
Abstract
Cytotoxic T lymphocytes (CTL) are critical in cellular immune responses; therefore the study of CTL responses is profound in HIV-1 eradication. We aim to dissect the relationship between HIV-1 reservoir size and the magnitude and recognition of viral-specific CTL responses. An IFN-γ ELISpot assay with peptides spanning the HIV-1 clade C consensus sequences were designed to analyze HIV-1c-specific CTL responses. HIV-1 DNA, integrated HIV-1 DNA, and 2-LTR HIV-1 DNA were quantitated by real-time PCR. We observed significant increases in total HIV-1 DNA and integrated HIV-1 DNA after highly active antiretroviral treatment (HAART) compared with naive patients. Total HIV-1 DNA had a significant negative correlation with HIV-1c-specific CTL response magnitude. Baseline CD4+ T lymphocyte counts and antiretroviral treatment affected the size of the HIV-1 reservoirs. Taken together, HIV-1-specific CTL responses correlated with the size of HIV-1 reservoir. In addition, HIV-1-specific CTL response against p17 was associated with low integral efficiency of HIV-1, which might be a biomarker to evaluate the efficacy of HAART.
Introduction
H
Cellular immune responses are believed to play crucial roles in antiviral defense. It is well known that cytotoxic T lymphocytes (CTL) are crucial to cellular immune responses; therefore the study of CTL responses is profound toward HIV-1 eradication. A previous study observed strong HIV-1-specific CTL responses in HIV-1-infected individuals with long-term nonprogression (LTNP) (16), and another study also indicated an inverse correlation between HIV-1-specific CTL responses and viral load (22). However, the role of HIV-1-specific CTL response in the latent HIV infection is still not extensively studied. Previously Chun et al. investigated the suppression of HIV-specific CD8+ T cells on the HIV-1 replication in the resting CD4+ T-cell reservoir (10), and their results indicated that CD8+ T cells provided a potent suppressive activities against HIV replication in the latent viral reservoir through direct cellular contacts in patients who are naturally LTNPs and treated with HAART, but this antiviral activity was independent of CLT response. Furthermore, they found that early initiation of HAART in HIV patients may provide a sustaining antiviral activity to contain viral replication in patients with interrupted therapy (10, 29, 35).
An IFN-γ ELISpot assay with a total of 426 overlapping peptides spanning the HIV-1 clade C consensus sequences were applied to comprehensively analyze HIV-1-specific CTL responses. Moreover, an optimized real-time (RT)-PCR was utilized to measure integrated, 2-LTR circular, and total HIV-1 DNA in patients with HIV-1 infection. Our study may help to understand the relationship between HIV-1 reservoir size and cellular immune responses.
Materials and Methods
Study objectives
Ninety two HIV-1-infected individuals (58 males and 34 females) were recruited from the Department of Infectious Disease of Tangdu Hospital in Xi'an, China. Sixty six out of these patients were antiretroviral-treatment naive, and the other 26 subjects were undergoing HAART. The test subjects had a median age of 37 years (36.65 ± 9.52), and their median CD4+ T-lymphocyte count was 286 cells/μL (285.91 ± 260.39) and their median plasma viral load (log10 copies/mL) was 4.08 (4.08 ± 2.11). All 92 subjects were infected with HIV for at least 12 months. All studies were conducted with the approval of the ethics review board of Tangdu hospital, and each individual gave written informed consent for this study.
Viral load and lymphocyte subset test
HIV plasma viral load was analyzed by an immunofluorescence quantifying PCR assay (Roche Corporation) according to the manufacturer's instructions. T-lymphocyte subsets were enumerated by flow cytometry using TriTEST (CD4-FIITC/CD8-PE/CD3-PerCP, Becton Dickison) according to the “lyse-no-wash” procedure.
Synthetic HIV-1-peptides
A total of 426 synthetic 15–20 amino acid peptides overlapping by 10 amino acids spanning the entire HIV-1 clade C consensus sequences were synthesized at the Massachusetts General Hospital Peptide Core Facility on an automated peptide synthesizer using Fmoc technology.
