
Abstract
Classical swine fever (CSF) vaccine based on HAdV-5 had achieved an efficient protection in swine. Both classical swine fever virus (CSFV) E0 glycoprotein and E2 glycoprotein were the targets for neutralizing antibodies and related to immune protection against CSF. Interleukin-2 (IL2), as an adjuvant, also had been used in CSF vaccine research. In this study, coexpression of the CSFV E0, E2, and IL2 genes by HAdV-5 (rAdV-E0-E2-IL2) was constructed and immunized to evaluate its efficacy. Three expressed genes had been sequentially connected with foot-and-mouth disease virus 2A (FMDV 2A). The vaccine was administered by intramuscular inoculation to CSFV-free pigs (108 TCID50) twice at triweekly intervals. No adverse clinical signs were observed in any of the pigs after vaccination. The vaccine induced strong humoral and cellular responses that led to complete protection against clinical signs of lethal CSFV infection, viremia, and shedding of challenge virus. The rAdV-E0-E2-IL2 is a promising, efficient, and safe marker vaccine candidate against CSFV.
Introduction
C
IL2 as an adjuvant has been proven to enhance the antigen-specific humoral and cellular immunities by promoting lymphocyte proliferation and activation in lots of reports (2,10,11,18). In current CSF vaccine research, porcine IL2 could enhance the immunogenicity of an alphavirus replicon-based DNA vaccine by coinjecting (29). In addition, rabbits immunized with CSF-vectored vaccine based on HAdV-5, which coexpressed E2 and IL2, could be completely protected against CSFV C strain and induced higher the CEFV E2-specific antibody titer compared to the rAdV-E2 expressing only the E2 glycoprotein in our previous study (8).
On this basis, coexpression of the CSFV E0, E2, and porcine IL2 genes by HAdV-5 (rAdV-E0-E2-IL2) was constructed and immunized to evaluate its efficacy in this study. The results suggest that the vaccine induced strong humoral and cellular responses that led to complete protection from lethal CSFV challenge.
Materials and Methods
Reagents, cells, and viruses
The rAdVs were packaged, propagated, and titrated in human embryonic kidney (HEK) 293 cells. The virulent CSFV Shimen strain was provided by the Control Institute of Veterinary Bioproducts and Pharmaceuticals. CSFV C-strain vaccine (lot No. 2014005) is a commercially available vaccine manufactured by LvFang Biotech Co.
Construction of the rAdVs
The CSFV E0, E2 genes (a and b) (GenBank: AY77517 8.2) and IL2 gene (GenBank: AY775178.2) were amplified by polymerase chain reaction (PCR) using the specific primers CSFV E0 F/R, E2 F1/R1, E2 F2/R2, and porcine IL2 F/R, respectively (Table 1). The E0-E2 and E2-IL2 were obtained by overlap extension PCR (E0 and E2a, E2b and IL2), digested by the restriction enzymes BglII (amino acids 525–530 at full-length E2 gene), and then ligated by T4 DNA ligase. The obtained fragment E0-E2-IL2 was cloned into the adenoviral shuttle vector rAdTrack-CMV (SalI and NotI sites) to obtain plasmid rAd-Track-E0-E2-IL2.The plasmid was homologous recombined with rAdEasy-1 in AdEasier-1 host bacteria BJ5183 and then was packaged in HEK293 cells. The positive plasmid was identified by PCR, restriction enzyme digestion analysis, and sequencing. Subsequently, the plasmid was linearized with the Pac I restriction endonucleases and transfected into HEK293 cells for viral amplification and package. The rAdV-E0-E2-IL2 was propagated and titrated in HEK293 cells. CSFV E0 and E2 protein expressed by rAdV-E0-E2-IL2 infected 293 cells determined by Western blot and IL2 expression was detected by a porcine IL2-specific ELISA (Hailan).
The restriction sequences of SalI (GTCGAC), XbaI (TCTAGA), KpnI (GGTACC), and NotI (GCGGCCGC) are underlined. The 2A sequences of FMDV are italicized. The overlapping 2A sequences of FMDV on primers are bold and italicized.
CSFV, classical swine fever virus; IL, interleukin.
Immunization and challenge
According to the Guidelines for the Care and Use of Animals of Northwest A & F University, this study was carried out, and animal experiments were approved by Care and Use of Animals Center, Northwest A & F University.
