
Abstract
The novel H7N9 influenza virus has been infecting humans in China since February 2013 and with a mortality rate of about 40%. This study compared the pathogenicity of the H7N9 and 2009 pandemic H1N1 influenza viruses in a ferret model, which shows similar symptoms to those of humans infected with influenza viruses. The H7N9 influenza virus caused a more severe disease than did the 2009 pandemic H1N1 influenza virus. All of the ferrets infected with the H7N9 influenza virus had died by 6 days after infection, while none of those infected with the 2009 pandemic H1N1 influenza virus died. Ferrets infected with the H7N9 influenza virus had higher viral titers in their lungs than did those infected with the 2009 pandemic H1N1 influenza virus. Histological findings indicated that hemorrhagic pneumonia was caused by infection with the H7N9 influenza virus, but not with the 2009 pandemic H1N1 influenza virus. In addition, the lung tissues of ferrets infected with the H7N9 influenza virus contained higher levels of chemokines than did those of ferrets infected with the 2009 pandemic H1N1 influenza virus. This study suggests that close monitoring is needed to prevent human infection by the lethal H7N9 influenza virus.
Introduction
I
In March 2013, the Chinese Center for Disease Control and Prevention reported the first case of human infection with a novel influenza A virus, H7N9, to which humans did not have immunity (2,10,16,27,31). As of March 3, 2015, 277 deaths had occurred out of a total of 602 laboratory-confirmed human infections with the H7N9 influenza virus (13). Those infected with the H7N9 influenza virus suffered from acute respiratory distress syndrome with fever, cough, pneumonia, lymphocytopenia, and thrombocytopenia (10,11,29).
Genetic analysis of the novel H7N9 influenza virus showed that all of the genes of this virus were of avian origin (10,16). The HA gene of the H7N9 virus belongs to the Eurasian lineage of avian influenza viruses and shares 95% homology with the HA genes of low pathogenicity avian influenza A (H7N3) viruses isolated in 2011 in China (16). The NA gene of the H7N9 virus is 96% identical to those of low pathogenicity avian influenza A (H11N9) viruses isolated in the Czech Republic (16). The sequences of the remaining genes are closely related to those of avian influenza A (H9N2) viruses that have recently been circulating in poultry in China.
In April 2009, a swine-origin novel H1N1 influenza A virus infected humans in Mexico and the United States, and spread to numerous other countries around the world (5,9,18,23). It was estimated that more than 200,000 humans died of the novel 2009 H1N1 infections (6). The 2009 pandemic H1N1 influenza virus contained gene segments that have not been reported in other swine or human influenza viruses. Its HA, NP, and NS genes belonged to the classical swine lineage; its PB2, PB1, and PA genes were related to the swine triple reassortant lineage; and its NA and M genes originated from the Eurasian swine lineage (12).
The previous studies showed the pathogenesis of H7N9 influenza viruses in ferrets and mice (2,27,31). H7N9 influenza viruses were more pathogenic in ferrets than the seasonal H3N2 influenza virus was (2). H7N9 influenza viruses were more pathogenic in mice than the 2009 pandemic H1N1 influenza virus was (27). Moreover, the various strains of H7N9 influenza viruses showed different pathogenesis in ferrets (31).
This study compared the pathogenicity between the 2009 pandemic H1N1 influenza virus and the novel H7N9 influenza virus in ferrets to find out the virulence of the pandemic potential H7N9 influenza virus.
Materials and Methods
Virus
Novel H7N9 (A/Anhui/1/2013) and 2009 pandemic H1N1 (A/California/07/2009) influenza viruses were kindly provided by the World Health Organization collaborating Center for Disease Control and Prevention (Atlanta, GA), and were grown in 10-day-old hens' eggs before being used for this study.
Animals
Ferrets aged about 7–8 weeks were bought from Path Valley Farm (Spring Run, PA), and the ferrets that were serologically negative to human influenza viruses (2009 pandemic H1N1, H3N2, and human B viruses) and the H7N9 influenza virus by hemagglutination (HA) inhibition assay were used for this study. All animal experiments were performed at an enhanced biosafety level 3 facility approved by the Korean government.
Ethics statement
Animal studies were performed according to the guidelines and recommendations of the Guide for the Care and Use of Laboratory animals of the Korean National Veterinary Research and Quarantine Service, and were approved by the Committee on the Ethics of Animal Experiments of Chungnam National University.
Studying clinical signs in infected ferrets
Ferrets (n = 10 per group) were infected intranasally (i.n.) with 0.5 mL of saline solution containing 107 × EID (egg infectious dose)50 of H7N9 (A/Anhui/1/2013) or the 2009 pandemic H1N1 influenza virus (A/California/07/2009). An uninfected control group was used for comparison. Respiratory clinical signs and mortality in the infected ferrets were monitored daily for 10 days post-infection (dpi), and body weight and rectal temperature were recorded for 10 dpi. The ferrets were infected with 107EID50/mL rather than 106EID50/mL, since this dose caused the enhanced mortality in ferrets by the H7N9 influenza virus, which causes about 40% mortality in humans.
Collection and histopathological staining of ferrets' tissues
To collect tissues, ferrets (n = 10 per group) were infected i.n. with 0.5 mL of saline solution containing 107 × EID50 of H7N9 (A/Anhui/1/2013) or the 2009 pandemic H1N1 influenza virus (A/California/07/2009). The ferrets were euthanized with high doses of Zoletil® at 3 and 5 dpi, and thoroughly exsanguinated prior to the collection of tracheae and the left cranial lobes of lungs, which showed more pathological damage in ferrets infected with both viruses. Tissues were fixed by submerging them in 10% (w/v) neutral buffered formalin and embedding them in paraffin. Tissues of 5 μm sections were stained with hematoxylin and eosin (H&E) as described (1). The stained tissues were evaluated under an Olympus DP70 microscope (Olympus Corporation, Tokyo, Japan).
