
Abstract
Influenza is an acute and highly contagious respiratory disease. The error prone RNA polymerase and segmented nature of the influenza A virus genome allow antigenic drift and shift, respectively. Therefore, most influenza vaccines are inefficient along time and against different viral subtypes. In this study, for the first time, protection properties of a new recombinant fusion of HA2 and M2e peptides originated from influenza virus A/Brisbane/59/2007-like (H1N1) in BALB/c mice model were investigated. After immunization of the BALB/c mice, the protection property of fusion peptide was determined by a neutralizing assay test. For further study, mice were lethal challenged by the (mouse adapted, A/PR8/34 [H1N1]) and heterologous (mouse adapted, A/Brisbane/10/2007 [H3N2]) influenza virus subtypes. Then, the lung viral titers, body weight, and survival rate of the immunized mice were monitored. The results showed that immunization by the M2e-HA2 recombinant fusion peptide provides strong protection against homologous challenge and an infirm protection against heterologous. These protections against homologous and heterologous influenza A virus challenges meant the universal nature of these recombinant peptides in an immunity manner against influenza A virus. However, more studies are needed to optimize this recombinant construction, and this experiment recommends HA2-M2e fusion peptide as a universal influenza A vaccine candidate.
Introduction
T
The segmented nature of the influenza A virus genome is the cause of antigenic shift incidence. Antigenic shift could lead to pandemics by emerging new viruses possibly transferable between different species (14). Influenza virus error prone polymerase causes antigenic drift. Antigenic drift induces variation in surface glycoproteins of the virus (14). Antigenic shift and drift make conventional vaccines ineffective in influenza disease control over time through antigen mismatch between the vaccine and the circulating viral strains (15). Universal influenza vaccines hurdle these problems through inducing protection against different subtypes of influenza A virus at the same time (30). Ectodomain of matrix protein 2 (M2e), stem subunit of the hemagglutinin glycoprotein (HA2), and nucleoprotein are the most conserved proteins of the influenza A virus, which are protected against genetic drift and yet can induce effective protection against the virus and these characteristics make them proper targets for universal influenza vaccines (17,22,23).
M2e with only 23 amino acid residues is the external domain of the matrix protein2 (M2). M2e has lasted almost invariable since the first human influenza virus strain isolated in 1933 (5,12,13). It also has borne few changes among different species, for example, the typical amino acid residue sequence of M2e in human and avian cases is different only on four positions 11, 14, 16, and 20 (26). The N-terminal SLLTEVET (residues 2 to 9) is conserved in all subtypes of influenza A viruses (6). These make M2e an attractive target to produce universal vaccine against influenza A virus. The M2e induces specific IgG and stimulates T-cell responses (29). However, the mechanism of M2e-based vaccines function has not been clear (29).
Influenza virus hemagglutinin consists of two subunits, HA1 and HA2. The HA2 subunit with 221 residues has a hair spin structure composed of two antiparallel alpha-helixes (24). HA2 is smaller and more conserved than HA1. HA2 antibodies can protect mice from the challenges by viruses from different clades (2).
A recommended trend in the design of universal flu vaccines is constructing fusion peptides based on the combination of different conserved epitopes of the virus proteins (18). As a result, it could preserve universality features and improves the efficacy of the vaccine. Furthermore, this approach reduces the time and cost of manufacturing compared to processes that target single peptides (6).
Based on the above mentioned, considering well-conserved sequences of M2e and HA2 and deficiencies in immunity induction in the use of the single epitopes (M2e or HA2), it seems that the chimeric antigens of these can offer effective and universal protection against influenza A virus subtypes. Recently, we reported the design and cloning of a new structural model containing HA2 gene partial cds and M2e gene partial cds fused together by a small linker in Escherichia coli. M2e peptide is the production of a spliced mRNA that includes nucleotide 1 to 26 and 715 to 757 of 7th segment of influenza virus genome. HA2 is a 666 nucleotide fragment of 4th segment of influenza virus genome, from 1030 to 1695, which was linked together by the AEAAAKEAAAKA sequence as a small linker (Fig. 1) (1). Therefore, the purpose of this study was to survey the protection effects of M2e-HA2 fusion peptide on a mouse model. To do this, we immunized mice groups (n = 12) with the M2e-HA2 fusion peptide and 14 days after the last immunization, challenged them with homologous and heterologous mouse adapted viruses and determined induced protective effects.