ELISpot assays
Fresh peripheral blood mononuclear cells (PBMCs) were separated from whole blood by Ficoll–Hypaque (Sigma) density gradient centrifugation. 1 × 105 of cells/well were plated in 96-well polyvinylidene difluoride-backed plates (MAIP S45; Millipore), which had been previously coated with 100 μL/well of anti-gamma-interferon (IFN-γ) MAb 1-D1k (0.5 μg/mL; Mabtech) overnight at 4°C. Peptides were added directly to the wells at a final concentration of 10 μM with a final volume of 130 μL/well in RPMI 1640 with 10% of FBS. Wells containing PBMCs only were used as negative controls and were run in triplicate on each plate. Wells containing PBMCs stimulated with phytohemagglutinin (PHA) served as positive controls. The plates were incubated at 37°C with 5% CO2 overnight (14–16 h) and then washed six times with phosphate-buffered saline (PBS) before 100 μL of biotinylated anti-IFN-γ MAb 7-B6-1 (1 μg/mL; Mabtech) was added for 90 min of incubation. After being washed again with PBS, 100 μL/well of 1:20,000-diluted streptavidin–alkaline phosphatase conjugate (Mabtech) was added to all wells of the plates. The plates were incubated at room temperature for 45 min. Wells were washed with PBS again and incubated in dark for 20–30 min with 5-bromo-4-chloro-3-indolyl phosphate and nitroblue tetrazolium (Bio-Rad Laboratories). IFN-γ-producing cells were identified as darkly stained cells and were counted by an Elispot reader (AID EliSpot reader system; Autoimmune Diagnostika GmbH). The results were expressed as the number of spot-forming cells (SFC) per million PBMCs. The numbers of specific IFN-γ-secreting T cells were calculated by subtracting the negative control value from the calculated SFC count. Responses were considered positive if there were >50 SFC/106 PBMCs and had at least thrice more than the mean number of SFC of the negative controls. The overall specificity of using the IFN-γ ELISpot to test responses of CD8+ T cells to synthetic peptides was demonstrated previously by others in a series of CD8+ and CD4+ depletion and enrichment experiments (28, 31, 33, 34).
Generation of standards for real-time PCR assay
The primers and probes for real-time PCR were optimized based on published sequences of geographical variants in the Los Alamos HIV databases (
HIV DNA quantification
The total DNA from patients' PBMCs was extracted using a QIAamp DNA blood mini kit (Qiagen) according to the manufacturer's protocol. Modified real-time PCR was performed in 25 μL solution containing 2.5 μL of DNA target, 12.5 μL of Gene Expression Master Mix (Applied Biosystems 4369016), 1 μM of primers, and 0.2 μM of the probe under the following conditions: 95°C for 10 min, followed by 95°C for 15 s, and 60°C for 1 min (40 cycles) in an ABI 7500 PCR machine (Applied Biosystems). For the integrated HIV-1 DNA, the first round of PCR was conducted with the following conditions: 95°C for 8 min, and then 12 cycles of amplification: 95°C for 10 s, 60°C for 10 s, and 72°C for 170 s. One-tenth of the first round PCR product was used for following real-time PCR detection. The PCR signal from first round PCR without the Alu primers was subtracted from the total signals before the estimate of copy numbers. A standard curve was created for each run in a 7-log-unit range by 1:10 serial dilutions.
Statistical analysis
Statistical analysis and graphical presentation were performed using SPSS 13.0 and Sigma Plot 10.0. Student's t-test was used for data that fulfilled normal distribution. Data were expressed as the mean ± standard deviation (SD). The nonparametric Mann–Whitney U test was used to perform the comparison between Naive and On-HAART adults. Data were expressed as median values and interquartile range. Wilcoxon rank sum tests were used to compare the distribution of cumulative CTL responses to a particular peptide or protein. A rank-based test was used for this comparison because it appropriately accounts for the censoring of within-well Elispot responses of >50 spots/well. A response was considered positive if the probability of a cell forming a spot was higher in the test well compared to the negative control well. Spearman rank correlation was used for correlation analysis. All tests were two-tailed, and p-values of <0.05 indicated statistical significance.
Results
Characteristics of study population and HIV-1-specific CTL responses
Sixty six out of 92 HIV-1-infected patients were antiretroviral-treatment naive, and the other 26 subjects were already undergoing HAART for at least 6 months. The HAART consisted of two RT nucleoside analog inhibitors (NRTI) plus one RT non-nucleoside analog inhibitor (NNRTI) treatment. The general characteristics of the two study groups are presented in Table 1. The average age of the naive group was 36.52, and 37.00 in the On-HAART group. CD4+ T-lymphocyte count showed no statistical difference between the two study groups (p > 0.05). HIV-1 viral load was lower in the On-HAART group than in naive group, but this difference was not statistically significant (p > 0.05).
HAART, highly active antiretroviral treatment.
We assessed HIV-1-specific CTL responses in the PBMCs of two study groups using 426 overlapping peptides spanning all expressed HIV-1 proteins by Elispot assay. To identify responses with both high magnitude and high frequency, a normalized cumulative CTL response was used by combining data on CTL magnitude and frequency (21, 34). HIV-1c-specific CTL responses directed against all the HIV-1c proteins were detectable in the two groups (Fig. 1). The most immunogenic Nef, P24, and RT were targeted by 80.30%, 71.21%, and 72.73% of the tested naive patients, respectively. The HIV-1c-specific CTL responses against each HIV-1c protein was greatly reduced in the On-HAART group, especially in P15, Rev, and Vpu, whose CTL responses magnitude dropped by 92.55%, 90.45%, and 90.36%, respectively (Fig. 2A). However, there was less difference in the percentage of HIV protein recognition between the naive and On-HAART group (Fig. 2B). Compared to the naive group, recognition of P15, P17, and Rev in the On-HAART group were decreased by 42.89%, 32.05%, and 23.61%, respectively. Overall, P15 and Rev were the two proteins that showed the greatest change in response between naive patients and those on HAART.