Viral titration in the lung tissues of infected ferrets
One-gram parts of the left-lung cranial lobes of the same ferrets used for histopathological staining were homogenized, then frozen and thawed three times in 1 mL of phosphate-buffered saline (PBS; pH 7.4) supplemented with 2× antibiotic–antimycotic solution (Sigma, St. Louis, MO). A dilution series of each sample was prepared by performing serial 10-fold dilutions in PBS (pH 7.4), and each diluted sample was inoculated into 10-day-old embryonated eggs. The presence of virus in the inoculated eggs was identified by HA assay. Viral titers were determined by log10EID50/g as described (21).
Quantification of proinflammatory cytokines and chemokines by quantitative real-time polymerase chain reaction
Total RNA was collected from the left-lung cranial lobes (1 g) of the same ferrets (n = 3) used for histopathological staining and peripheral blood mononuclear cells of the same ferrets (n = 3 per group) used for histopathological staining by TRIzol® reagent (Invitrogen, Carlsbad, CA). cDNA was synthesized using the collected RNAs. SYBR® Green-based real-time polymerase chain reaction (PCR) was performed using a Rotor-Gene 6000 apparatus (Corbett, Mortlake, Australia) and SensiMix™ Plus SYBR (Quantace, London, United Kingdom) based on the recommendations of the manufacturer. Samples were run in duplicate, and each reaction contained a total volume of 20 μL, comprising 2 μL cDNA, 10 μL SYBR mixture, and proinflammatory cytokine-specific or chemokine-specific primers for ferrets as described (19). Cytokine and chemokine expression levels in tissues were normalized to those of ferret glyceraldehyde-3-phosphate dehydrogenase. The results of real-time PCR were quantified by the comparative threshold method after subtracting the data for uninfected control ferrets.