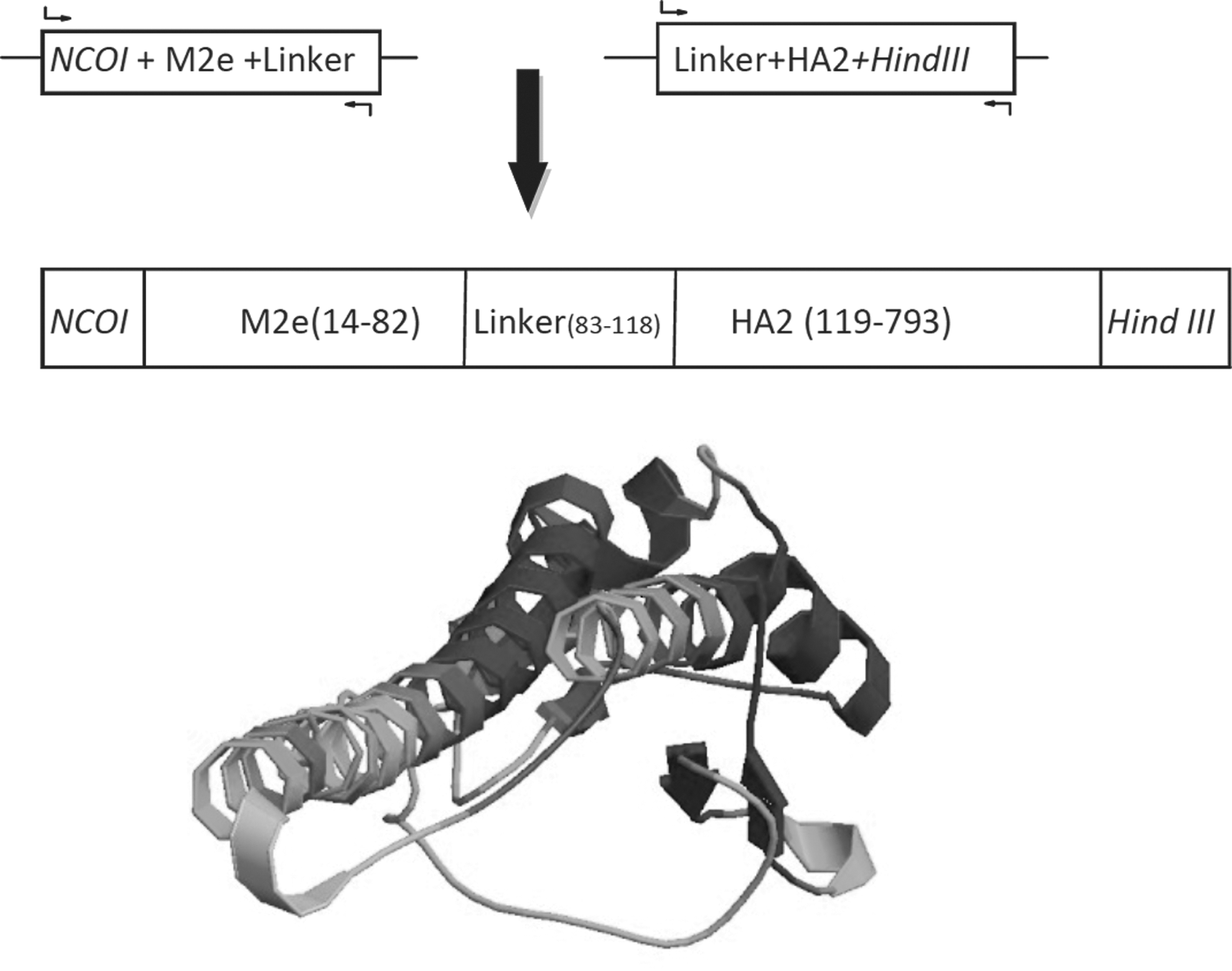
M2e-HA2 fusion peptide constructs: From top to bottom: Fusion PCR to fuse M2e and HA2 segments. The M2e-HA2 fusion nucleotide include NcoI incision site, M2e segment include nucleotide 1 to 26 and 715 to 757 of 7th segment of influenza virus genome, AEAAAKEAAAKA sequence as a small linker, HA2 is a 666 nucleotide fragment of 4th segment of influenza virus genome, from 1030 to 1695 and HindIII incision site. M2e-HA2 fusion peptide3D model depicted by
Materials and Methods
Viruses, cells, and proteins
Influenza A/Brisbane/59/07 (H1N1) was purchased from the NIBSC (United Kingdom) and mouse adapted influenza virus, A/PR8/34 (H1N1), was kindly gifted by Kheiri (Pasteur Institute, Iran). A/Brisbane/10/2007 (H3N2) mouse adapted after 10 time passages in mouse lungs, kindly gifted by Taghizadeh (Razi Institute, Iran). Fifty percent of infectious dose (EID50) titers of the viruses were determined by serial titration in fertilized eggs and calculated by the method of Reed–Muench (16). Median lethal dose (MLD50) titers of mouse adapted A/PR8/34 (H1N1) and A/Brisbane/10/2007 (H3N2) were determined separately by serial titration in groups of five BALB/c mice with serial 10-fold dilutions of virus and calculated using the statistical method of Reed and Muench (16).
