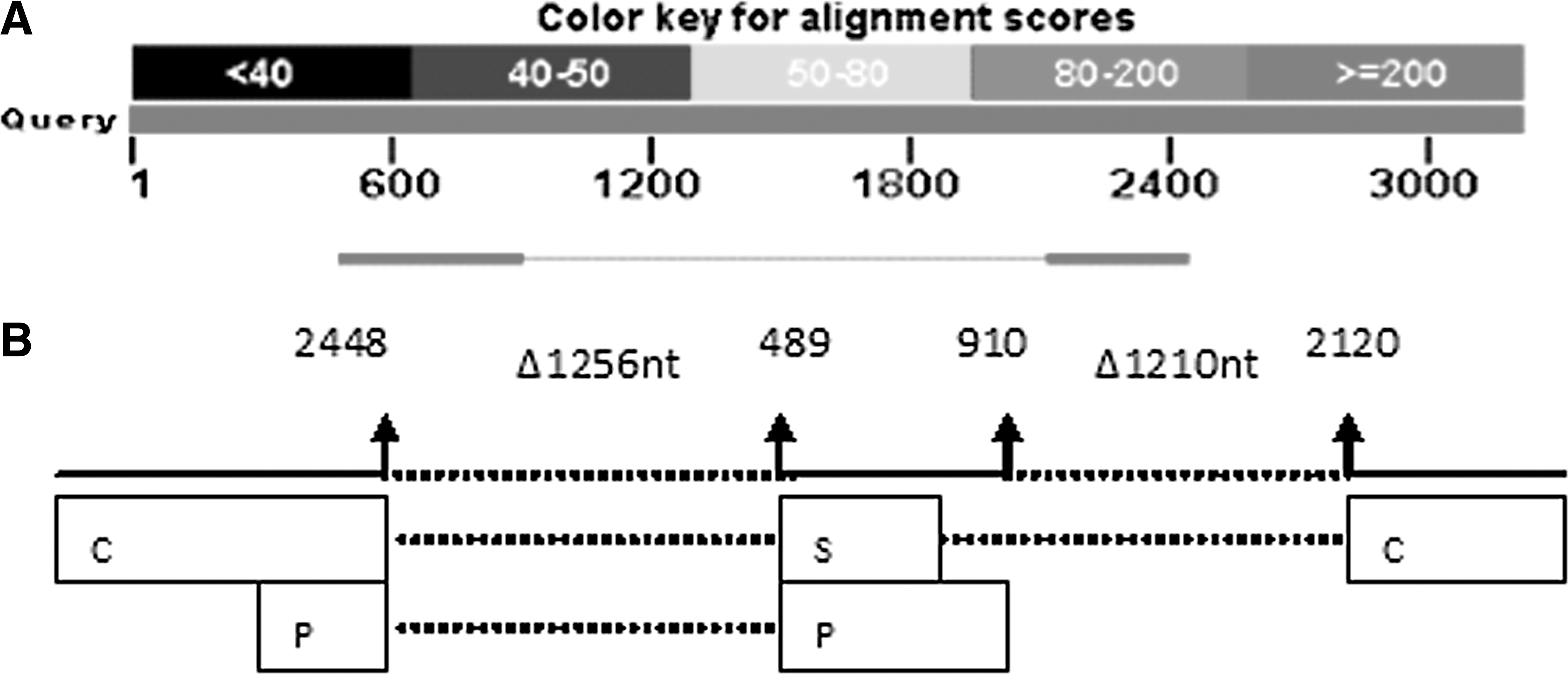
The genome of ASN.
Viral load and the level of splicing variant
By comparing the viral load in HCC tissues and paracarcinoma tissues, we found that viral load was significantly higher in liver carcinoma tissues (p = 0.0081). We also compared the difference of viral load between the HBV genotype B and genotype C, but the result showed no significant difference between HBV genotype B and genotype C (p = 0.7079) (Fig. 3A, B).