Statistical analysis
Statistical significance was determined by a two-tailed, paired Student's t-test. p-Values of <0.05 were regarded as significant.
Results
Clinical signs of ferrets infected with H7N9 or 2009 pandemic H1N1 influenza virus
Ferrets were infected i.n. with the H7N9 influenza virus (A/Anhui/1/2013) or the 2009 pandemic H1N1 influenza virus (A/California/07/2009), and their body temperatures, body weights, and mortality were monitored for 10 dpi. Ferrets infected with both viruses showed elevated body temperatures (Fig. 1A). The body temperatures of ferrets infected with the H7N9 influenza virus peaked at 2 dpi, with a mean rectal temperature of 40.3°C, while those of ferrets infected with the 2009 pandemic H1N1 influenza virus peaked at 1 dpi, with a mean rectal temperature of 40.2°C. The ferrets infected with the H7N9 influenza virus showed greater losses of body weight than did those infected with the 2009 pandemic H1N1 influenza virus (Fig. 1B). At 5 dpi, the uninfected ferrets gained as much as 43% of their body weight compared with their weight before infection. Ferrets infected with the 2009 pandemic H1N1 influenza virus gained as much as 10% of their body weight. However, ferrets infected with the H7N9 influenza virus lost as much as 23% of their body weight. A high rate of mortality was observed in ferrets infected with the H7N9 influenza virus (Fig. 1C). All of the ferrets infected with the 2009 pandemic H1N1 influenza virus survived, but all of the ferrets infected with the H7N9 influenza virus had died by 6 dpi.