SPF fertilized chicken eggs were prepared from the Razi Vaccine and Serum Research Institute. Madin–Darby canine kidney (MDCK) cells (ATCC CCL-34 was kindly gifted by Dr. Taghizadeh) were cultured as monolayers in Dulbecco's modified Eagle's medium (Gibco) containing 10% fetal bovine serum (Gibco) and incubated in a CO2 incubator at 37°c. Influenza virus A/Brisbane/59/07 (H1N1) M2e peptide was kindly gifted by Taghizadeh. Influenza virus A/Brisbane/59/07 (H1N1) HA2 peptide was bought (Sino Biological). TIV or (VAXIGRIP®) a three-valent inactivated influenza vaccine contains the following: A/California/7/2009 NYMC X-179A (A/California/7/2009 [H1N1]pdm09-like), A/Victoria/361/2011 IVR-165 (A/Victoria/361/2011 [H3N2]–like), and B/Hubei-Wujiagang/158/2009 NYMC BX-39 (B/Wisconsin/1/2010-like) were bought from SANOFI-PASTEUR.
HA2-M2e fusion protein production
As described previously (1) in brief, total RNA of the influenza A/Brisbane/59/07 (H1N1) was extracted, and the virus cDNA was amplified using universal primers (10) and RT-PCR (3). PCR amplification of M2e and HA2 genes was done by designed M2e and HA2 gene segment primers on the virus cDNA as template. After confirmation of the PCR products by sequence analysis, fusion PCR was carried out for the M2e-HA2 chimeric fusion gene construction (20). M2e, HA2, and M2e-HA2 chimeric fusion genes were cloned separately in pET-22b+(Novagen). Transformed E. coli BL21 cells were grown and induced by IPTG. After 8 h, the cells were sonicated and the inclusion bodies were separated from cell lysate. By using the Ni-NTA column (Qiagen), the proteins were purified from the inclusion body solutions. The proteins concentration was measured using the Bradford assay.
Animal experiments
Animals
Six- to 8-week-old female Balb/c mice were obtained from the Razi Vaccine and Serum Research Institute (Karaj, Iran) and kept in an animal facility at the same institute according to the policy of the Department of Animal Care and the European Communities Council Directive of 86/609/EEC.
Immunization of mice
Mice in five groups (HA2, M2e, HA2-M2e, TIV, and control) (n = 12) were subcutaneously immunized three times at 2-week intervals (on 0, 14, and 28 days) with 5 μg of each protein. The purified proteins were quantified by the Bradford assay (1) and emulsified with adjuvant Stimune® (Prionics, Inc.). Stimune is a water in oil adjuvant composed of defined and purified light mineral oil (
Lethal homologous and heterologous challenge
The mice groups were challenged with 50 μL of the viral suspension containing 5MLD50 mouse adapted A/PR/8/34 (H1N1) or A/Brisbane/10/2007 (H3N2) viruses 2 weeks after last immunization, by intranasal drop after light anesthesia by ketamine/xylazine (Roche). Mice body weights were measured daily. Mice that lost body weight more than 25% were humanely euthanized.
Lung viral titers
Three days after the challenges, four mice from each group were sacrificed. Mice lungs were homogenized in 3 mL Dulbecco's modified Eagle's medium (HiMedia) plus 0.1% bovine serum albumin (Life technologies). MDCK cells were cultured in 24-well plates and formed a confluent monolayer. The homogenized lungs were 10-fold serially diluted from 1:10 and inoculated on MDCK cells; six wells of every concentration were repeated. After incubating at 37°C for 2 days, the cytopathic effect was observed. The virus titer of each specimen was calculated as 50% tissue culture infection dose (TCID50) by the Reed–Muench method (16).
