
Abstract
Respiratory syncytial virus (RSV) infection is a serious health problem in young children, immunocompromised patients, and the elderly. The development of novel prevention strategies, such as a vaccine to RSV, is a high priority. One strategy is to design a peptide-based vaccine that activates appropriate CD8+ T-cell responses. However, this approach is limited by the low number of RSV peptide epitopes defined to date that activate CD8+ T cells. We aimed to identify peptide epitopes that are presented by common human leukocyte antigen types (HLA-A*01, -A*02, and -B*07). We identify one novel HLA-A*02-restricted and two novel HLA-A*01-restricted peptide epitopes from RSV polymerase. Peptide-HLA multimer staining of specific T cells from healthy donor peripheral blood mononuclear cell, the memory phenotype of such peptide-specific T cells ex vivo, and functional IFNγ responses in short-term stimulation assays suggest that these peptides are recognized during RSV infection. Such peptides are candidates for inclusion into a peptide-based RSV vaccine designed to stimulate defined CD8+ T-cell responses.
Introduction
T
RSV vaccine development has been complicated by multiple factors (1,10,23, 28). First, natural infection with RSV does not induce durable protective immunity. Although antibody can protect, patients with the highest levels of neutralizing antibody are not always protected from infection, and some patients with low antibody levels are resistant (5,14,28). These data suggest that antibody-independent immunity may play an important role for protection. In support of this, children with primary deficiencies affecting T-cell function suffer more severe infection with RSV (15), adoptive transfer of memory T cells can clear RSV in mice (3,30), and fatal infantile RSV infection is characterized by a lack of T cells in the lung (40). A vaccine that elicits T-cell responses may provide long-term protective immunity (29) and add to the multiple approaches required to vaccinate all RSV target populations (1).
A second complicating factor for vaccine development is immunopathology caused by RSV infection, as well as by early vaccines. In young children, RSV can cause bronchiolitis, characterized by vigorous inflammatory responses, resulting in edema, cellular infiltration, and tissue necrosis (28). A formalin-inactivated whole virus vaccine not only failed to protect against infection but also led to enhanced disease severity following subsequent natural RSV infection in young children (20). Although T cells play an important role in viral clearance, these cells also contribute to immunopathology in mouse models (3,11,30,38). A safe and effective vaccine should activate T cells that can control the virus, but limit activation of T cells that cause severe immunopathology.
One approach to selectively activate appropriate T-cell responses to RSV, which so far has been relatively underexplored, is the development of a peptide-based RSV vaccine. Vaccine-elicited CD8+ T cells specific for an immunodominant epitope in susceptible BALB/c mice reportedly protect against both RSV infection and immunopathology (21). A detailed mechanistic analysis using major histocompatibility complex congenic susceptible and resistant mice suggests that broad high-avidity CD8+ T-cell responses promote viral clearance, whereas narrow low-avidity responses contribute to immunopathology (19). Last, in human leukocyte antigen (HLA)-A*02 transgenic mice, vaccination with some defined RSV peptide epitopes selectively induce CD8+ T cells that secrete interferon-γ, without increasing eosinophil infiltration in the lungs, and protect against RSV infection (37). Taken together, these studies suggest that defined peptide epitopes can induce protective CD8+ T-cell responses and such epitopes could be used in a safe RSV vaccine. In this study, we aimed to identify novel RSV peptide epitopes restricted by the common HLA-A*01, -A*02, and -B*07 types for use in a peptide-based vaccine.
Materials and Methods
HLA-A*01:01, HLA-A*02:01 (from Elizabeth Jaffee, Johns Hopkins University, MD), and HLA-B*07:02 constructs (GeneCust) were cloned into pMXsIP (BioCat) and expressed by retroviral transduction in K15 cells (K562 expressing membrane-bound interleukin-15, CD137 ligand, and green fluorescent protein (GFP) kindly provided by Dario Campana, St Jude Children's Research Hospital, Memphis, TN) (7). Expression was verified by staining transduced cells with pan-HLA class I antibody (W6/32; eBiosciences).