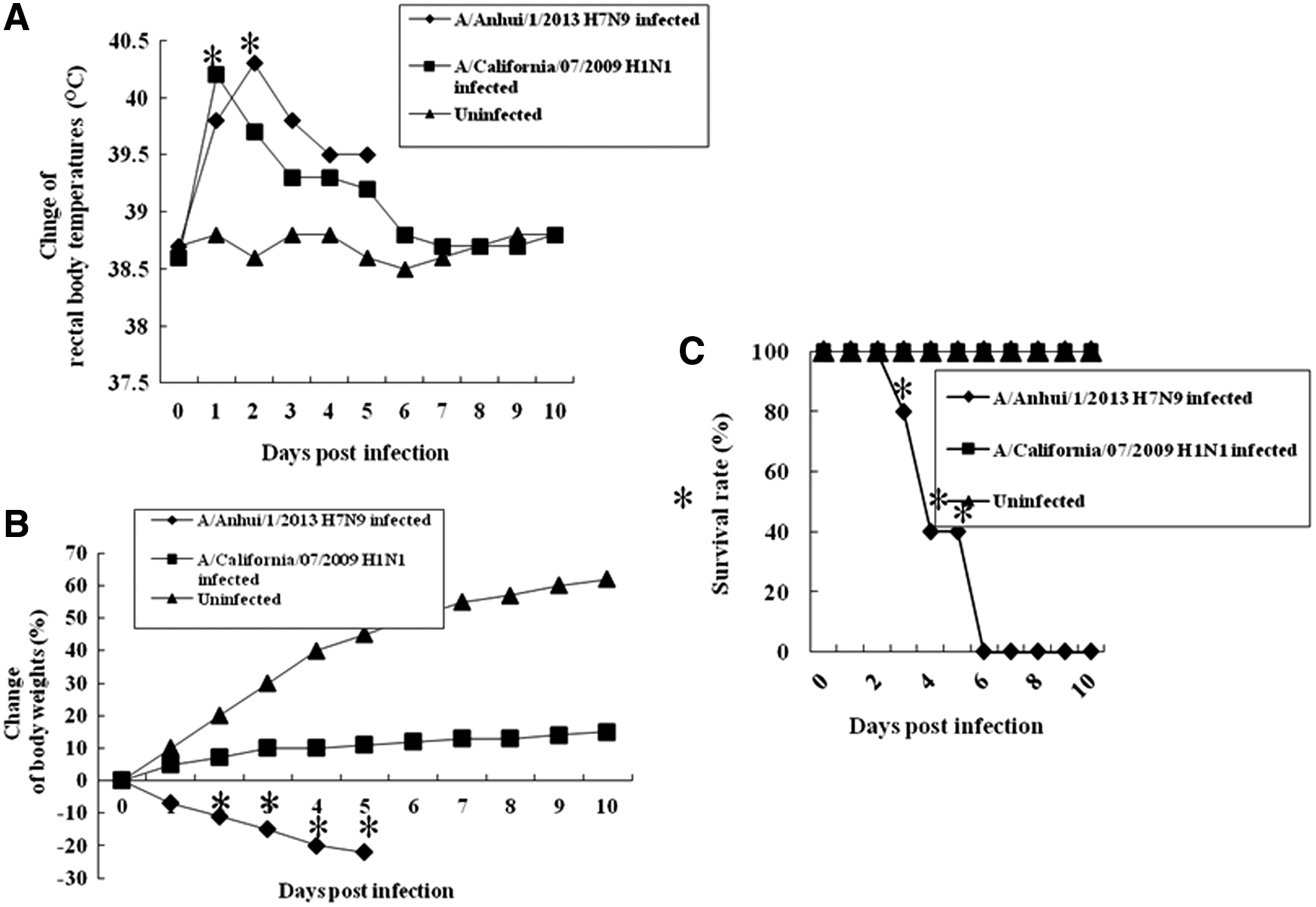
Body temperatures, body weights, and survival rate in infected ferrets. Ferrets (n = 10 per group) were infected intranasally (i.n.) with 107 EID(egg infectious dose)50/mL of (A/Anhui/1/2013) or 2009 pandemic H1N1 influenza virus (A/California/07/2009). The infected ferrets were monitored for 10 days post-infection (dpi). (
Gross pathology of the lungs of ferrets infected with the H7N9 and 2009 pandemic H1N1 influenza viruses
We examined the gross pathological damage to the lungs of infected ferrets at 3 and 5 dpi. The lungs of ferrets infected with the H7N9 influenza virus suffered from more severe damage than did those of ferrets infected with the 2009 pandemic H1N1 influenza virus (Fig. 2). The lungs of ferrets infected with the H7N9 influenza virus showed severe hemorrhagic pneumonia (Fig. 2A-1, A-2, B-1, and B-2), while those of ferrets infected with the 2009 pandemic H1N1 influenza virus showed mild pneumonia (Fig. 2C-1, C-2, D-1, and D-2). The lungs of uninfected ferrets appeared normal (Fig. 2E-1 and E-2).
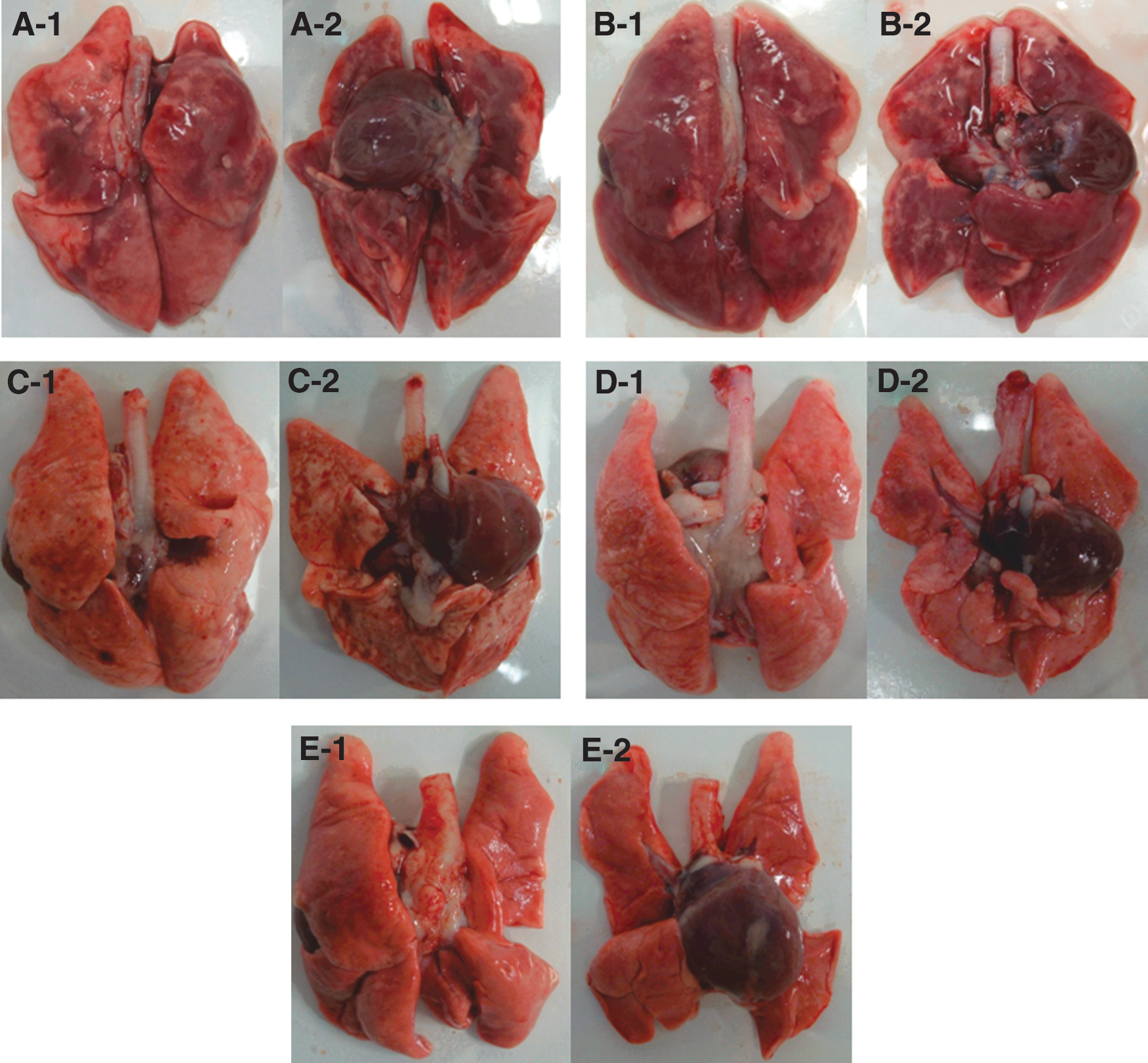
Gross lesions of lungs of infected ferrets. Ferrets (n = 6 per group) were infected i.n. with 107 EID50/mL of (A/Anhui/1/2013) or 2009 pandemic H1N1 influenza virus (A/California/07/2009), and the survived ferrets were euthanized at 3 and 5 dpi before the lung lesions were photographed. (
Tissue histopathology of ferrets infected with the H7N9 and 2009 pandemic H1N1 influenza viruses