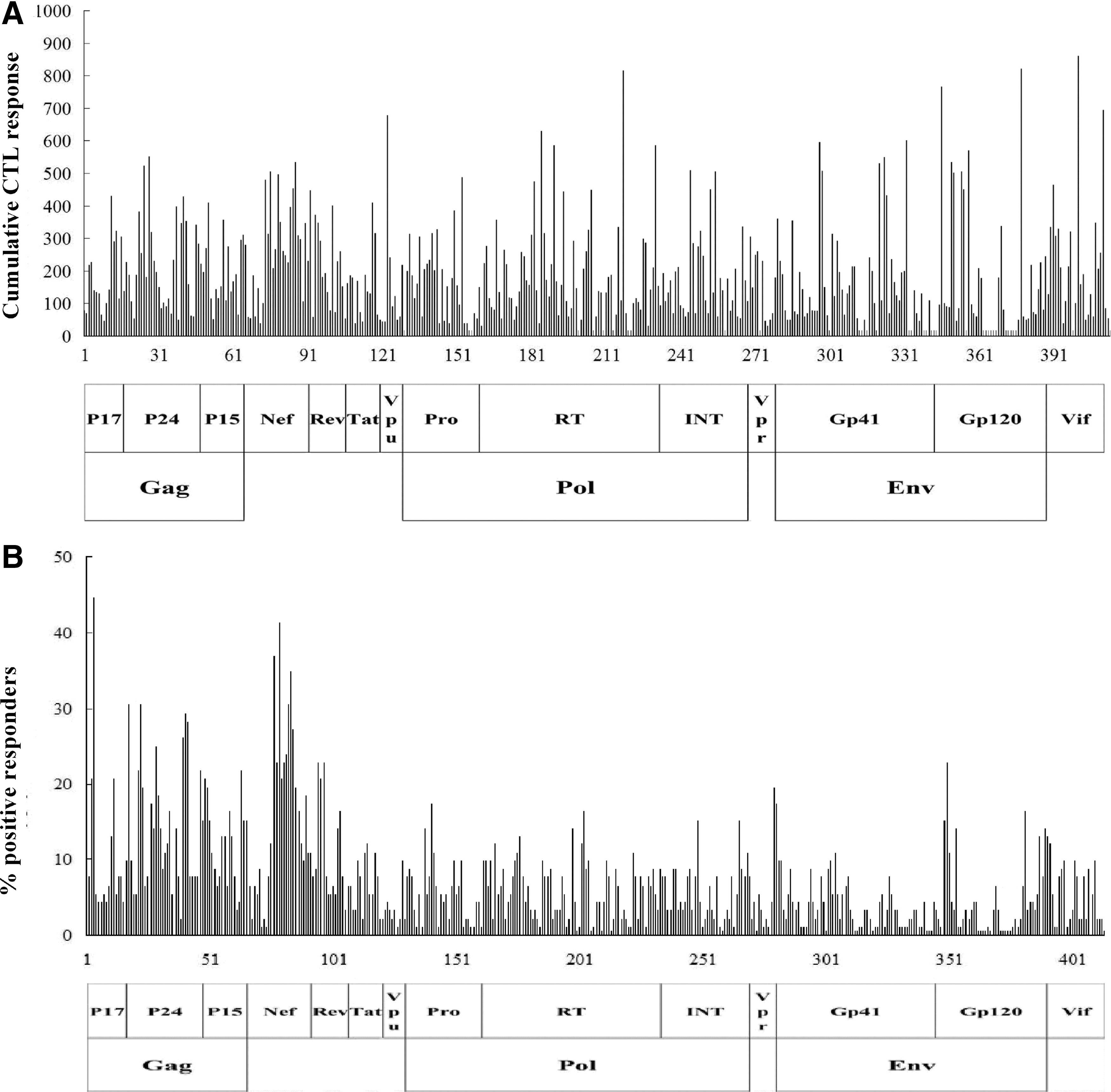
Intensity and breadth of HIV-1c-specific CTL responses in peptides levels. HIV-1-specific CTL responses were directed in the PBMCs of 66 naive patients and 26 HAART subjects using 426 overlapping peptides by IFN-γ-ELISpot assay.