Neutralization assay
The neutralizing antibody titers were measured against A/PR/8/34 (H1N1) and A/Brisbane/10/2007 (H3N2) viruses in microneutralization assay as described previously (28). Serum samples were obtained 14 days after the last immunization. Briefly, 10 μl of serially diluted test sera was incubated with diluted virus for 1 h. Then, 1.5 × 104 MDCK cells were added to wells and incubated overnight. The cells were then fixed and growth of the virus was determined by ELISA with anti-influenza A Np mouse monoclonal antibody (Millipore) as the primary antibody and HRP-conjugated anti-mouse (Invitrogene) as the secondary antibody. Ten minutes after adding the substrate (o-phenylenediamine; Invitrogen), the absorbance at 490 nm was measured.
Hemagglutination inhibition assays
The sera from each group of immunized mice were diluted twofold serially with PBS (treat with receptor-destroying enzyme, Denka Seiken, Tokyo, Japan) on a 96-well microplate; virus suspension containing four hemagglutinin units was added to each well. After incubation of the plate at room temperature (RT) for 1 h, 50 μL of 0.5% (v/v) chicken red blood cells was added to each well and the plate was incubated at RT for 1 h. The hemagglutination inhibition (HI) titers were determined as the highest serum dilution that completely inhibited hemagglutination.
Statistical analysis
Statistical analysis of the mice group's data was performed using the Student's t-test with SPSS 16 software by using mean standard deviation, also using SigmaPlot software for designing the graphs.
Results
HA2-M2e fusion protein production