To investigate the histopathological damage to the tissues of respiratory tracts, H&E staining was performed for tracheal, bronchial, and lung alveolar tissues collected from ferrets infected with the H7N9 and H1N1 influenza viruses at 3 and 5 dpi. Overall, the lower respiratory tracts of ferrets were more damaged in those infected with the H7N9 influenza virus, while the upper respiratory tracts of ferrets were more damaged in those infected with the 2009 pandemic H1N1 influenza virus (Figs. 3 –5).
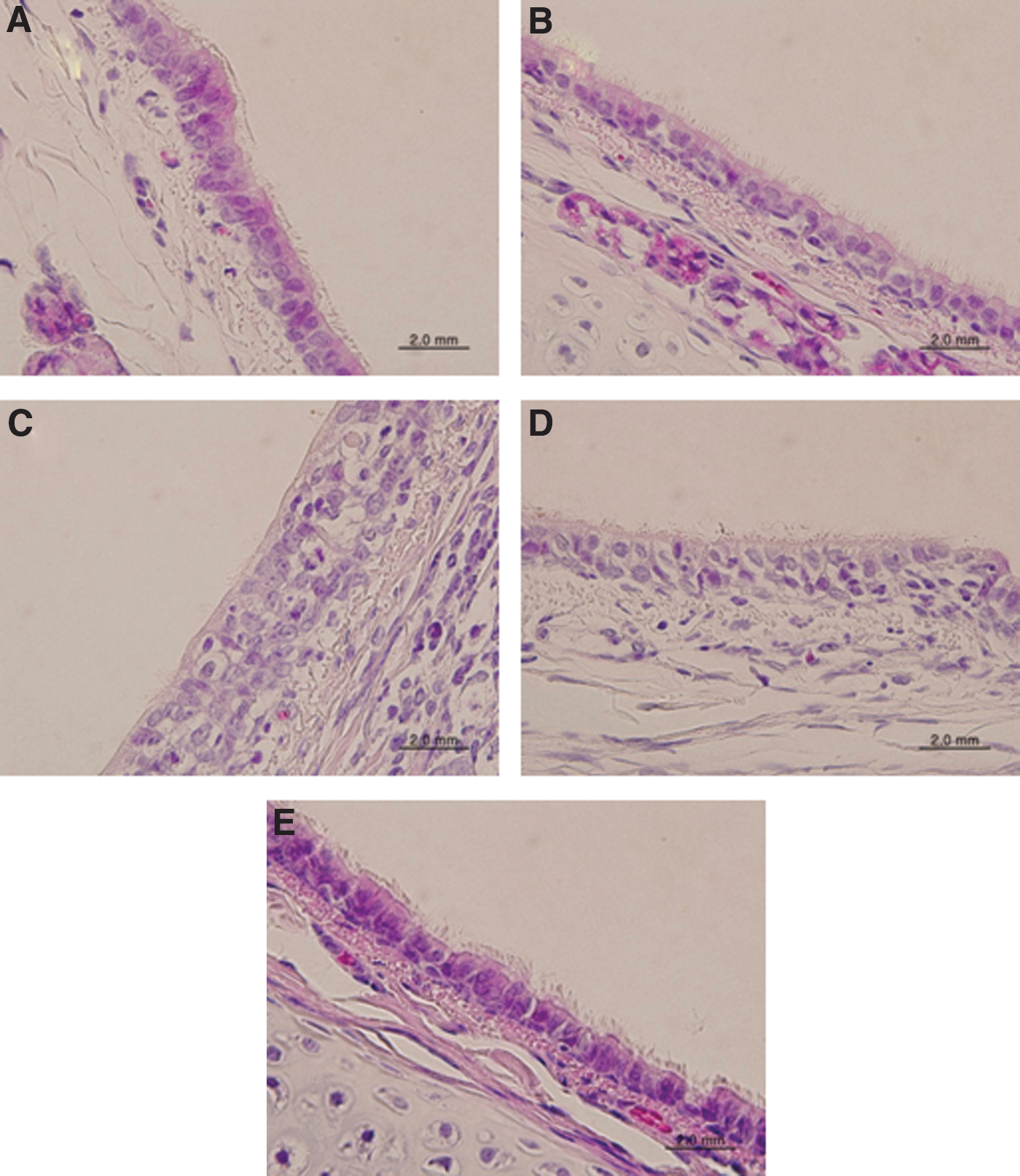
Histopathology of trachea tissues in the infected ferrets. Ferrets (n = 10 per group) were infected i.n. with 107 EID50/mL of (A/Anhui/1/2013) or 2009 pandemic H1N1 influenza virus (A/California/07/2009), and the survived ferrets were euthanized at 3 and 5 dpi before the tracheae were collected and processed for hematoxylin and eosin (H&E) staining. (
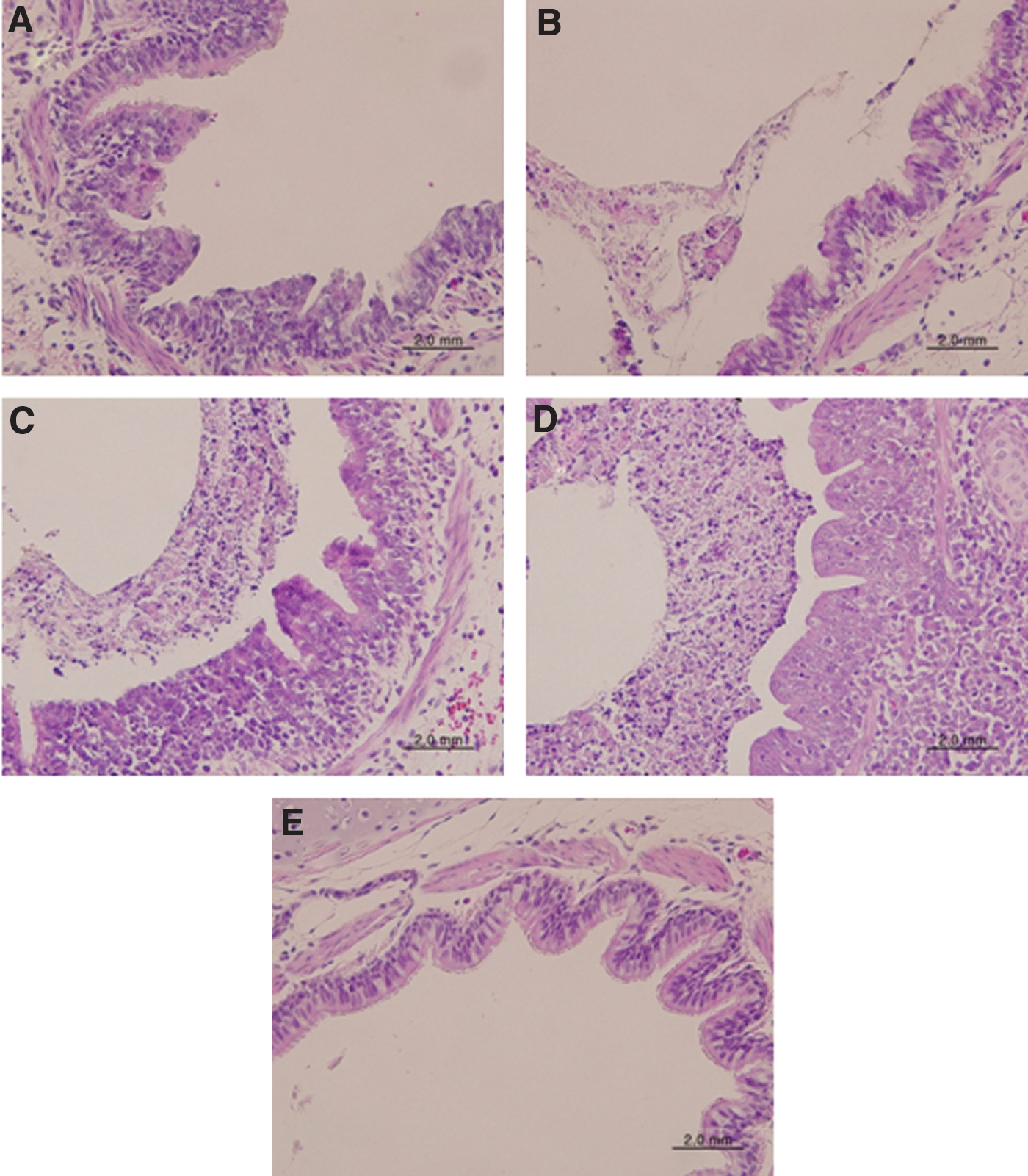
Histopathology of bronchus tissues in infected ferrets. The bronchus tissues from the same ferrets in Fig. 3 were stained with H&E staining. (
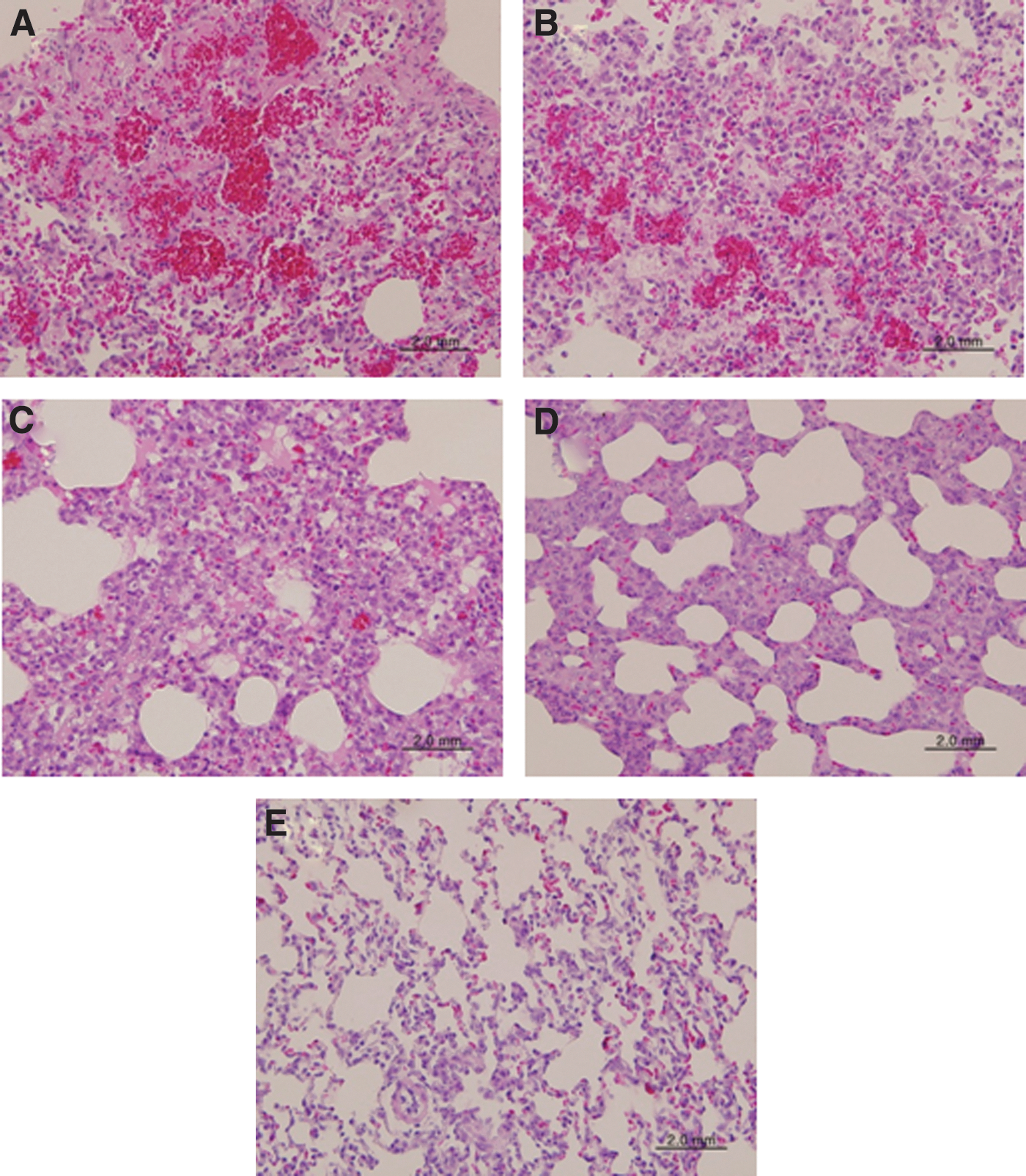
Histopathology of alveolar tissues of lungs in infected ferrets. The alveolar tissues of lungs from the same ferrets in Fig. 3 were stained with H&E staining. (
The tracheal tissues of ferrets infected with the H7N9 influenza virus were almost intact, with clear cilia and no inflammatory cells under the sub-mucus layers (Fig. 3A and B). However, those of ferrets infected with the 2009 pandemic influenza virus were damaged, with detached cilia and infiltration by inflammatory cells under the sub-mucus layers (Fig. 3C and D). The tracheal tissues of uninfected ferrets showed no damage, with clear cilia and no infiltration by inflammatory cells under the sub-mucus layer (Fig. 3E).