Peptide-HLA multimers were generated as previously described (12,13). HLA-A*01:01, HLA-A*02:01, and HLA-B*07:02 multimer constructs encoding a C-terminal twin-strep-tag (IBA) and sequences for control human p68 RNA helicase YLLPAIVHI, adenoviral LTDLGQNLLY, and VPATGRTLVL peptides (13), or relevant RSV peptides, flanked by Xho1 and BamH1 restriction sites, in pMXsIP were retrovirally transduced into J558L cells, and supernatants containing peptide-HLA multimers were harvested.
To determine whether peptides fold on their relevant HLA molecule, 1.5 mL supernatant from peptide-HLA multimer-transduced J558L cells was incubated overnight with 50 μL protein A/G-sepharose (Thermo Scientific). Beads were washed with PBS, boiled for 5 min in 50 μL SDS-PAGE loading buffer (125 mM Tris-HCl pH 6.8, 2% SDS, 13% glycerol, 5% 2-mercaptoethanol), and 20 μL resulting supernatant was resolved by SDS-PAGE. Resolved proteins were stained with Coomassie blue (Sigma). Peptide-HLA multimers were identified according to their molecular mass of 98 kDa, as well as by Western blotting using an antibody (Dianova) specific for mouse IgG1 in the fusion proteins (data not shown). Stabilization of HLA by peptides was additionally assessed by cell surface expression of HLA covalently linked to relevant peptides as described previously (26). For this purpose, the peptide-HLA multimer constructs described above were fused with the transmembrane and intracellular domains of HLA-A*02:01 in pMXsIP using the common Sbf1 site in the HLA alpha3 domain, and the resulting constructs were retrovirally expressed on K15 cells. Cell surface expression was assessed by staining with pan-HLA class I antibody (W6/32; eBiosciences) and quantified as the difference between the median fluorescence intensities of staining of cells transduced with peptide-HLA constructs and control-untransduced cells (ΔMFI).
Buffy coats were obtained from the Blood Bank of the University Hospital Tübingen with approval from the institutional ethics board. Peripheral blood mononuclear cells (PBMC) from healthy HLA-A*01/A*02/B*07-positive donors (typed using low-resolution PCR-based HLA typing) were isolated through Ficoll and incubated at a density of 107 cells/mL in DMEM containing 10% fetal calf serum, 2 ng/mL interleukin-2 (PeproTech), and 1 μg/mL relevant peptides (synthesized by GeneCust). After 7 days, cells were stimulated with irradiated (100 Gy) K15 HLA-A*01:01/A*02:01/B*07:02 cells preloaded with 1 μg/mL relevant peptide or with irradiated K15 cells expressing HLA covalently linked to relevant peptides generated as described above. After 2 weeks of expansion on K15 feeder cells, peptide-HLA multimers concentrated from 7.5 mL transduced J558L cell supernatant to 100 μL per staining using 100 kDa cutoff centricons (Merck-Millipore) and preincubated with 5 μL streptactin-phycoerythrin (IBA) were used to stain cells in combination with CD3-allophycocyanin antibody (BD Biosciences) before analysis on a fluorescence-activated cell sorter Calibur (BD Biosciences). Cells were gated using a forward scatter/side scatter living lymphocyte gate, followed by gating on CD3+ cells, and analyzed with CellQuest software.
Memory T-cell phenotype determination was performed as previously described (13). CD45RO+ cells from healthy donor PBMC were purified ex vivo using CD45RO microbeads (Miltenyi Biotech) and stimulated for 2 weeks with K15 cells expressing HLA-A*01/A*02/B*07 covalently linked to relevant RSV peptides as described above. Cells were stained with peptide-HLA multimers precoupled to streptactin-phycoerythrin (IBA), together with CD45RO-allophycocyanin and CD28-FITC (BD Biosciences).