Fifteen CSFV-free cross-bred piglets, which were 6–7 weeks old, were double confirmed to be free to CSFV-specific serum antibodies and antigens using the IDEXX HerdChek* CSFV Antibody Test Kit and real-time reverse transcription (RT-PCR), respectively, and were randomly divided into five groups (A, B, C, D, and E) with three animals in each group. Groups A, B, and C were vaccinated with a dose of 108 TCID50 of rAdV-E0-E2-IL2, rAdV-E0-E2, and rAdV-E2-IL2 by intramuscular (i.m.) inoculation, respectively. All pigs were given a booster immunization at 21-day intervals. Group D was immunized with one-dose CSFV C-strain vaccine and served as the positive control. Group E was injected with Dulbecco's modified eagle medium (DMEM). Finally, all animals were challenged with 1 × 103 TCID50 of CSFV Shimen strain by intramuscular injection at 21 days after the second vaccination. Following the challenge, pigs were monitored daily for clinical signs (fever, anorexia, depression, shivering, hemorrhage, constipation, and diarrhea) of disease, and rectal temperatures were recorded. At 12 days postchallenge (DPC), all surviving pigs were euthanized.
Blocking ELISA and neutralization peroxidase-linked assay
Serum samples were collected from all pigs at 0, 7, 14, 21, 28, 35, and 42 days after immunization. CSFV-specific neutralizing antibodies in sera were tested by using the IDEXX HerdChek* CSFV Antibody Test Kit according to manufacturer's instructions and neutralization peroxidase-linked assay (NPLA) (28).
Lymphocyte proliferation assays and detection of swine interferon-γ
Cell-mediated immunity was evaluated by measuring lymphocyte proliferation with the WST-1 Cell Proliferation and Cytotoxicity Assay Kit (Beyotime) according to the manufacturer's instructions. T lymphocytes were stimulated with complete virus antigens of CSFV Shimen strain (MOI = 100), Concanavalin A (5 μg/mL; Sigma), or DMEM. The stimulation index (SI) = (the mean of OD450nm for CSFV-stimulated cells)/(the mean of OD450nm for medium-stimulated cells) was used to evaluate lymphocyte proliferation.
Swine interferon-γ (IFN-γ) was measured in serum by using ELISA kits (XinLe) following manufacturer's instructions 3 weeks after the booster immunization.
Swine IFN-γ gene expressions were assessed through real-time RT-PCR. Three weeks after the booster immunization, the whole blood samples of all pigs were collected. Total RNA was extracted from mixture, which had been mixed with equal amount of three samples in the group. The following two pairs of primers were used: forward, 5′-TGGTAGCTCTGGGAAACTGAATG-3′ and reverse, 5′-GGCTTTGCGCTGGATCTG-3′ for swine IFN-γ gene; forward, 5′-CAAGGACCTCTACGCCAACAC-3′ and reverse, 5′-TGGAGGCGCGATGATCTT-3′ for β-actin gene. PCR was performed using the SYBR ExScript™ RT-PCR Kit (Takara Bio) in iQ5 Real-Time PCR Detection System (Bio-Rad) with the following procedures: 95°C for 3 min followed by 35 cycles of 95°C for 10 sec; 60°C for 20 sec and 72°C for 20 sec. The tests were determined in triplicate. The β-actin was used as endogenous control to normalize the quantification of target genes. Data were assessed using 2−ΔΔCt relative quantitative analysis according to the Ct value.
Virus isolation
Blood samples, as well as nasal and fecal swabs, were collected on day 0 after the challenge and every other day thereafter until pigs were euthanized. After freeze thawing thrice in phosphate-buffered saline, all samples were centrifuged at 8,000 g for 5 min, and the supernatant was used for viral isolation in PK15 cells. After 96 h, the monolayer cells were air dried, fixed for 2 h, and then incubated with anti-CSFV sera (diluted 1:100). After washing, the cells were incubated with Rabbit Anti-pig IgG/HRP antibody (diluted 1:200; BIOSS). Immunoreactivity of stained foci of cells was detected by a light microscope. If a negative result was identified, the supernatant was blind passaged thrice in cells and again subjected to CSFV detection.