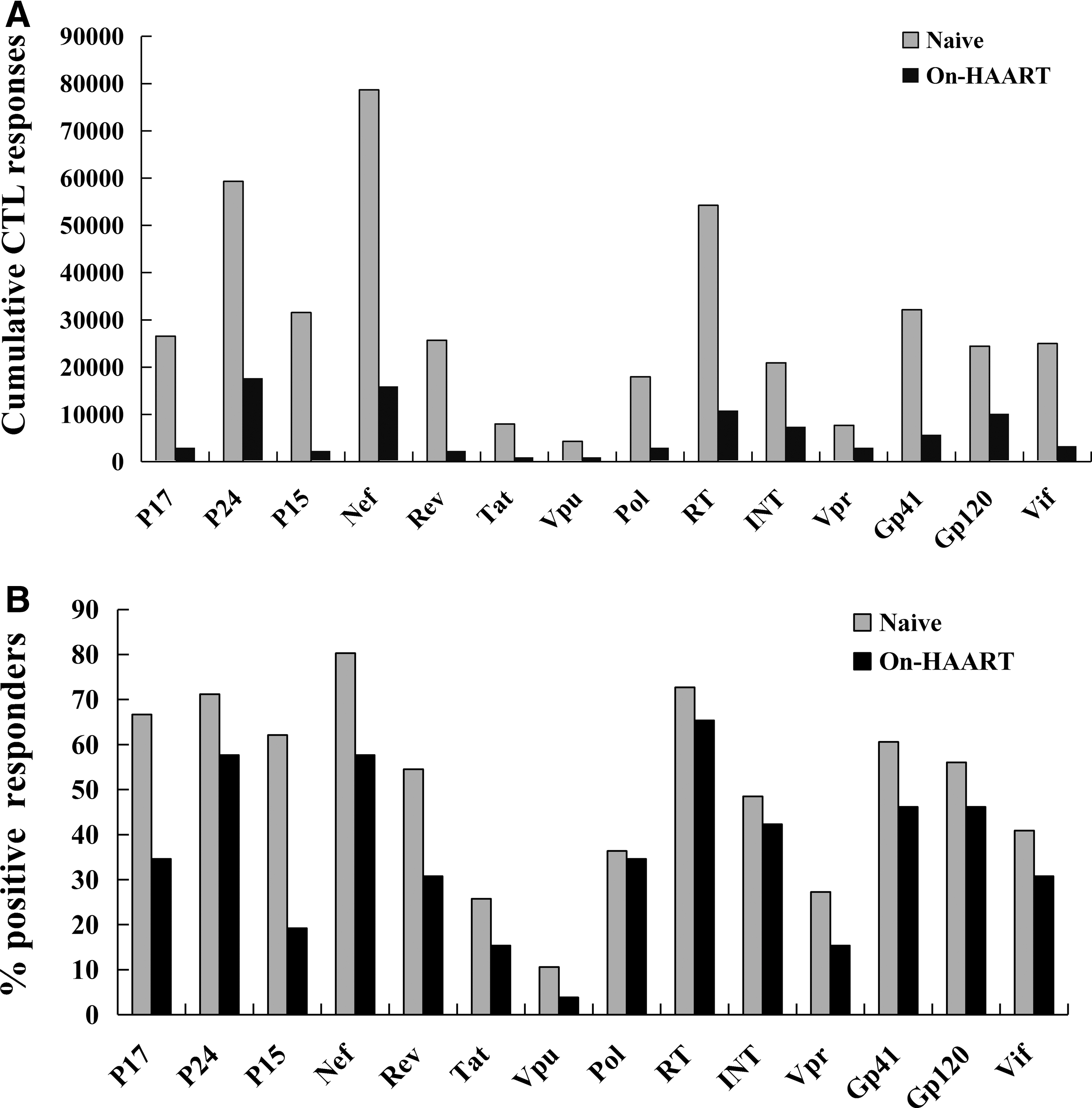
Magnitude and breadth of HIV-1c-specific CTL responses in protein levels. HIV-1-specific CTL responses were directed in the PBMCs of two study groups using 426 overlapping peptides spanning all expressed HIV-1 proteins by IFN-γ-ELISpot assay (x-axis).
Patients receiving HAART have higher total HIV-1 DNA and integrated HIV-1 DNA
After HIV-1 binding and entry into the host cells, the viral genome is reverse transcribed into the DNA, transported into the nucleus, and integrated into the host genome for viral replication (15). Cellular HIV-1 reservoirs are fundamental to latency and have three main existence forms (3). To identify the major HIV forms associated with its persistence, we measured the levels of total HIV-1 DNA, integrated HIV-1 DNA, and 2-LTR circles in the PBMCs from the 92 study subjects (8, 35). The results were shown in Figure 3. Interestingly, we found the median total HIV-1 DNA copies in the naive patients group were 8.79 (IQR: 2.400–50.340) copies/106 PBMC, while that in the On-HAART group was 31.03 (IQR: 13.370–107.540) copies/106 PBMC (Fig. 3A). The differences between the two groups were significant with a p-value of 0.0073. As for the integrated HIV-1 DNA (Fig. 3C), there was the same distinction between the two study groups as the quantity of integrated HIV-1 DNA of the On-HAART group [1.488 (IQR: 0–7.105) copies/106 PBMC] was significantly higher than the naive group [0.6978 (IQR: 0–9.819) copies/106 PBMC] (p = 0.0342). The result of 2-LTR circles (Fig. 3B) of the On-HAART group [3.086 (IQR: 0.047–7.105) copies/106 PBMC] was higher than the naive group [0.6978 (IQR: 0.391–3.225) copies/106 PBMC], but this difference was not significant (p = 0.3845).