Cultured IFNγ ELIspot assays were performed as described by Geyeregger et al. (8). Briefly, thawed PBMC were incubated overnight and then stimulated with 1 μg/mL relevant peptide in individual wells. IL-2 at a final concentration of 2 ng/mL was added on day 2, 5, 7, 9, and 11. PBMC were harvested after 12 days and seeded at 5 × 105 cells/well in DMEM (10% fetal calf serum) onto nitrocellulose plates (Immunospot M200; Merck Millipore). Stimulation was for 20 h with 1 μg/mL relevant RSV peptide, 1 μg/mL human p68 RNA helicase peptide YLLPAIVHI as a negative control, 1 μg/mL HLA-A*01-restricted LTDLGQNLLY or HLA-A*02-restricted LLDQLIEEV adenoviral peptides, HLA-A*02-restricted NLVPMVATV HCMV peptide, and 10 μg/mL PHA (Sigma) as positive controls. IFNγ secretion was detected using an ELISpot kit (Mabtech), and spot numbers were counted by an ImmunoSpot Series 6 ELISPOT Reader (CTL). Peptide responses were considered positive if the mean number of spots per well was at least 20 and more than thrice the mean number of spots in the negative control. Samples in which the negative control exceeded 120 spots per well were excluded.
Results
We aimed to identify novel RSV peptide epitopes suitable for use in a peptide-based vaccine. We chose to work with HLA-A*01, -A*02, and -B*07 given that these HLA types are common in Europe and their peptide-binding motifs are well defined (25,35). Using peptide epitope precision software (NetMHC 3.4) (25), we chose 14 peptides with high-predicted affinity for their respective HLA allotypes (Table 1). All of these peptides have been shown by Rock et al. (31) to stabilize their respective HLA molecules, although it is not clear if these peptides are epitopes. We generated peptide-HLA multimers and analyzed these by