The bronchial tissues of ferrets infected with the H7N9 influenza virus showed signs of minor damage, with mucus in the lumen and a low level of infiltration by inflammatory cells under the sub-mucus layers (Fig. 4A and B). However, those of ferrets infected with the 2009 pandemic influenza virus were severely damaged, with thick accumulated mucus in the lumen and a high level of infiltration by inflammatory cells under the sub-mucus layers (Fig. 4C and D). The bronchial tissues of uninfected ferrets were normal, with no mucus in the lumen and no infiltration by inflammatory cells under the sub-mucus layer (Fig. 4E).
The lung alveolar tissues of ferrets infected with the H7N9 influenza virus were severely damaged, with hemorrhaging and a high level of infiltration by inflammatory cells in the interstitial lesions (Fig. 5A and B). However, those of ferrets infected with the 2009 pandemic influenza virus were mildly damaged, with a low level of infiltration by inflammatory cells in the interstitial lesions (Fig. 5C and D). The lung alveolar tissues of uninfected ferrets were normal, with intact interstitial lesions (Fig. 5E).
Viral titers and induction of proinflammatory cytokines and chemokines in the lung tissues of ferrets infected with the H7N9 and 2009 pandemic H1N1 influenza viruses
To determine the reason for the H7N9 influenza virus causing more severe disease to ferrets than did the 2009 pandemic H1N1 influenza virus, the viral titers and proinflammatory chemokines and cytokines in the lung tissues of infected ferrets were measured at 3 and 5 dpi (Figs. 6 and 7).