Western blot using the anti-His tag monoclonal antibody was done on the transformed cell lysates and showed a ≈30KD band and confirmed the expression of the pET22b(+)/M2e-HA2 in the system. After purification of the fusion peptide by Ni-NTA column (Qiagen), the peptide concentration measurement was done by the Bradford assay. The final product contained a concentration of 25 μg/mL of HA2-M2e.
Lethal homologous and heterologous challenge
Fourteen days after the challenge against H1N1 virus, the TIV group, M2e-HA2 group, and HA2 group had 83%, 66%, and 50% survived mice, respectively. At the same condition, TIV and M2e-HA2 groups showed minimum weight changes. In challenge by H1N1virus, survived mice of HA2 group showed weight increase after the 10th day. Fourteen days after the challenge against H3N2 virus, the TIV group, M2e-HA2 group, and HA2 group had 75%, 33%, and 25% survived mice, respectively. M2e groups in both cases showed the weakest results, which were a little better than the control group (Fig. 2A). In challenge against the heterologous virus, the TIV and M2e-HA2 groups showed minimum weight changes too; however, weight decrease in these cases was more a homologous challenge. The M2e and control groups showed maximum weight decrease in both homologous and heterologous challenges (Fig. 2D).
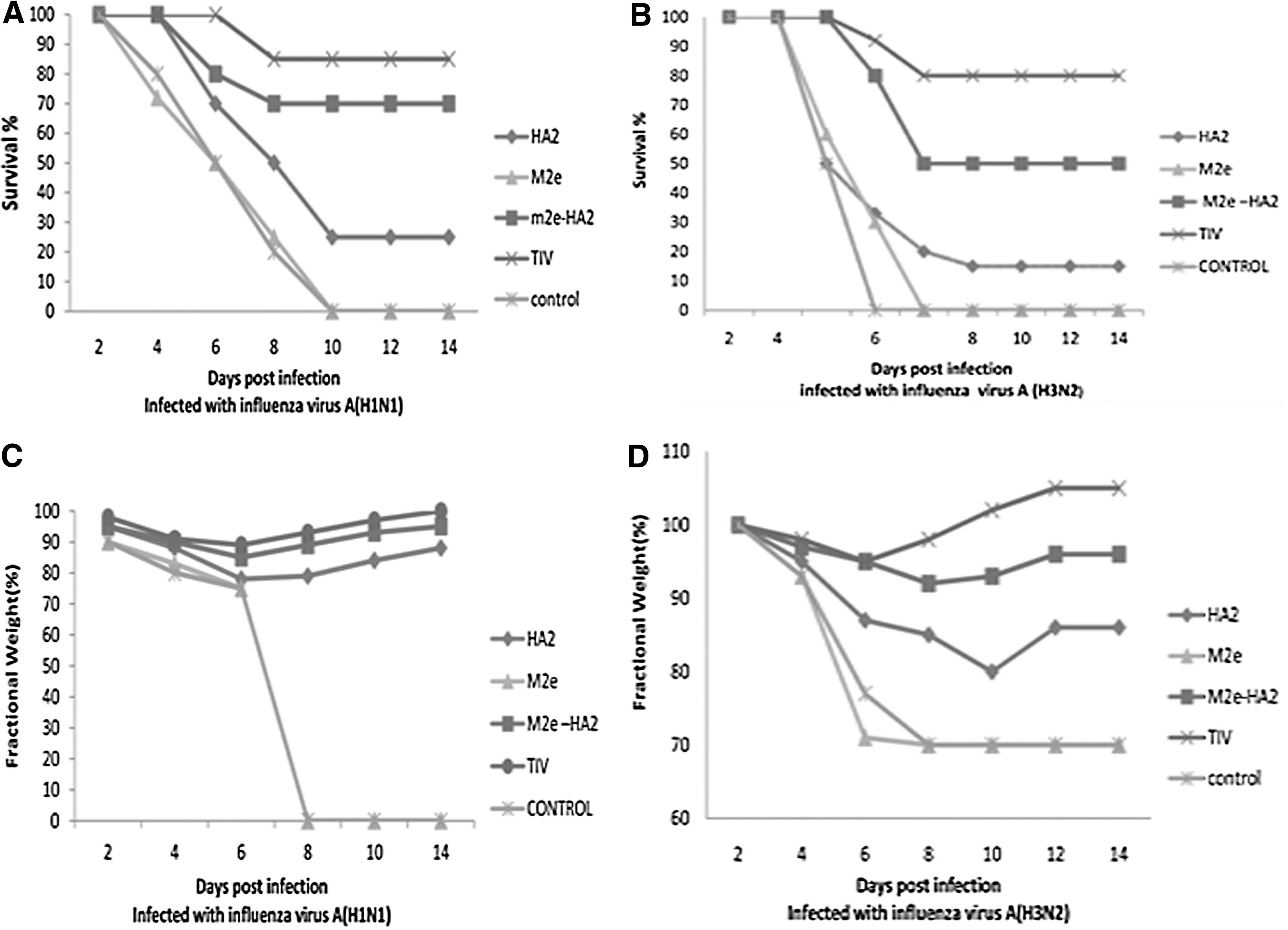
Protection of mice groups against homologous and heterologous lethal challenges:
Lung viral titers
The minimum viral titer was observed in TIV group samples. M2e-HA2 group samples showed a significantly lower viral titer than HA2 group samples (p < 0.05%). Also, the M2e group showed a significantly lower viral titer than the control group but it was significantly higher than the HA2 group (p < 0.05%) (Table 1).