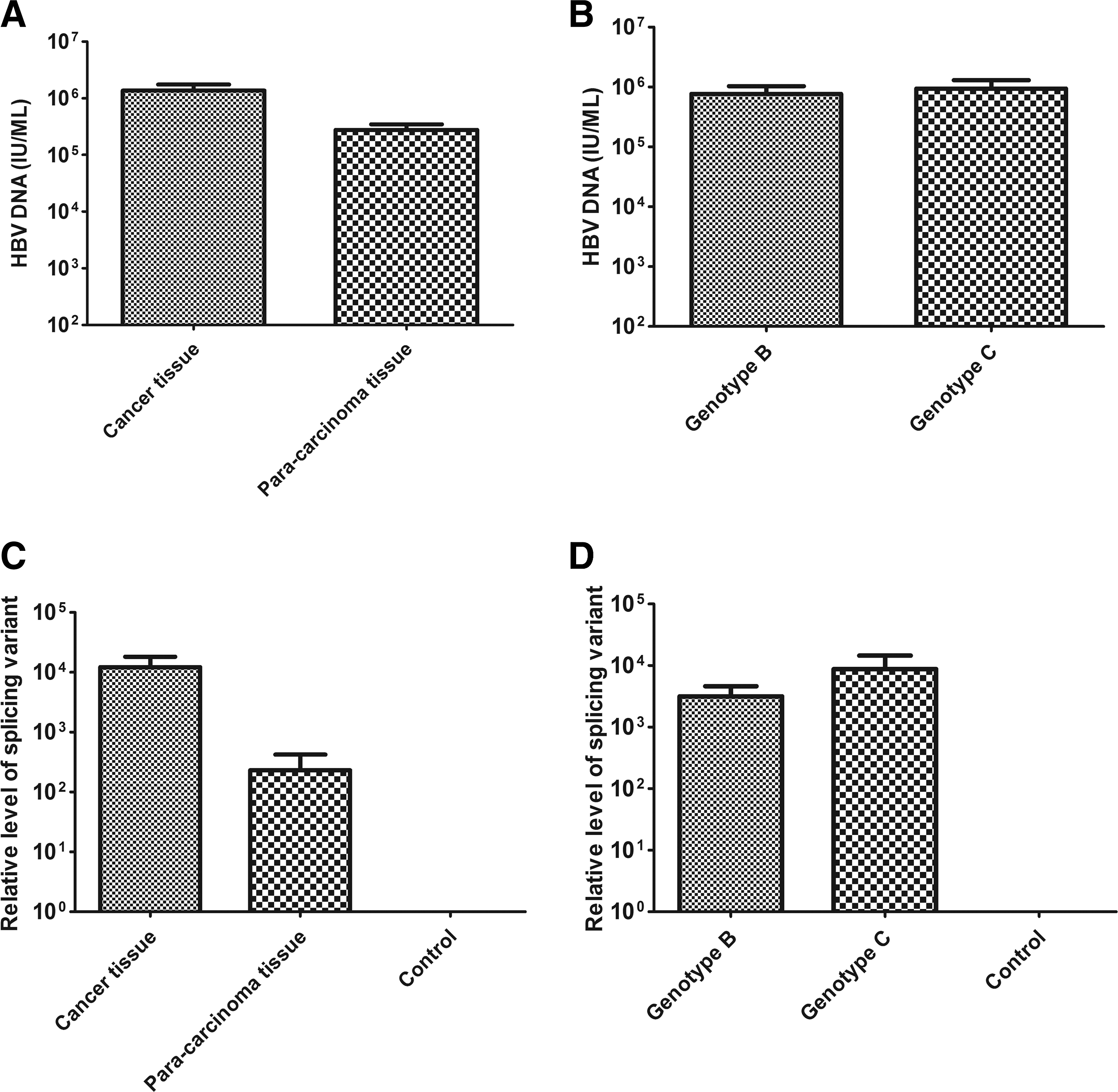
Comparison of viral load in DNA samples and the level of splicing variant DNA in cDNA samples.
Fourteen samples contained two types of splicing variants ASN and AS1 that share common deleted splicing sequences from 2448 to 489. Comparing the level of splicing variant transcripts between carcinoma and adjacent liver tissues, we found that the level of splicing variant transcripts in liver carcinoma tissues was significantly higher than that in adjacent tissues (p = 0.0498). We also compared the level of splicing variant transcripts between HBV genotype B and C. There was no significant difference between HBV genotype B and C (p = 0.2360) (Fig. 3C, D).
Cotransfection of ASN/AS1 with the full-length HBV in HepG2 cells
Both splicing variants exerted an enhancing effect on the replication of HBV. Increasing the amount of ASN/AS1, the replication enhancing effect raised correspondingly. Compared to the results of transfecting HBV1.3, the replication enhancing effect of AS1 was much stronger, while the new splicing variant ASN was relatively weaker (Fig. 4A–C). The results of transfection of HepG2215 cells with ASN, AS1, and the full-length HBV are in accordance with those for HepG2 (data not shown).