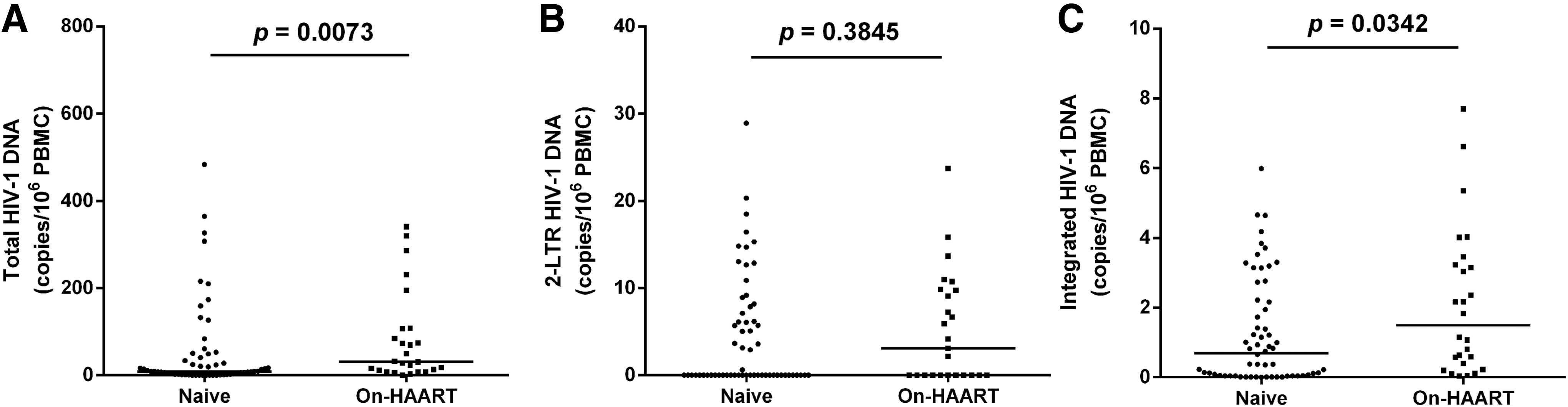
Comparison of HIV-1 DNA copies in PBMCs between naive and On-HAART of HIV-1-infected patients. Medians and p-values from Mann–Whitney test are shown.
Next, we counted the portion of integrated HIV-1 DNA from the total HIV-1 DNA detected in patients' PBMCs. As shown in Figure 4, integrated HIV-1 DNA in total HIV-1 DNA ranged from 0% to 7.7% for all the study subjects, which is smaller compared with 2-LTR circles with the value from 0% to 99.88%. The increase was highly significant for the proportion of integrated HIV-1 DNA between the naive group and the On-HAART group (Fig. 4B), whereas the increase in the proportion of 2-LTR was not significant between the naive group and the On-HAART group (p = 0.7658, Fig. 4A). Statistical analysis indicated that the proportion of integrated HIV-1 DNA was statistically significant between the two study groups (p = 0.0342).