SDS-PAGE to confirm whether the peptides fold on and hence stabilize their respective HLA allotypes (Fig. 1A). As positive controls, we used the HLA-A*02-restricted self-peptide YLLPAIVHI and the adenoviral peptides LTDLGQNLLY and VPATGRTLVL (HLA-A*01 and HLA-B*07 restricted, respectively), which all stabilize their respective HLA molecules (13). With the exception of FTGLIGMLY, which in contrast to Rock et al. (31) did not stabilize HLA-A*01, all RSV peptides folded to varying degrees in HLA multimers. Notably, HLA-A*01-restricted FSECNALGSY and ISDYFHNTY, HLA-A*02-restricted YLSELLNSL, and all HLA-B*07-restricted peptides folded comparably well to positive control peptides in this qualitative assay (Fig. 1A).
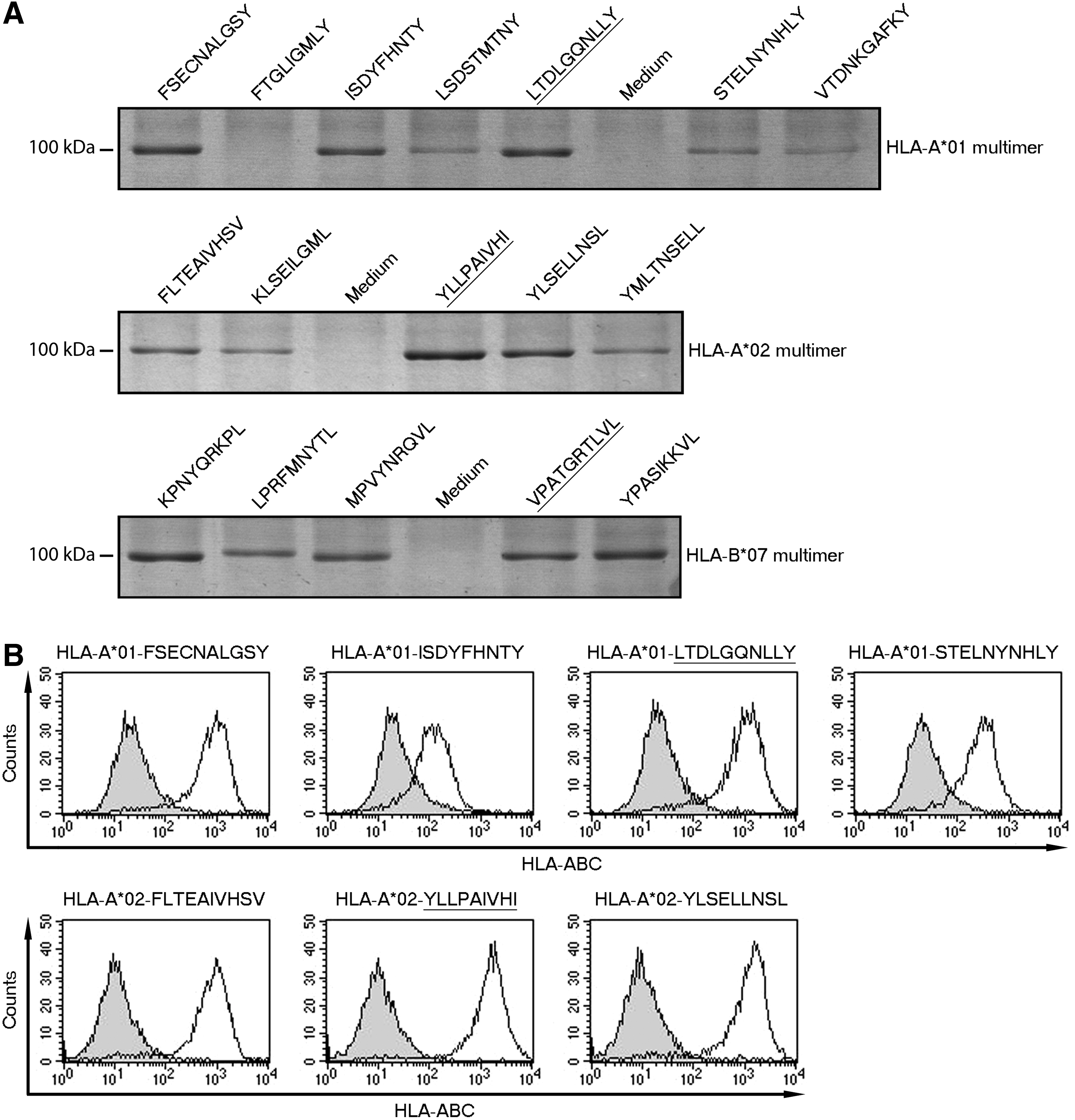
Stabilization of HLA by predicted peptide epitopes from RSV.
HLA, human leukocyte antigen; NA, not analyzed.
We also analyzed folding of peptides on HLA in a quantitative manner, using K15 cells expressing HLA covalently linked to selected peptides. High expression of HLA, determined by flow cytometric staining with a pan-HLA class1 antibody, was regarded as indicating efficient binding and stabilization of the relevant HLA molecule (Fig. 1B). In these assays, HLA-A*01-restricted FSECNALGSY folded comparably well to the positive control LTDLGQNLLY (ΔMFI 778 and ΔMFI 944, respectively). However, HLA-A*01-restricted ISDYFHNTY and STELNYNHLY stabilized HLA-A*01 less efficiently (ΔMFI 89 and ΔMFI 244, respectively). Both HLA-A*02-restricted peptides YLSELLNSL and FLTEAIVHSV bound comparably well to HLA-A*02 as the positive control YLLPAIVHI (ΔMFI 1220, ΔMFI 753, and ΔMFI 1423, respectively, Fig. 1B). A number of candidate RSV peptide epitopes therefore bind and stabilize their respective HLA molecules.