CSFV RNA detected by real-time RT-PCR
The presence of CSFV in different tissues and organs was analyzed by the one-step quantitative real-Time RT-PCR assay as described previously (14). One-step Prime Script RT-PCR kit (Takara) and the primers: forward, 5′-GTTCTGCGAGGTGACCAAAAG-3′, reverse, 5′-GATGCACACATAAGTATGGTAAAGC-3′, and Probe 5′-(FAM) TCCGTCGCTACCTGTCACCCTACCT (Eclipse)-3′ were used under the following conditions: 20 min at 42°C, 30 sec at 95°C, and 40 cycles of 15 sec at 95°C, 20 sec at 57°C, and 15 sec at 72°C. Data were analyzed according to the Ct method. According to the established standard curve, the following formula: copy number = (45.665 − CT value)/3.4692 was implemented to calculate the copy number of CSFV RNA.
Statistical analysis
Data are presented as mean ± SEM. Differences in each group were examined for statistical significance using Student's t-test of SPSS 13.0 software. Differences of p < 0.05 were considered statistically significant.
Results
The package and identification of rAdV-E0-E2-IL2
The rAdV-E0-E2-IL2 was typically generated at day 9 and exhibited fluorescence in 293 cells. Cytopathogenic effects appeared in the 293 cells 24 h following infection with the obtained rAdV-E0-E2-IL2. The titer of rAdV-E0-E2-IL2 was maintained above the 108 TCID50/mL after five passages. The CSFV E0 and E2 protein expressed in 293 cells infected with rAdV-E0-E2-IL2 were confirmed by Western blot with the use of anti-CSFV serum (Fig. 1). Approximately 45- and 50-kDa-specific bands were clearly visible in the rAd-E0-E2-IL2-infected cell extracts. According to molecular weight of the above bands, the predicted proteins were CSFV E0 and E2, respectively. No specific bands were observed in the cell lysates of mock group and rAdV empty vector-infected group. In addition, IL2 expression in the supernatant of infected cells was confirmed by a porcine IL2-specific ELISA. The results showed that three proteins could be expressed correctly and independently in vitro.
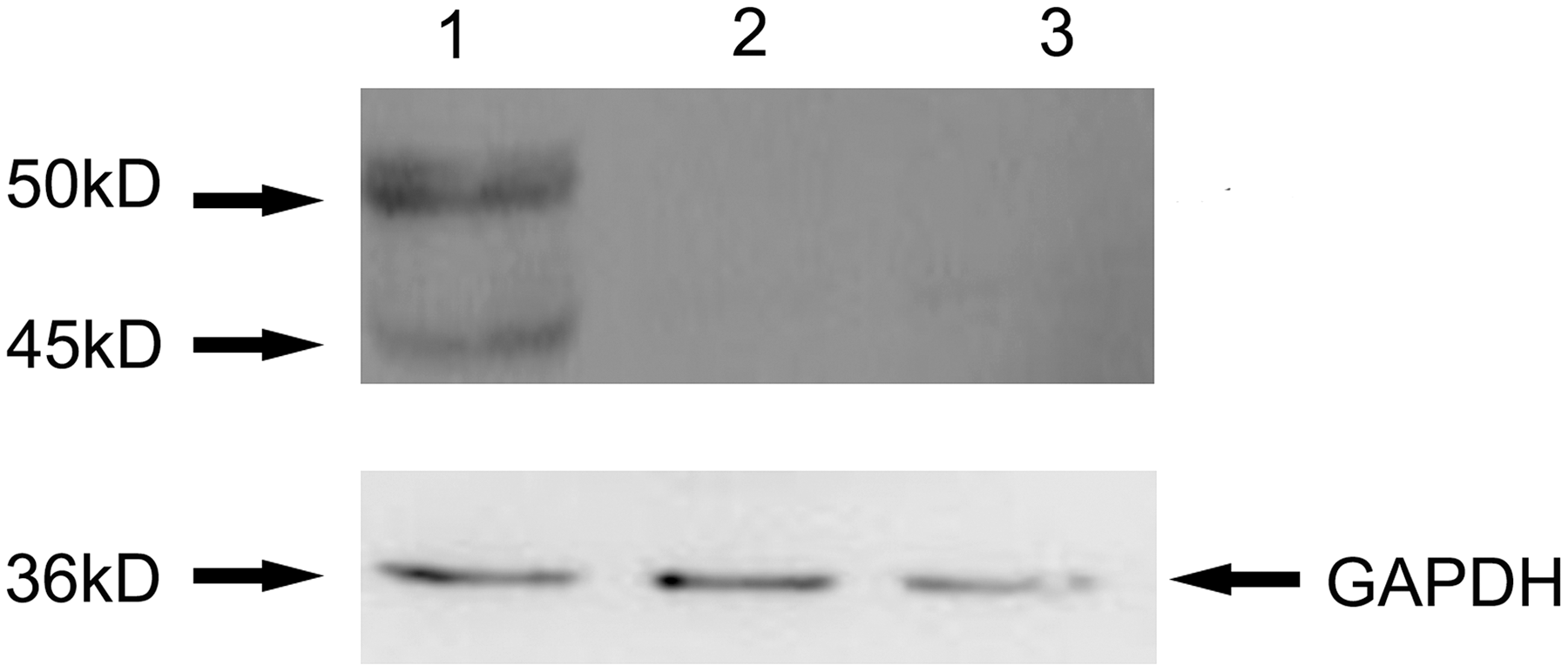
Western blot analysis of the E0 and E2 protein expression in rAdV-E0-E2-IL2 infected 293 cells. Lane 1, lysates of 293 cells infected by rAdV-E0-E2-IL2; lane 2, lysates of 293 cells infected by rAdV empty vector; lane 3, normal 293 cells lysates. The arrows indicate the bands of protein. The anti-CSFV sera (diluted 1:100) and Rabbit anti-swine IgG (diluted 1:1,000; Sigma) were used as the first antibody and the secondary antibody, respectively. Porcine GAPDH protein was determined by using a mouse anti-porcine GAPDH antibody (LifeSpan Biosciences). CSFV, classical swine fever virus; IL, interleukin.