Weight loss: percentage difference from prechallenge weight to the lowest weight observed after challenge.
Signs of disease: change in observed clinical signs.
Mild, ruffled fur, slowing activity, loss of body conditions; None, no change from prechallenge condition.
Neutralization assay
The TIV group, M2e-HA2 group, and HA2 group displayed 10%, 30%, and 53% infectivity, respectively. In the neutralization assay against H3N2 virus, the TIV group, M2e-HA2 group, and HA2 group revealed 28%, 55%, and 70% infectivity, respectively. The M2e group in both cases showed the weakest results that were a little better than the control group (Fig. 3).
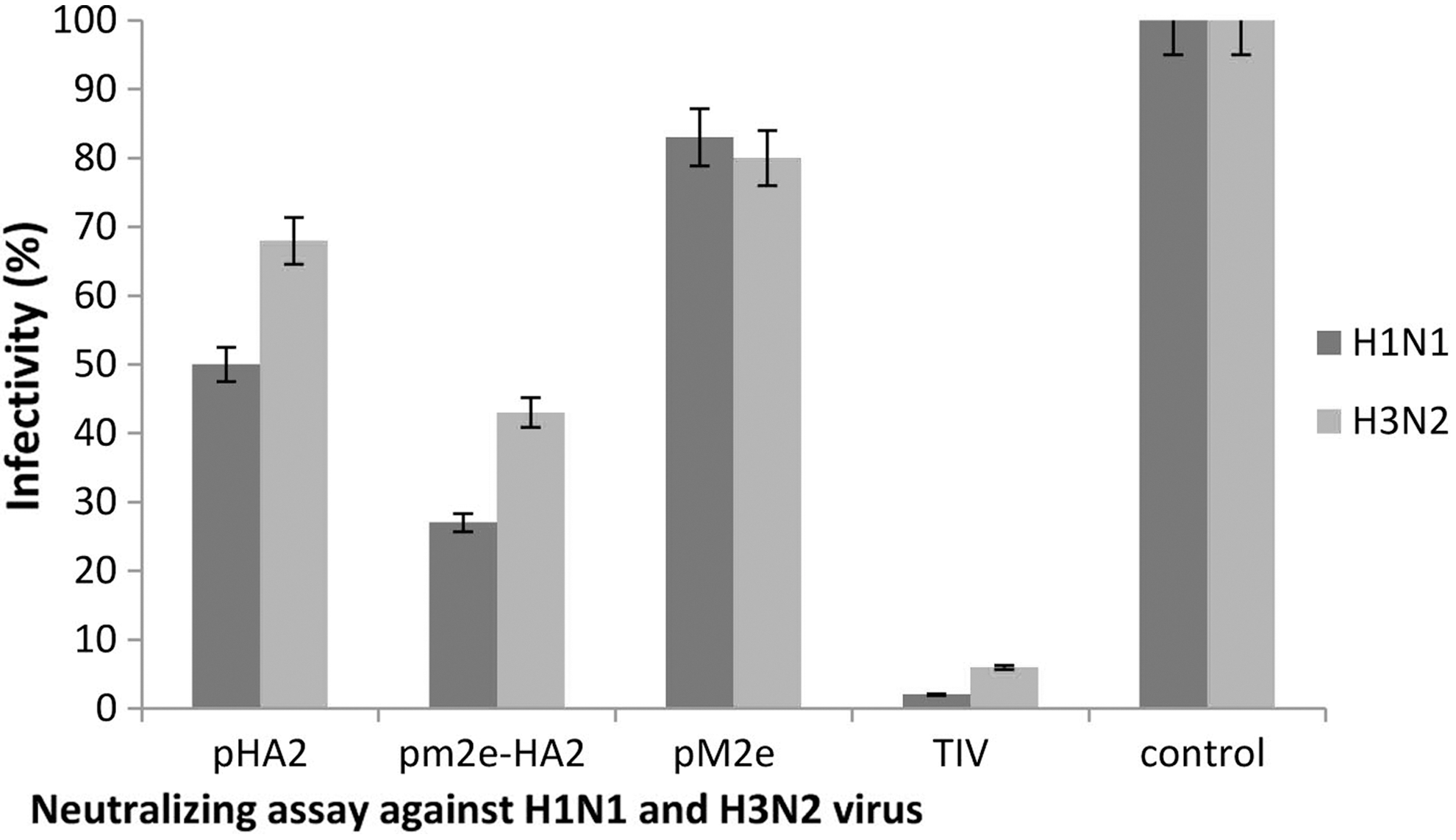
Neutralization assay against homologous (H1N1) and heterologous (H3N2) viruses: Infectivity rate of Madin–Darby canine kidney cell cultures after exposing to treated viruses. The treatment done by sera of mice groups immunized with different antigens.
HI assay
HI assay results on day 0 showed 0 for all groups, these results showed a significantly higher titer in TIV and M2e-HA2 groups on days 14, 28, and 42. However, titers for the TIV group were higher than M2e-HA2 (Fig. 4).
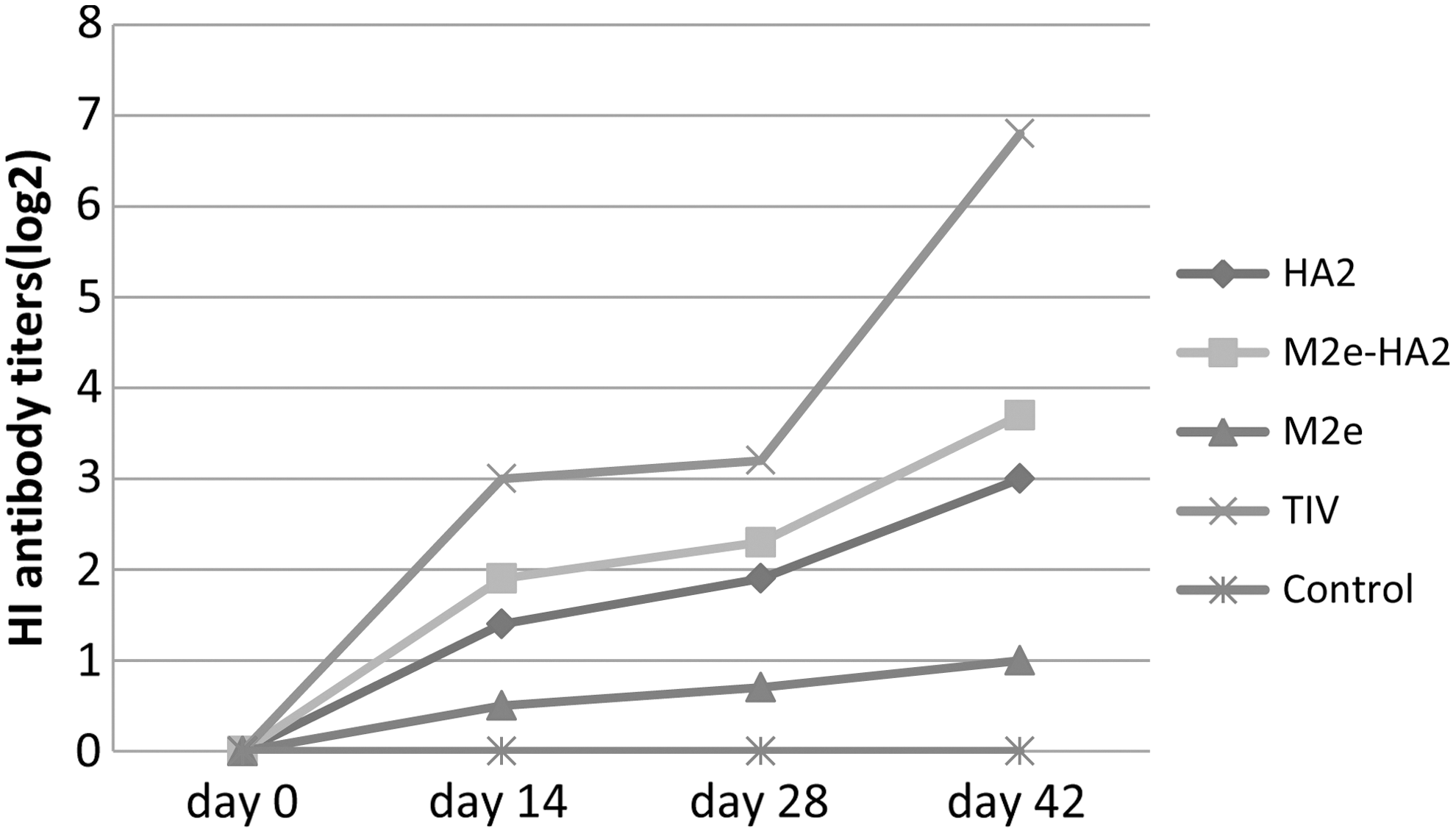
HI assay: HI titers of HA2, M2e, M2e-HA2, TIV, and control groups after each immunization on 0, 14, 28, 42 days; day 42 was time point before the challenge. HI, hemagglutination inhibition.
Discussion