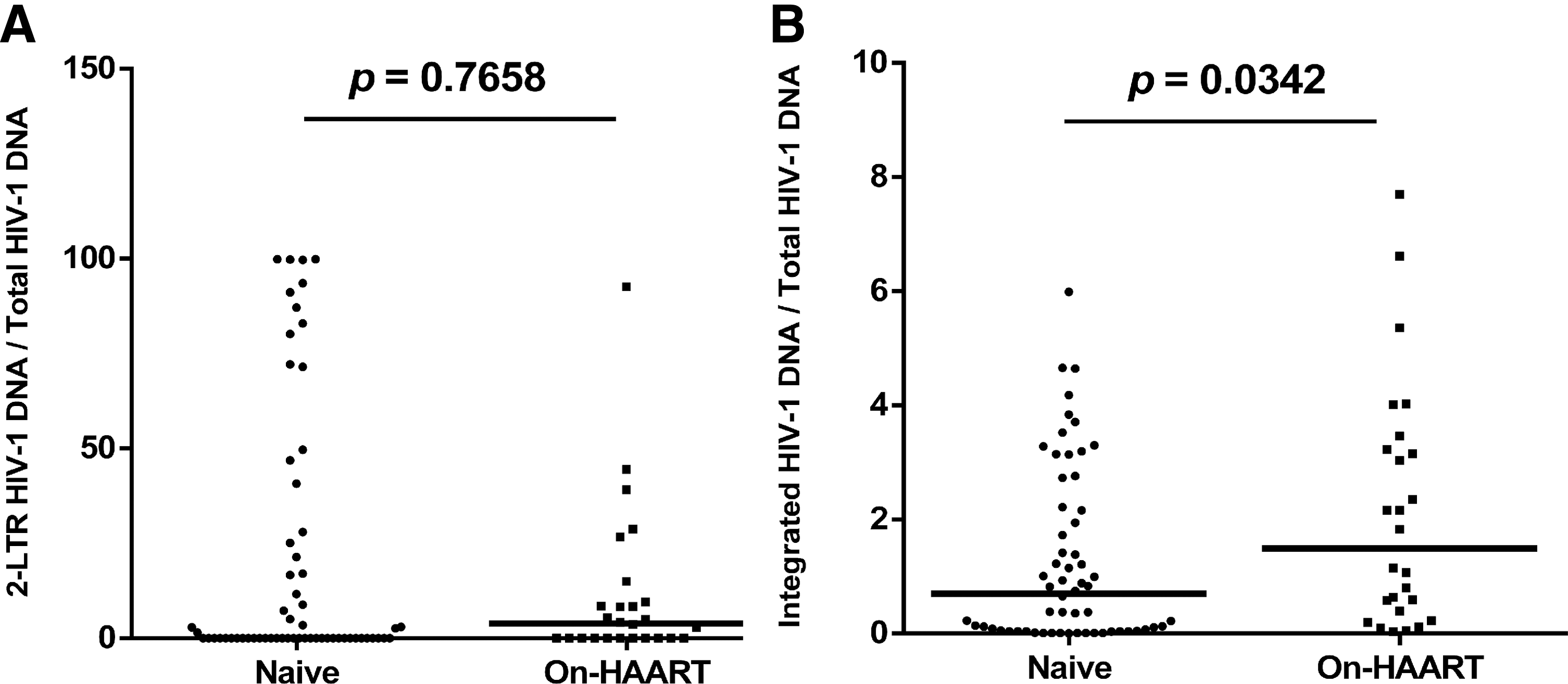
Comparison of the proportion of 2-LTR HIV-1 DNA and integrated in total HIV-1 DNA between naive and On-HAART of HIV-1-infected patients. Medians and p-values from Mann–Whitney test are shown.
Analysis of the correlation between HIV-1c-specific CTL responses and HIV-1 reservoir size
The magnitude of HIV-1c-specific CTL cumulative responses was significantly reduced after HAART treatment (Fig. 5). To illustrate the correlation between HIV-1-specific CTL responses and different forms of HIV-1 reservoirs, we evaluated the correlation effectors by using a nonparametric statistical method. The results are shown in Table 2. HIV-1-specific CTL cumulative responses negatively correlated with the absolute copies of HIV-1 DNA and relative proportion of 2-LTR circles and integrated HIV-1 DNA in both naive and On-HAART groups, who had the CD4+ T-cell counts that were greater than 350 cells/μL. However, the naive group had strong statistically significant correlations, while the On-HAART group presented no statistical significance.
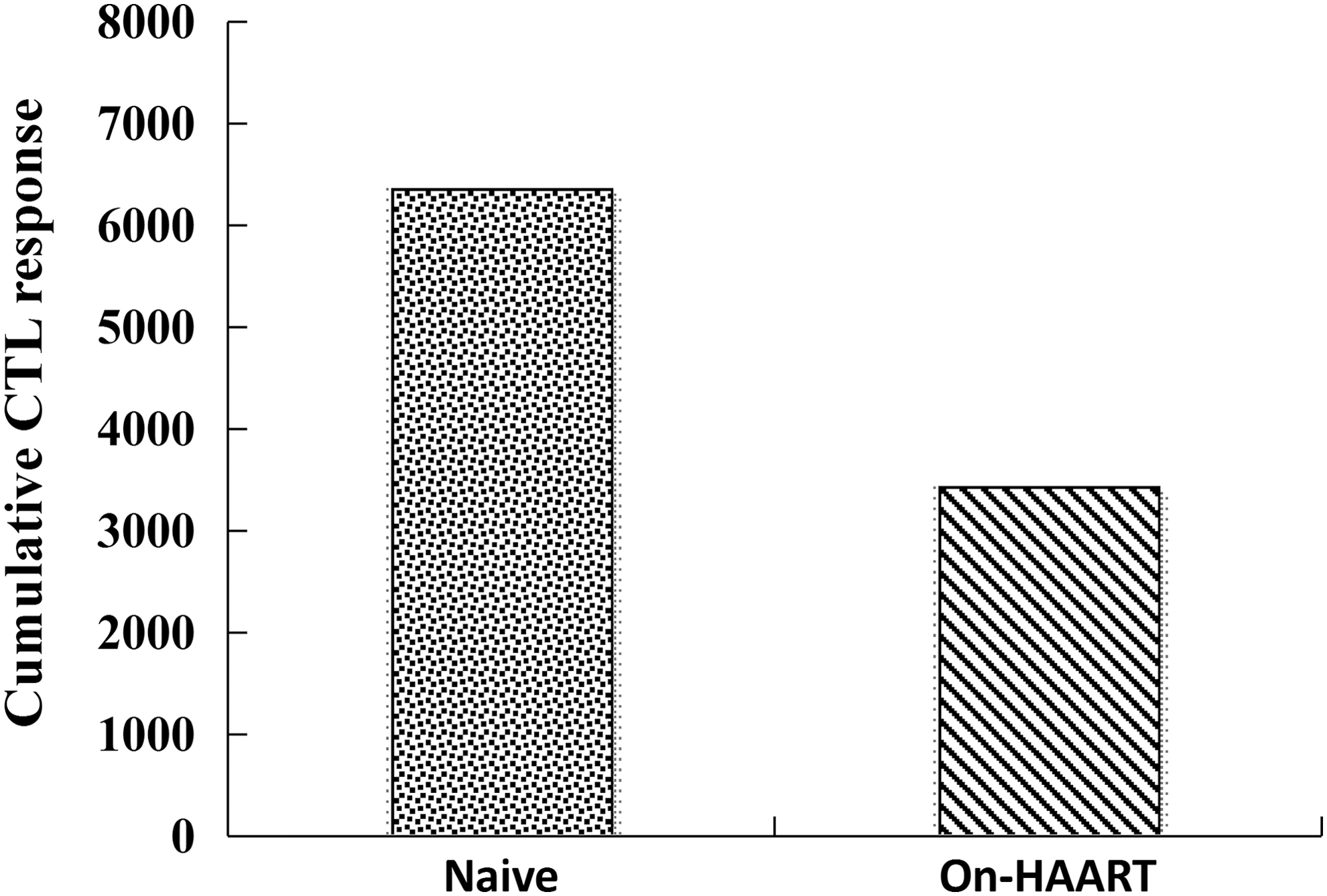
The magnitude of HIV-1c-specific CTL cumulative responses significantly reduced after HAART was initiated. Cumulative Elispot-based CTL responses were measured as a sum of naive or On-HAART of HIV-1 group to 426 overlapping peptides and expressed as the number of cumulative SFC/106 PBMC (y-axis).The peptide concentration was 10 μM with a final volume of 130 μL/well in RPMI 1640 with 10% of FBS.
Correlation is significant at the 0.01 level (two-tailed).
Correlation is significant at the 0.05 level (two-tailed). However, the naive group had strong statistically significant correlations, while the On-HAART group presented no statistical significance. These correlations disappeared after HAART which suggested that different immune cells could influence the HIV-1 reservoirs and reassured the necessary consideration of basic immune cell levels of study subjects.