Antibody production
After vaccinating, the CSFV E2-specific antibodies in the sera of all pigs were measured by NPLA and blocking ELISA. As shown in Table 2, the pigs immunized with rAdV-E0-E2-IL2, rAdV-E0-E2, and rAdV-E2-IL2 developed detectable antibody titers 2 weeks after the prime immunization. CSFV E2-specific antibodies were markedly increased after booster immunization. The CSFV E2-specific antibody titer of group A was higher than that in the groups B and C after the booster immunization. However, the CSFV E2-specific antibody titer of group A was lower than that in the group D after the booster immunization. The pigs immunized with DMEM (group E) did not develop detectable antibody titers (<4) during the experiment.
NPLA titers are expressed as the reciprocal of the serum dilution that neutralized 200 TCID50 of the CSFV “Shimen” strain in 50% of the replicate cultures.
First vaccination.
Second vaccination.
Challenge.
DMEM, dulbecco's modified eagle medium; NPLA, neutralization peroxidase-linked assay.
As shown in Table 3, the mean antibody blocking rate of the group A, B, C, D, and E was 56.7%, 52.0%, 45.3%, 63.0%, and 6.2%, 3 weeks after the booster immunization, respectively. According to the cutoff of the assay (40%), CSFV E2-specific antibodies were detected in the groups of A (3/3), B (3/3), C (3/3), D (3/3), and E (0/3), respectively. The mean antibody blocking rate of the groups A, B, C, and D increased gradually, especially the group D in which all pigs seroconverted at 35 days after immunization.
All serum samples were collected weekly and tested the anti-CSFV antibodies with the IDEXX HerdChek* CSFV Antibody Test Kit.
First vaccination.
Second vaccination.
Challenge.
Cell-mediated immune response
Cell-mediated immunity in immunized pigs was evaluated through CSFV-specific lymphocyte proliferation 3 weeks after the booster immunization. Lymphocyte proliferative responses were observed in groups A, B, C, and D. Cell-mediated immunity was the highest in group D, followed by group A and C. No CSFV-specific proliferation was observed in group E. Significant differences in SI were observed between the groups A and B (p < 0.01), or groups A and C (p < 0.05), whereas no significant difference in SI was observed between the groups A and D (p > 0.05) (Fig. 2).