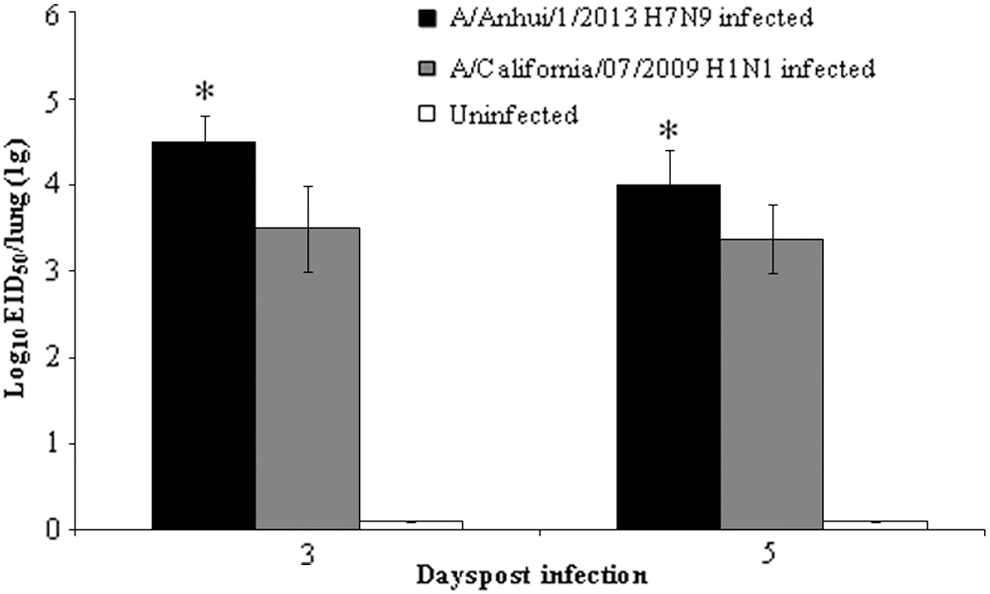
Viral titers in the lungs of infected ferrets. Parts (1 g) of the left-lung cranial lobes of ferrets used for Fig. 3 were homogenized, and viral titers were determined by log10EID50/g. Statistical analysis was performed to compare data from infected ferrets between H7N9 and H1N1. *p < 0.05.

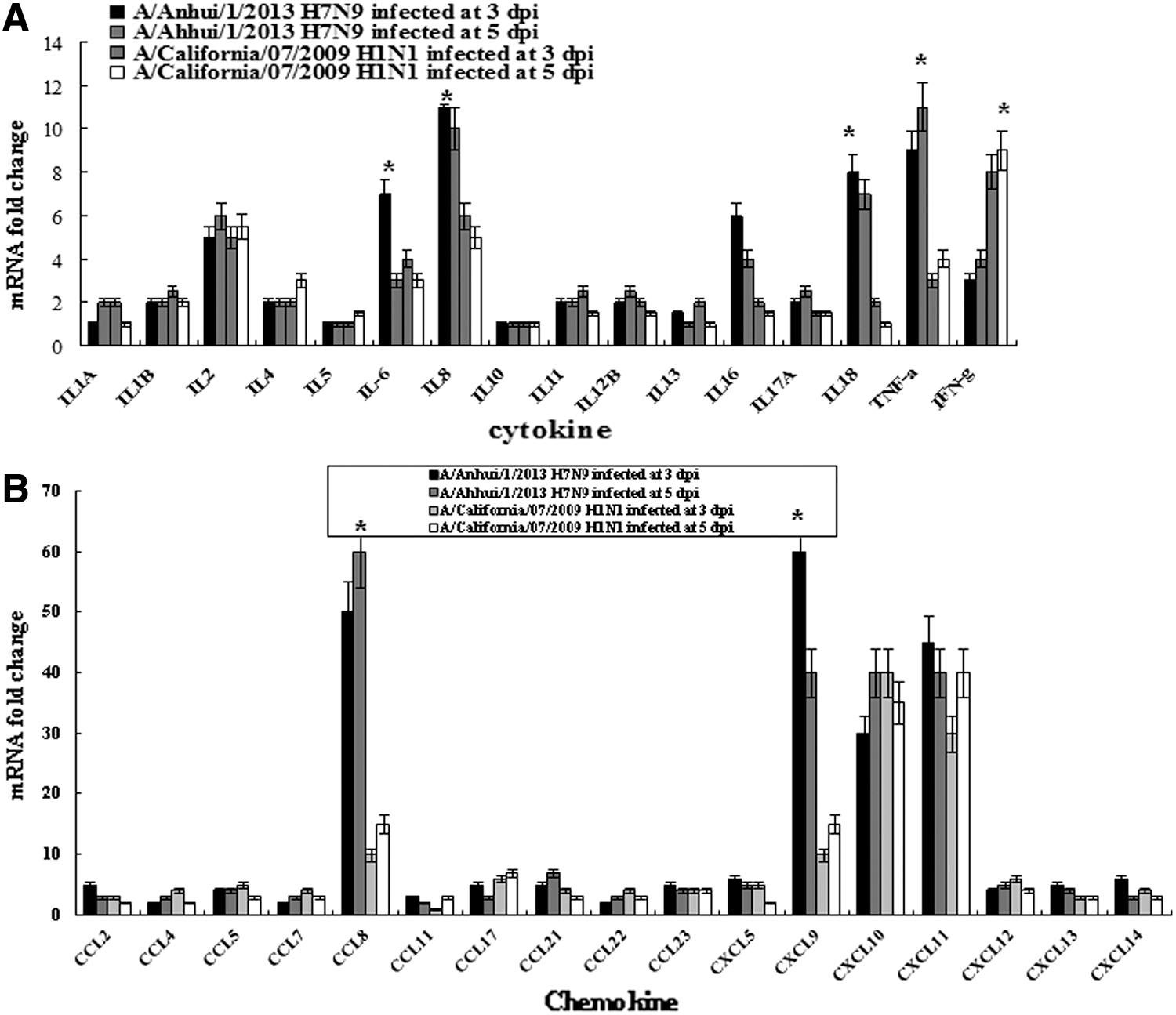
Induction of inflammatory cytokines and chemokines in the lungs of infected ferrets. Total RNAs were collected from lung tissues from the same ferrets used for Fig. 3, and inflammatory cytokine and chemokine expressions were quantified using specific primers for each gene by SYBR real-time polymerase chain reaction (PCR). The expression level of each gene in the tissues was normalized to that of ferrets' GAPDH. Statistical analysis was performed to compare data from infected ferrets between H7N9 and H1N1. (
Viral titers were higher in the lung tissues of ferrets infected with the H7N9 influenza virus than they were in those infected with the 2009 pandemic influenza virus (Fig. 6). The mean viral titers of 104.5 and 104.0 × EID50/g were detected in the lung tissues of ferrets infected with the H7N9 influenza virus at 3 and 5 dpi, respectively, whereas titers of 103.5 and 103.5 × EID50/g were found in the lung tissues of those infected with the 2009 pandemic H1N1 influenza virus at 3 and 5 dpi, respectively.