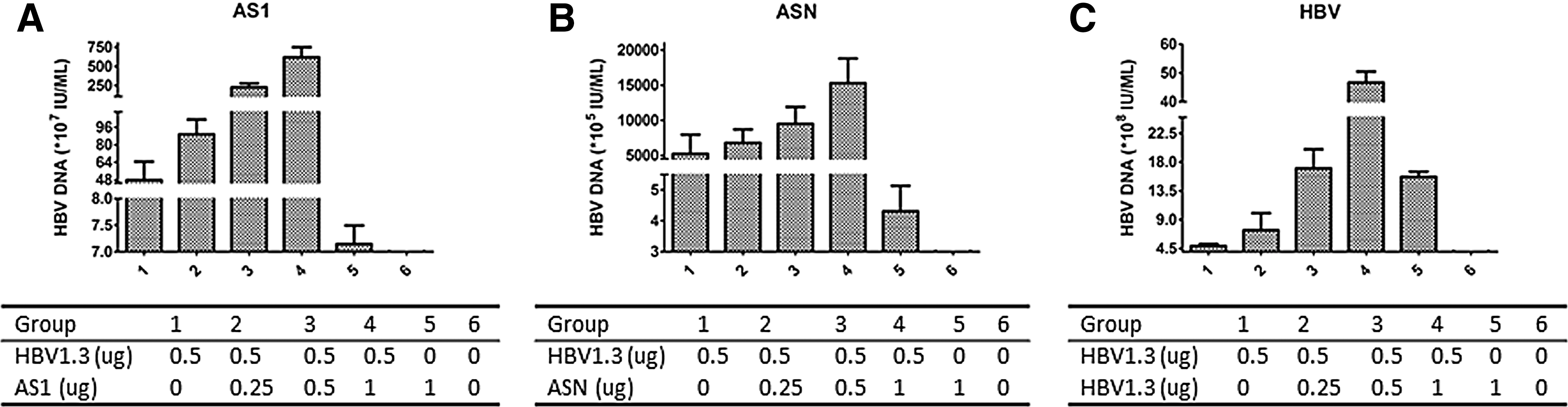
Comparison of replicative competencies of HepG2 cells transfected with different plasmids detected by real-time PCR.
HBeAg and HBsAg levels in transfected cell supernatants
The expression levels of HBsAg and HBeAg were both increased with incremental ASN/AS1, but compared with AS1, ASN exerted a relatively lesser effect (Fig. 5A, B), which was in accordance with the enhanced replication of ASN/AS1. The effect of transfection of ASN and AS1 into HepG2215 cells was similar to what was shown in HepG2 (data not shown).
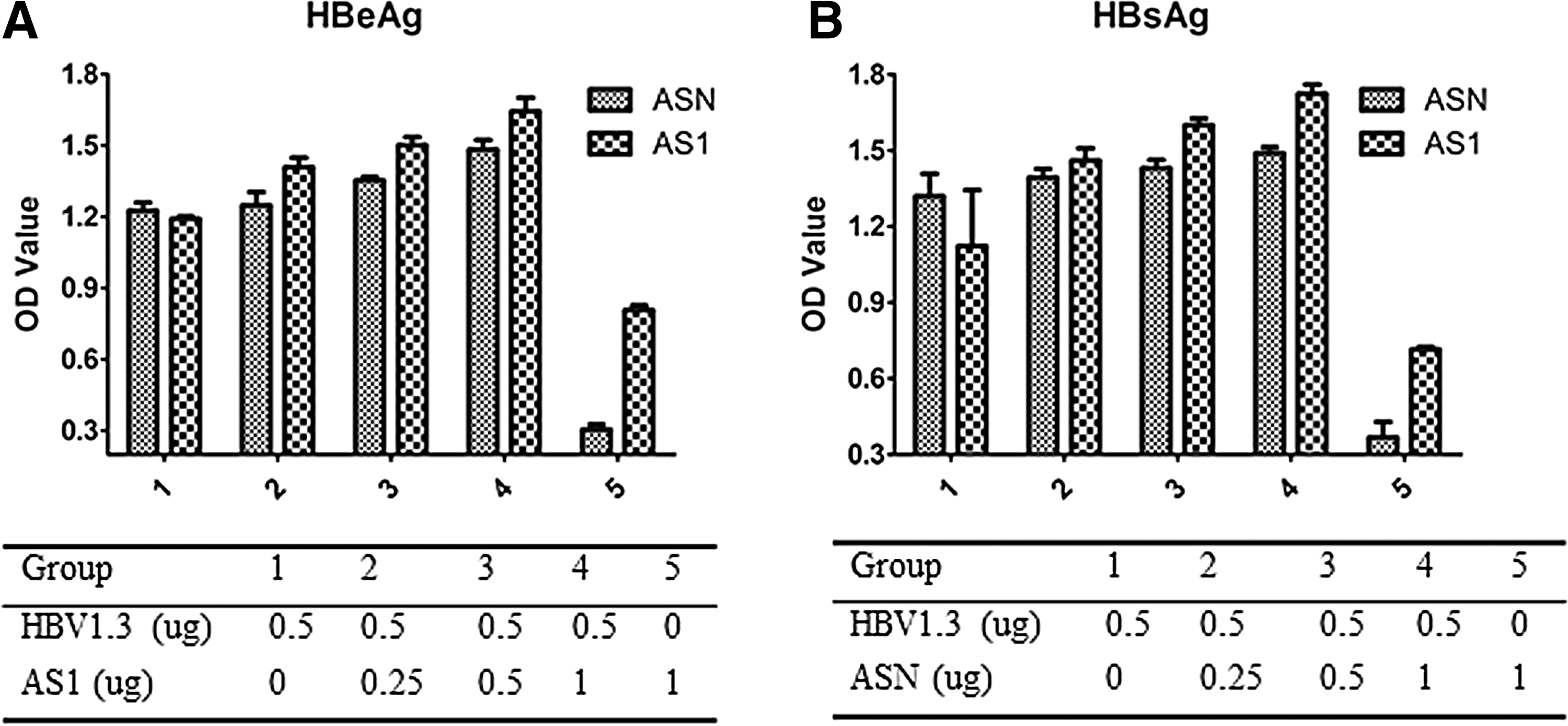
HBeAg and HBsAg levels from supernatants of transfected HepG2 cells by ELISA.
Discussion
HBV is a critical cause of HCC worldwide, especially in developing countries. Although the pathogenesis of HBV-related HCC still remains unclear, there is strong evidence showing that HBV itself is a direct cause of HCC (6).
HBV is known to encode just four main proteins, but it remains persistent and chronic in the hepatocyte. It has been reported that HBV produces several different kinds of splicing variants, which may exert different functions on viral persistence. In this study, we addressed the alternative splicing of HBV and the difference of splicing variant levels in carcinoma and adjacent liver tissues. It has been reported that there are 14 types of splicing variants of HBV transcript, and the splicing is regulated by the splicing regulatory network (5,24). The splicing sites of HBV genome are not random, rather, there are five common donor splicing sites at the nucleotide position 2067, 2447, 2471, 2985, and 2087 and five common acceptor splicing sites at nucleotide position 489, 2350, 2236, 2902, and 282. Among the splicing variants discovered, the AS1 (with nucleotide position 2447-489 removed) is the most common one (1,5). In this study, we have found a new splicing variant that is double spliced, with nucleotides 2447-489 and 910-2120 deleted. In general, the splicing variants tend to keep the completeness of X gene (9), but interestingly, the newly identified splicing variant ASN discards the whole X gene and part of S gene, C gene, and P gene. This suggests that there might be more splicing variants that have not been discovered yet.