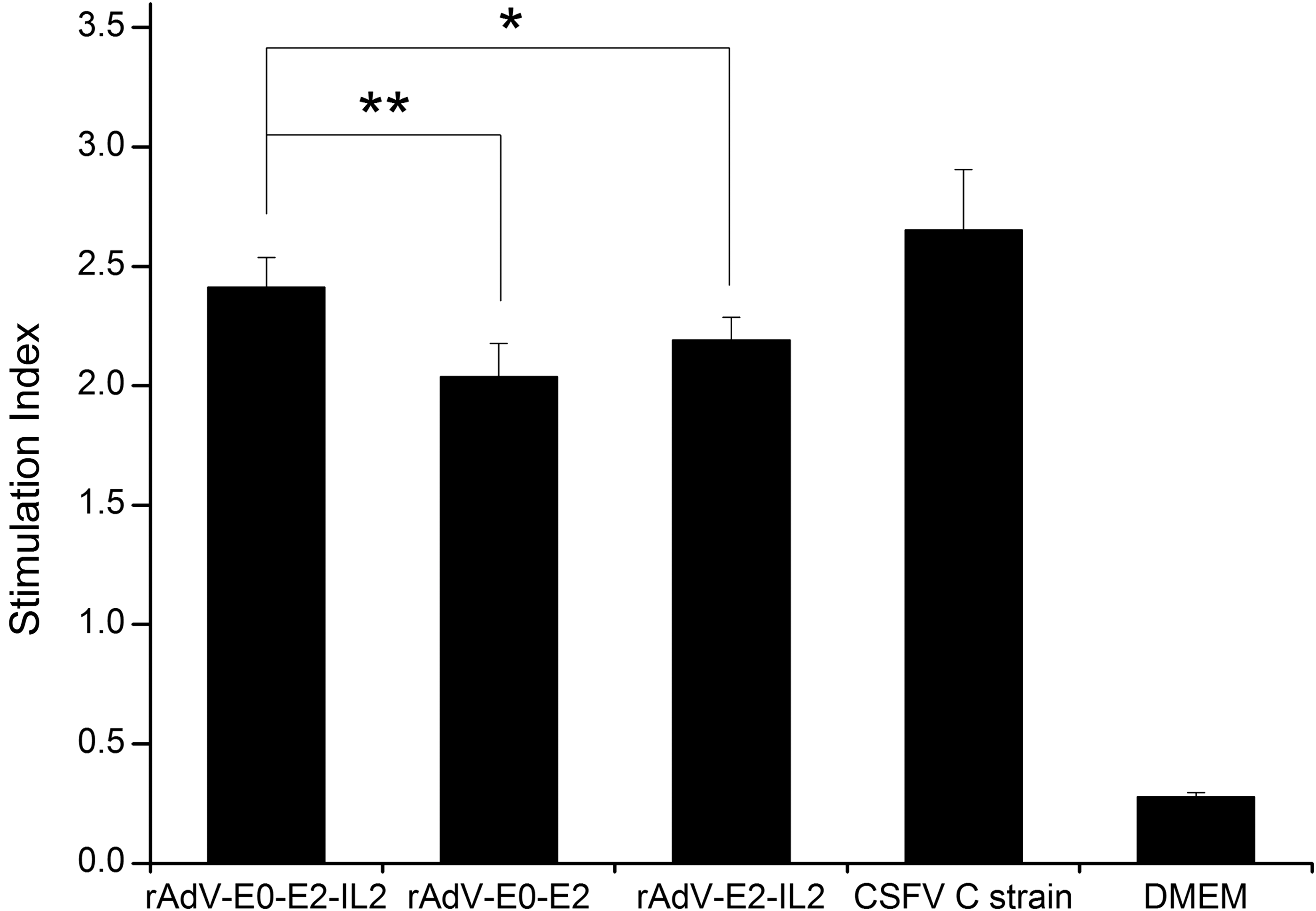
SI of the group rAdV-E0-E2-IL2. Significant differences in SI were observed between the rAdV-E0-E2-IL2 and rAdV-E0-E2 immunization groups (**p < 0.01), or rAdV-E2-IL2 immunization groups (*p < 0.05), whereas no significant difference in SI was observed between rAdV-E0-E2-IL2 and CSFV C strain immunization groups (p > 0.05). SI, stimulation index.
The level of antigen-specific IFN-γ production can be used as an indicator of cellular immunity. Swine IFN-γ content in the immunized pig sera was measured by using ELISA kits 3 weeks after booster immunization. We observed that there were significant differences in swine IFN-γ between the groups A and B (p < 0.01), or groups A and C (p < 0.05). However, we found no significant differences in swine IFN-γ content between the group A and D (p > 0.05) (Fig. 3).
Content of the Swine IFN-γ in the sera of experimental piglets. There were significant differences in swine IFN-γ content between the rAdV-E0-E2-IL2 and rAdV-E0-E2 (**p < 0.01), or rAdV-E2-IL2 immunization groups (*p < 0.05).