We selected eight peptide-HLA multimers, which folded adequately for their use in flow cytometry, to detect RSV-specific T cells. Given the low frequency of RSV-specific T cells in vivo in healthy donors (16) and relatively high background staining with our multimers (13), we expanded specific T cells before detection. PBMC from HLA-A*01/A*02/B*07-positive healthy donors were stimulated with peptide and interleukin-2 and further expanded using K15 HLA-A*01/A*02/B*07 feeders before staining with peptide-HLA multimers. No specific T cells could be stained with any of the peptide-HLA-B*07 multimers or the YLSELLNSL-HLA-A*02 multimer from at least three independent donors each, suggesting that these peptides are either not recognized by T cells or recognized as subdominant epitopes. By contrast, all three HLA-A*01 multimers stained RSV-specific T cells, with mean responses of 5.5% ± 2.0% for FSECNALGSY, 10.7% ± 4.4% for ISDYFHNTY, and 6.4% ± 3.6% for STELNYNHLY of all CD3+ T cells (Fig. 2A, B and Supplementary Table S1; Supplementary Data are available online at
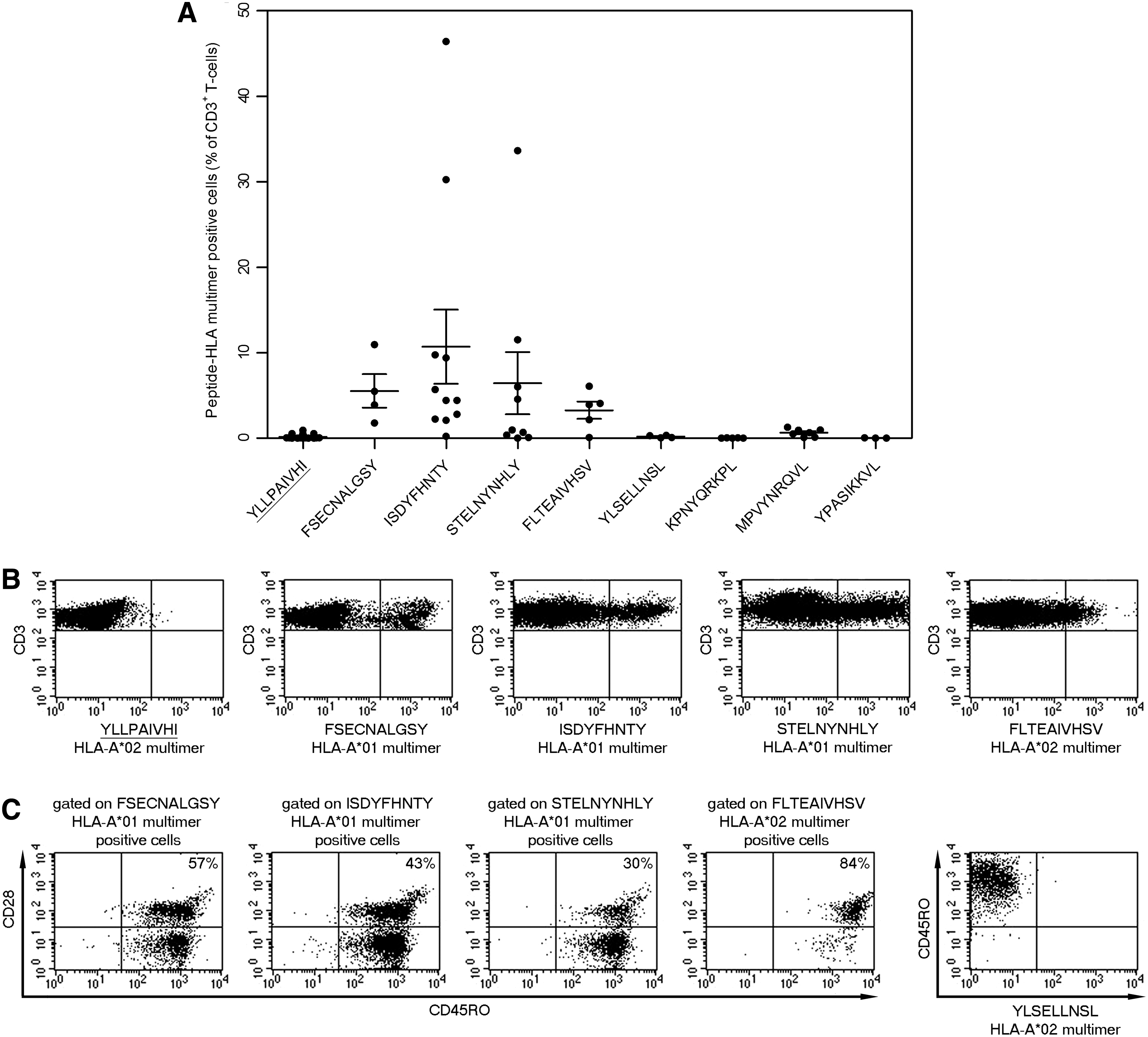
Detection and memory phenotype of RSV-specific T cells.