Because of antigenic shift and drift in influenza A virus, there is a need for universal vaccines to protect human against all subtypes of influenza A viruses, including future pandemics. Conserved viral proteins such as M2e and HA2 are the targets for this approach (17,22,23). The M2e peptide contains 23 amino acid residues. It is highly conserved, for example, M2e human immune serum has cross reaction with almost all varieties of avian M2e (7). There are some reports about the M2e immunogenicity against influenza virus. Schotsaert et al. and Fiers et al. have demonstrated the safety and immunogenicity of M2e-based vaccine in a phase I clinical study. This vaccine is a production of VaxInnate company, it is an M2e-based influenza vaccine with a tool-like receptor-mediated immune enhancement (8,19).
Ebrahami and his colleagues reported that the use of M2e with an adjuvant can stimulate the immune system and improve the immune responses (25). HA2 subunit with 221 amino acids is a conserved peptide. The 11 first residues of HA2 N-terminal are conserved among all influenza virus subtypes (6). Bommakanti et al. expressed HA2 in E. coli (2). There are some reports about the HA2 immunogenicity against influenza virus (2,27). The HA2 molecule is larger than M2e and also is more immunogenic (18). Maybe fusions of these two peptides can increase synergetic effects on the universality and immunogenicity of both of them. In this study, we used the HA2 protein for its specific immune response and as an immune system stimulating along M2e and for the first time, we fused these two genes, expressed M2e-HA2 fusion peptide, and tried to study the protective effects induced by this fusion peptide against homologous and heterologous influenza virus subtypes.