The level of splicing variants in liver carcinoma tissues is significantly higher than that in adjacent tissues. We postulate that the higher level of splicing variants might relate to the severity of the disease, but the mechanism of pathogenesis needs further study, and the level of splicing variants may be used as a biomarker to predict prognosis of the disease.
We have done fundamental research in vitro about the potential function of these two splicing variants, AS1 and ASN. Results showed that the common splicing variant AS1 could substantially enhance the replication of HBV, but the new splicing variant ASN could not exert as strong an effect on enhancing the replication of HBV as AS1 does. This suggests that the splicing variants are involved in viral replication and persistence, but different splicing variants may have different functions. It is not clear whether all the splicing variants of HBV play a role in pathogenesis. Since the hepatocarcinoma tissue samples we have studied were limited, their effect on HBV replication and pathogenicity in vivo needs further investigation. It will be very important to understand the function and the mechanism of HBV splicing variants. By combining with clinical manifestation, alternative splicing of HBV transcript will shed light on new ways to the treatment of HBV.
Footnotes
Acknowledgment
The authors thank Professor Zhenghong Yuan for kindly providing the pcDNA3.1 plasmid containing the HBV full length.
This work was supported by the National Major Scientific and Technological Special Project (2012ZX10002006-002-003), the National High Technology Research and Development Program of China (2011AA02A114), the National Natural Science Foundation of China (31370927 and 30571650), and the Science and Technology Innovation Action Plan of Shanghai (13431900602).
Author Disclosure Statement
No competing financial interests exist.