Swine IFN-γ gene expression was assessed in whole blood by using real-time RT-PCR. Figure 4 shows that after vaccination, the highest relative swine IFN-γ mRNA fold was detected in group D compared with the groups A, B, and C, 3 weeks after the booster immunization. Between the groups A and B, or C, there were significant differences in relative swine IFN-γ mRNA fold in the blood (p < 0.05). However, between the groups A and D, the differences in relative swine IFN-γ mRNA fold in the blood were unremarkable (p > 0.05) (Fig. 4).
The relative swine IFN-γ mRNA fold in the whole blood of experimental piglets. Between the rAdV-E0-E2-IL2 and rAdV-E0-E2, or rAdV-E2-IL2, the difference in relative swine IFN-γ mRNA fold in the blood was also remarkable (*p < 0.05).
Protection of immunized pigs from virulent challenge
Twenty-one days after the booster immunization with rAdV-E0-E2-IL2 or rAdV-E2-IL2 (day 42 p.i.), all experimental pigs were inoculated intramuscularly with CSFV Shimen strain (103 TCID50). Protective immunity was assessed through clinical signs, including body temperature. After challenge with virulent CSFV, no clinical signs were observed in pigs immunized with rAdV-E0-E2-IL2, rAdV-E0-E2, rAdV-E2-IL2, or CSFV C-strain. However, all pigs in Group E developed a high fever (rectal temperature above 40°C) from 2 DPC and gradually displayed the typical clinical signs of CSF (inappetence, apathy, chill, prostration, incoordination, and constipation, followed by diarrhea, locomotor ataxia, and posterior paresis). Finally, all the pigs died within 10 DPC.
The viruses were readily detected in all samples of group E between 4 to 10 DPC. However, the pigs immunized with rAdV-E0-E2-IL2, rAdV-E0-E2, rAdV-E2-IL2, or CSFV C-strain did not show any virus after lethal challenge.
All the survived pigs were euthanized and gross lesion of pigs was examined at 12 DPC. The pigs immunized with rAdV-E0-E2-IL2, rAdV-E0-E2, rAdV-E2-IL2, and CSFV C-strain did not present any gross lesion. The pigs in group E presented severe pathological changes of typical CSF (hemorrhages with necrotic foci in the tonsils, enlargement and hemorrhage of the lymph nodes, infarcts in the spleen, extensive petechiae in the kidney and bladder, and button-like ulcers in the ileocecal valve). Moreover, CSFV was detected from various organs (blood, spleen, kidney, intestines, lymphonodi mesenterici, and tonsil) of the pigs through real-time PCR. No viral RNA was noted in pigs immunized with rAdV-E0-E2-IL2, rAdV-E0-E2, rAdV-E2-IL2, or CSFV C-strain. CSFV RNA was also detected in all pigs vaccinated with DMEM. The copy number of CSFV RNA was detected in group E and the amount of CSFV virions was different in different tissue (103–106 copies/μL) of each pig. These results were consistent with gross lesion examination.
Discussion
In many countries, the morbidity and mortality of CSF were significantly reduced because of vaccination with live attenuated vaccine, which has been widely used. However, this vaccine was unable to discriminate naturally infected pigs from vaccine-injected pigs (Differentiating Infected from Vaccinated Animals [DIVA]) and unacceptable to “stamping-out” policy. Therefore, developing a new type of CSF marker vaccine is necessary. To achieve this, numerous candidate vaccines surrounding CSFV or its structural protein E2 and/or Erns, which can induce virus-NAbs and offer protective immunity in the natural host, were investigated by various strategies and methods. Among them, HAdV-5 could provide an effective vaccine delivery of a variety of agents to swine and had been demonstrated. Although various vaccines with high efficacy have been developed, a more potent adjuvant with defined and appropriate properties is required to improve the immunogenicity of many antigens.