Given that almost all adults have been infected by RSV (22), we next measured recall T-cell responses to the candidate peptide epitopes. We first determined whether peptide-specific T cells have a memory phenotype ex vivo as previously described (13). We purified memory T cells ex vivo using CD45RO microbeads, expanded RSV-specific T cells with K15 cells expressing HLA-A*01/A*02 covalently linked to relevant RSV peptides, and stained such specific T cells with peptide-HLA multimers and memory markers (Fig. 2C and Supplementary Table S2). We detected T cells specific for all three HLA-A*01-restricted peptides, as well as the HLA-A*02-restricted peptide FLTEAIVHSV, which expressed both CD45RO and CD28, suggesting that these are either central or effector memory T cells in vivo (33,34). Surprisingly, however, in six out of nine donors, a high proportion of HLA-A*01-restricted T cells were CD28 negative, suggesting that many specific cells are differentiated effector T cells. By contrast, the majority of HLA-A*02-restricted FLTEAIVHSV-specific T-cells were CD28 positive in three out of three donors. Stimulation of memory T cells, purified directly ex vivo, with the peptide YLSELLNSL or with nonimmunogenic control peptides as previously described (13), did not result in expansion of peptide-specific T cells (Fig. 2C), thus demonstrating specificity of memory T-cell expansion with the RSV peptides FSECNALGSY, ISDYFHNTY, STELNYNHLY, and FLTEAIVHSV. These data suggest that there are memory T cells specific for all four RSV-derived peptides, and hence these peptides are epitopes recognized during RSV infection.
The differentiated phenotype of peptide-specific T cells may indicate that these cells are not able to respond to reinfection. For this reason, we performed functional assays using cultured ELISpot (Fig. 3). IFNγ-secretion responses to the HLA-A*01-restricted peptides STELNYNHLY and ISDYFHNTY were readily detectable in two of eight donors, respectively, with one donor responding to both peptides and a further two donors responding to either STELNYNHLY or ISDYFHNTY (Fig. 3A, C and Supplementary Table S3). IFNγ-secretion responses were also detected to the HLA-A*02-restricted peptide FLTEAIVHSV in two of six donors (Fig. 3A, C). Although responses to the HLA-A*01-restricted peptide FSECNALGSY were elevated in one of eight donors, such responses were not three times higher than the respective negative controls and were therefore considered negative. Responses to RSV peptides were low compared to responses