CTL, cytotoxic T lymphocytes.
To gain more information, we counted the number of recognized HIV peptides that had statistically strong correlations with different forms of HIV-1 reservoirs; the results are shown in Table 3. Consistently, there were statistical correlations between the different forms of HIV reservoirs and CTL response magnitudes of different HIV-1c peptides in the naive group when CD4+ T-cell counts were greater than 350 cells/μL. There were five HIV peptides that had a statistical correlation with the amount of total HIV-1 DNA (P17, P24, Nef, Pro, RT, Gp41), while two peptides (P17, Nef) correlated with the relative proportion of 2-LTR circles of HIV-1. Six peptides (P17, P24, Nef, Pro, Int, RT, Gp41) correlated with the relative proportion of integrated HIV-1 DNA (p < 0.05). Only one peptide (P17) correlated with the proportion of integrated HIV-1 DNA when CD4+ T-cell counts were greater than 350 cells/μL in the On-HAART group. In general, the results indicated that the correlation between HIV-1-specific CTL responses and HIV-1 reservoirs was more strong in naive patients than in patients who had antiretroviral therapy. Of note, the high magnitude of HIV-1 CTL responses against P17 suggests that this peptide is very immunogenic to be correlated with all three forms of HIV-1 reservoirs. Likewise, P24, Nef, Pro, RT, and Gp41 were also important peptides that may affect HIV latent infection.
Interestingly, we observed that the correlations of CTL responses against peptides with integrated HIV-1 DNA were all positive (0.391–0.529), while the correlations with total HIV-1 DNA and the proportion of 2-LTR circles of HIV-1 were negative (−0.723 to −0.400). This phenomenon suggested that the weaker magnitudes of CTL responses against some peptides in the On-HAART group compared with the naive group, but the more efficient integration of HIV-1 DNA and more existent of 2-LTR circles into host cells, as well as a higher amount of total HIV-1 DNA.