The proinflammatory cytokine IL-8 was dominantly induced (increased by 10-fold) in the lung tissues of ferrets infected with the H7N9 influenza virus, while the antiviral response inducing cytokine IFN-γ was dominantly induced (increased by 16-fold) in the lung tissues of ferrets infected with the 2009 pandemic H1N1 influenza virus (Fig. 7A). The T cell chemoattractant chemokine CXCL9 was greatly induced (increased by 90-fold) in the lung tissues of ferrets infected with the H7N9 influenza virus, but not in those infected with the 2009 pandemic H1N1 influenza virus (Fig. 7B). In the blood, proinflammatory cytokines such as IL-6, IL-8, IL-18, and TNF-α were induced higher in the ferrets infected with the H7N9 influenza virus, with 7-, 11-, 8-, and 9-fold increases at 3 dpi, respectively, than in those infected with the 2009 pandemic H1N1 influenza virus, with 4-, 6-, 2-, and 3-fold increases at 3 dpi, respectively. However, IFN-γ was induced higher in the blood of ferrets infected with the 2009 pandemic H1N1 influenza virus, with a ninefold increase at 5 dpi, than those ferrets infected with the H7N9 influenza virus, with a fourfold increase at 5 dpi (Fig. 8A). In the blood, chemokines, CCL8, and CXCL19 involved in attracting mononuclear cells were induced higher, with 48- and 60-fold increases at 3 dpi, respectively, than those infected with the 2009 pandemic H1N1 influenza virus with eight- and sevenfold increases at 3 dpi, respectively (Fig. 8B).
Induction of inflammatory cytokines and chemokines in the blood of infected ferrets. Total RNAs were collected from peripheral blood mononuclear cells from the same ferrets used for Fig. 3, and inflammatory cytokine and chemokine expressions were quantified using specific primers for each gene by SYBR real-time PCR. The expression level of each gene in the tissues was normalized to that of ferrets' GAPDH. Statistical analysis was performed to compare data from infected ferrets between H7N9 and H1N1. (
Discussion
A novel H7N9 influenza virus to which most humans lack immunity continues to circulate in China. This aim of this study was to compare the pathogenicity of the H7N9 influenza virus to that of the 2009 pandemic H1N1 influenza virus in ferrets. Ferrets infected with the H7N9 influenza virus suffered from a far more severe disease than did those infected with the 2009 pandemic H1N1 influenza virus, with hemorrhagic pneumonia and a higher induction of proinflammatory cytokines and chemokines.
All of the ferrets infected with the H7N9 influenza virus had died by 6 dpi, while all of the ferrets infected with the 2009 pandemic H1N1 influenza virus survived. This result may explain the mortality rate of about 40% among humans infected with the H7N9 influenza virus. It is surprising that the H7N9 influenza virus—the HA gene of which does not have polybasic amino acids in the cleavage site, which is a typical characteristic of the highly pathogenic H5 and H7 subtypes of influenza viruses (15)—can cause such high fatality. In a previous study using mice, the H7N9 influenza virus showed greater lethality than did the H9N2 influenza virus (2). That study showed that a three-pronged transcriptomic signature was detected in the lungs of mice infected with the H7N9 influenza virus, and the signature they detected was similar to that found in the lungs of mice infected with the 1918 pandemic H1N1 influenza virus, which is estimated to have killed more than 50 million humans (20). A study on the contribution of the viral genes to the lethality of the H7N9 influenza virus in mice showed that the PB2, M, and NP proteins were responsible for its high virulence in mice (3).
The lung tissues of ferrets infected with the H7N9 influenza virus were heavily damaged, with a high level of infiltration by inflammatory cells. A study with cynomolgus macaques reported that the histopathology of the lung tissues of monkeys infected with the H7N9 influenza virus were characterized by alveolar damage, the infiltration of polymorphonuclear cells, hyaline membrane formation, and pneumocyte hyperplasia (
The present study showed that the H7N9 influenza virus multiplied to higher titers and more strongly induced proinflammatory chemokines in the lung tissues of infected ferrets than did the 2009 pandemic H1N1 influenza virus. These findings correspond with previous findings in humans. A study using human cells showed that the H7N9 influenza virus induced proinflammatory chemokines in both human lung epithelial cells and endothelial cells (30). In blood, the present study showed that the proinflammatory cytokines were induced higher in ferrets infected with the H7N9 influenza virus than with the 2009 pandemic H1N1 influenza virus. Patients infected with the H7N9 influenza virus also contained higher levels of proinflammatory cytokines in their sera compared with those in the sera of healthy volunteers (14).
In conclusion, the H7N9 influenza virus is virulent in ferrets, and preventive measures are urgently needed to prevent human infections by this virus.
Footnotes
Acknowledgments
This work was funded by the 2014 research fund of Chungnam National University. This manuscript was edited by a scientific editor from Editage.
Author Disclosure Statement
No competing financial interests exist.