The neutralization assay and the lung viral titer results after the immunization revealed that the M2e-HA2 fusion peptide stimulated mice immune system properly. Considering homologous challenge results, the survival percent in the M2e-HA2 group was significantly higher than the HA2 and M2e groups (Fig. 2A) and the weight loss in the M2e-HA2 group was significantly less than the HA2 and M2e groups (Fig. 2C). These results showed synergetic effects of M2e and HA2 on immunologic response. Results in heterologous challenge showed a significantly higher survival rate in the M2e-HA2 group than the HA2 and M2e groups (Fig. 2B) and less weight loss in the M2e-HA2 group than M2e and HA2 groups (Fig. 2D), which once again confirmed the synergetic effects of M2e and HA2 on immunologic response.
From another point of view, the concomitancy of protective effects of M2e-HA2 and TIV groups in heterologous challenge suggests the fusion peptide potency as a universal vaccine candidate. However, the survival rate and weight loss results in the TIV group were a little better than in the M2e-HA2 group (Fig. 2B, D), perhaps because there was split inactivated H3N2 subtype virus in TIV content. Results of lung viral titer assay after homologous and heterologous challenges showed that the induced immune responses could inhibit the virus replication. In homologous challenge, TIV and M2e-HA2 group results were stronger than HA2 and M2e groups. However, in heterologous challenge, the antibody responses had conformity with the homologous challenge but the immune response could not inhibit the viral replication in the cases of M2e-HA2, HA2, and M2e groups.
Neutralization assay results showed that alone M2e could not neutralize the viruses that reported previously (9). Although HA2 group showed neutralizing properties but stronger neutralizing properties in the M2e-HA2 group showed synergic effects of these two peptides on each other's neutralizing properties.
In the TIV group, the HI response was significantly higher than other groups. In the M2e-HA2 group, because of the nature of HI test and conformational differences between HA2 and M2e peptides and HA1, a weak HI response was predictable. There was also a significant correlation between the level of neutralizing antibody and HI titer in the test groups' responses. Perhaps a better protective response for TIV groups comparing with M2e-HA2 groups can be related to this fact that TIV stimulates higher HI titer and it is normal and predictable that a vaccine with higher HI titer shows a higher protective response than a subunit vaccine such as M2e-HA2. However, the higher neutralizing antibody can induce a higher rate of natural selection against the immunity potential of the vaccine through genetic drift and in universal vaccine aspect, the invariability of the candid peptide is the most important factor.
However, there was a protective response for TIV groups against both H1N1 and H3N2 challenges, but as TIV contains both H1N1 and H3N2 viruses, the protection cannot be considered a real cross protection. With regard to the origin of M2e and HA2 from H1N1 subtype, the protective response against H2N3 subtype should be considered as a true cross protection. In influenza universal vaccines, cross protection is more important than efficacy. In an influenza pandemic case, a cross-protective vaccine can give primary protection, although poorly, to give time to the public health authorities to prepare a specific vaccine against the pandemic virus. The results of this study showed that the cross protection of M2e-HA2, as the fusion peptide originated from H1N1 subtype virus, could protect mice against H1N1 and H3N2 viruses.
Subunit vaccines can be manufactured in the bioreactor cell culture system with high efficiency and it can be a supremacy in case of an avian influenza pandemic in which shortage of embryonated eggs is normal. In addition, the use of embryonated eggs in whole virion vaccine manufacturing needs a period of time for virus/egg adaptation and that the bioreactor cell culture system can prepare the subunit vaccine in a shorter time period than in pandemic condition is very important. There are some strategies to overcome the lower efficiency of this fusion peptide, such as increasing the units of M2e or/and HA2 units on the contracture, higher concentration of the fusion peptide in each dose, and usage of stronger adjuvants that can be considered in future researches.
The areas for further research include expression of the HA2-M2e fusion protein in the eukaryotic system to modify the protein secondary modifications such as glycosylation, immunization studies on other animal models, study on humoral and cellular immunity responses, use of the fusion peptide with different adjuvants, and application of the construct as a DNA vaccine. The use of effective adjuvants is very important in subunit vaccine cases. In this study, we used Stimmune® as an adjuvant because of working on mice model; this adjuvant has veterinary application and to adapt the study on man model, it will need to change to MF59 or other acceptable adjuvants for human.
Footnotes
Acknowledgments
The results described in this article were a part of student thesis written by Ali Ameghi and was financially supported by a grant from the Immunology Research Center (IRC), Tabriz University of Medical Science (Grant No.123/90). We thank our colleagues from “Razi Vaccine and Serum Research Institute” (Iran) who provided us facilities and equipment. We like to show our gratitude to two anonymous reviewers for their so-called insights and comments.
Author Disclosure Statement
No competing financial interests exist.