Discussion
HIV-1-specific CTL responses provide an essential defense in controlling HIV-1 replication, especially in the early stage of infection. Meanwhile, HIV-1 reservoirs are established immediately after infection. The eradication of HIV-1 from infected patients is the ultimate goal of HIV therapeutic interventions. HIV-1 reservoirs have been identified to represent the major impediments for eradication. Recent studies indicated that elite suppressors (ES) had stronger HIV-1-specific CTL responses (17, 19, 24) and harbor low levels of integrated HIV DNA and high levels of 2-LTR circular HIV DNA compared with chronically infected progressors (14). These results inspire us to hypothesize that strong CTL responses could lead to the reduction of HIV DNA integration in CD4+ T cells because these targeting cells may express HIV proteins on cell surface. It has already been shown that antiretroviral therapy leads to weak HIV-specific CTL responses (2, 5, 23). However, whether there are any effects of HIV-specific CTL responses upon the initiation of HAART remains uncertain. Some researchers found there was an excess of unintegrated HIV DNA in patients on HAART (1). In this study, we detected HIV-1 specific CTL responses and three HIV-1 reservoirs forms (integrated HIV-1 DNA, 2-LTR circulating HIV-1 DNA, and total HIV-1 DNA) and thus tried to further explore the relationship of HIV-1-specific CTL responses with HIV-1 reservoirs.
Consistent with previous studies (31, 33, 34), we found that HIV-infected patients had a wide range of cumulative CTL responses against HIV-1 peptides and recognition of these peptides appeared to be extensive in Gag, Nef, and Env regions. Moreover, both the cumulative CTL responses and the recognition of HIV peptides were significantly weaker in the On-HAART group compared with the naive group. HIV-specific CTL responses became weaker with the initiation of HAART, but accompanied with more amounts of HIV reservoirs. Unexpectedly, our results are contradictory with those of Graf et al. (14). They showed that total and integrated HIV-1 DNAs both decreased in patients on HAART. We assumed that such discrepancy might be caused by the characteristics of study subjects or length on HAART. In their studies, the off-HAART group usually had a median of about 4,000 copies of total HIV DNA per mL PBMCs, but in our naive group, only a median of 53 copies of total HIV DNA was detected. Thus, their off-HAART group may represent the patients in rapid HIV progression, but our naive patients are more like ES with low total or integrated HIV DNAs.
In our study, CD4+ T-lymphocyte counts between the two study groups did not demonstrate statistical significance, which excluded the potential effect of CD4+ T-cell levels on the lower HIV-1 reservoirs in the On-HAART group. However, high CD4+ T-lymphocyte counts may affect HIV reservoirs. HIV-1 reservoirs became more strongly correlated with CTL cumulative responses when CD4+ T-lymphocyte counts were above 350 cells/μL. However, these correlations disappeared after HAART. These results suggested that different immune cells could influence the HIV-1 reservoirs and reassured the necessary consideration of basic immune cell levels of study subjects. Accordingly, CTL responses against different peptides are correlated with HIV-1 reservoirs. Also, the CTL responses were more remarkable when CD4+ T-lymphocyte counts were above 350 cells/μL, but less notable after HAART.
Interestingly, P17 peptide positively correlated with the amount of integrated HIV-1 DNA, but negatively correlated with the amount of 2-LTR DNA and total HIV-1 DNA. We propose that HIV-specific CTL responses against p17 may be associated with the low integrate efficiency of HIV-1, which might be a biomarker to evaluate the efficacy of HAART.
In summary, we analyzed the association of HIV-1-specific CTL responses and HIV reservoirs in patients with or without HAART. We observed an increase in total HIV-1 DNA and integrated HIV-1 DNA after HAART compared with naive patients. HIV-1c-specific CTL responses may be suppressed after HAART rather than HIV-1b-specific CTL responses. Moreover, we found that the baseline CD4+ T-lymphocyte counts and antiretroviral treatment affected the size of HIV-1 reservoirs. In addition, HIV-specific CTL responses against p17 were shown to correlate with low integrate efficiency of HIV-1, which can possibly be monitored to evaluate the efficacy of HAART.
Footnotes
Acknowledgments
This work was supported by the grant from National Key Technologies Research and Development Program of China (2008ZX10001-006, 2008ZX10001-002, and 2012ZX10001-003), Natural Science Foundation of China (grant 31000084 and 81202305), and Tangdu Technology Innovation and Development Program (2012LCYJ002).
This work was approved by the Ethics Board of Tangdu Hospital. The Judgment reference number was TDLL-2012016.
Author Disclosure Statement
No competing financial interests exist.