A variety of cytokines secreted by many cells (macrophages, T cells, etc) can be used as immune modulator proteins to enhance the normal immune response. IL2, which is an important regulator of immunity, can induce differentiation and proliferation of activated T cells (from G0 phase to S phase), enhances natural killer cell cytotoxicity and activates monocytes and macrophages, and can promote lymphocyte proliferation, activation, and the production of antibodies and cytokines (22,23). In addition, IL2 also stimulates proliferation of activated B lymphocytes and induces immunoglobulin (IgM and IgG) secretion (3). A number of results published to date have clearly shown that costimulation of plasmids encoding the IL2 gene with different kinds of vaccines results in enhancement of both humoral and cell-mediated immune responses in immunized animals and mostly favors Th1 cell differentiation (6,12,32). IL2 is a promising cytokine adjuvant for inactivated and subunit vaccines as well as DNA vaccines (1,2,9 –11,18). IL2 also had been used in CSF vaccine research (8,29) and increase CSFV-specific immune response. So, we combined this information with our previous research and further constructed CSF-vectored vaccine based on human adenovirus coexpressing the CSFV E0, E2, and porcine IL2 genes (rAdV-E0-E2-IL2). Three expressed genes had been linked by foot-and-mouth disease virus 2A (FMDV 2A) sequentially. FMDV 2A or 2A-like sequences have become an alternative method and had been devised to coexpress multiple proteins under the control of a single promoter. 2A mediates a cotranslational “ribosome skipping,” which allows translation of multiple proteins from a single mRNA and cleaves at its own C terminus between the last two amino acids during protein translation. Compared to internal ribosome entry site, the other most common coexpressing approach, 2A is particularly short and the activity of each protein expressed was completely independent (4,5,7,16,33). To date, many “2A-like” sequences have been used successfully and also were an effective strategy to express multiple glycoprotein genes in adenoviruses (27). Our results showed that three proteins could be proved to express correctly in vitro and could be separated during posttranslational self-cleavage of the 2A peptide linker.
The rAdV-E0-E2-IL2 with 108 TCID50 was intramuscularly administered twice to CSFV-free pigs at 21-day intervals. No adverse clinical reactions were observed in any of the pigs after the vaccination. Neither CSFV RNA nor pathological changes were detected in the tissues after the CSFV challenge. The pigs immunized with rAdV-E0-E2-IL2 could not detect any virus after lethal challenge, which indicated that the vaccine could effectively prevent virus shedding. Pigs immunized with rAdV-E0-E2-IL2 were completely protected against lethal challenge. There were some differences between rAdV-E0-E2-IL2 and rAdV-E0-E2 or rAdV-E2-IL2, although the three recombinant viruses provided complete protection to immunized pigs against the lethal CSFV challenge. CSFV E2-specific antibodies titer of rAdV-E0-E2-IL2 were markedly increased faster and higher than rAdV-E0-E2 or rAdV-E2-IL2 after booster immunization. The differences in CSFV E2-specific antibodies were statistically unremarkable, however, the cellular immunities of rAdV-E0-E2-IL2 were superior to that of rAdV-E0-E2 or rAdV-E2-IL2. Whatever the SI, swine IFN-γ content, or relation IFN-γ mRNA gene expression, significant differences were observed between the rAdV-E0-E2-IL2 and rAdV-E0-E2 groups, or rAdV-E0-E2-IL2 and rAdV-E0-E2 groups, whereas no significant difference was observed between the rAdV-E0-E2-IL2 and CSFV C strain groups (p > 0.05). These results demonstrated that the rAdV-E0-E2-IL2 was superior to rAdV-E2-IL2 in cell-mediated immune response. Based on the promising results of the above, rAdV-E0-E2-IL2 are still needed to comprehensively evaluate, such as, whether a single administering of the vaccines could protect pigs from CSFV challenge, what were the optimal dosage, and schedule for immunization. So, considering the good immunization protection and the advantage of DIVA, rAdV-E0-E2-IL2 can be used as an alternative to the existing DIVA vaccine in controlling CSF and eradicating epidemic areas.
In summary, we introduced the CSFV E0, E2, and porcine IL2 into vectored vaccine based on HAdV-5 (rAdV-E0-E2-IL2) by coexpressing trigene linked by FMDV 2A for the first time. The vaccine induced strong humoral and cellular responses that led to complete protection from lethal CSFV challenge. The rAdV-E0-E2-IL2 is a promising, efficient, and safe marker vaccine candidate against CSFV.
Footnotes
Acknowledgments
This study was supported by the key scientific and technological innovation team project of Shaanxi Province (No. 2013KCT-28) and scientific and technological coordination, innovation, and plan projects of Shaanxi Province: genetic engineering vaccine research and development of major pig disease (No. 2014KTCQ02-02). The authors appreciate the help of staffs from the Yangling Lvfang Bio-engineering Co. Ltd. during pig managements and sample collection.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.