to well-defined adenoviral peptide epitopes (582 ± 149 SFC for HLA-A*01-restricted LTDLGQNLLY and 440 ± 163 SFC for HLA-A*02-restricted LLDQLIEEV from the same donors, data not shown) (8). As a further control, we stimulated HCMV-seronegative donor PBMC with the immunodominant HLA-A*02-restricted peptide NLVPMVATV from HCMV (41). There were no responses to the HCMV peptide in HCMV-seronegative donor PBMC (Fig. 3B, D), indicating that there is no priming of naive T cells in this short-term assay, and hence strongly suggesting that the responses to RSV peptides are recall responses.
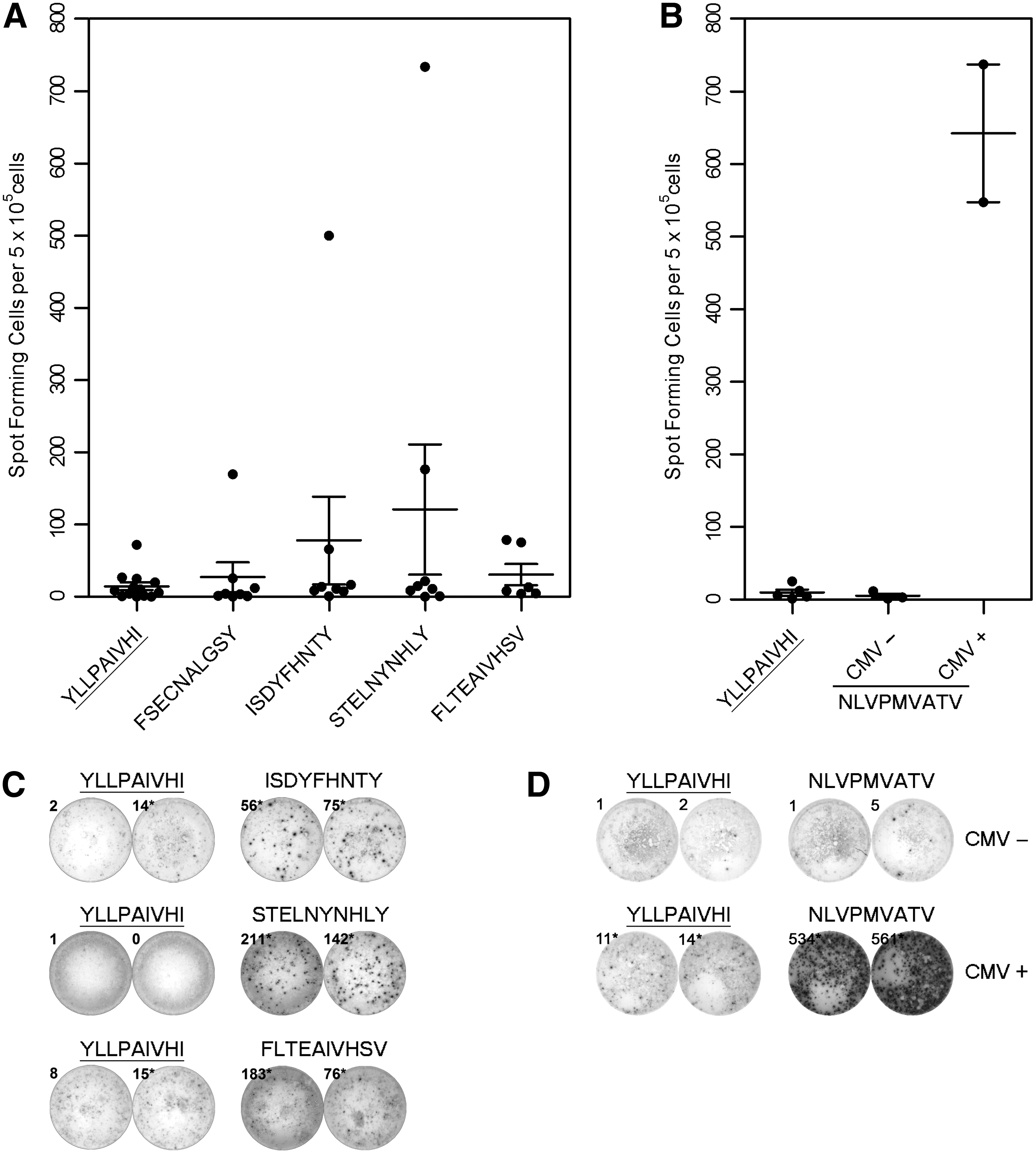
Functional analysis of RSV peptide-specific T cells.
Discussion
The immunopathology that occurs during RSV infection in young children, particularly in RSV vaccines subsequently naturally infected with RSV (20), is a major factor that has limited the development of an RSV vaccine. One way to largely circumvent such immunopathology may be to design vaccines that use short peptide epitopes to stimulate defined protective CD8+ T-cell responses (21,37) and facilitate appropriate memory responses in the lung. Given that there are relatively few well-characterized RSV epitopes described to date that stimulate CD8+ T cells (2,9,16,39), covering only seven HLA types (27), we aimed to identify and characterize more peptide epitopes that may be included into such a vaccine. We identify two novel HLA-A*01:01-restricted epitopes and one HLA-A*02:01 epitope. All of these peptide epitopes are from the RSV polymerase, and thus add to the one HLA-B*27-restricted epitope GRNEVFSNK from RSV polymerase (17), as well as to the well-described HLA-A*01-restricted epitope YLEKESIYY from RSV matrix protein (16).
In all donors, there were populations of RSV-specific memory T cells positive for both CD45RO and CD28, indicating the presence of central or effector memory T cells in vivo. However, in most donors, the majority of RSV-specific memory T cells were CD28 negative. This is in contrast to the results from Heidema et al. (16) who showed that memory T cells specific for the HLA-A*01-restricted peptide YLEKESIYY from five of five donors were uniformly CD28 positive. While these differences may reflect the different methodologies used, we have previously shown that adenovirus-specific memory T cells are uniformly CD28 positive using the same methodology as that used in this study (13). We would therefore argue that memory T cells specific for RSV are either more differentiated or differentiate more rapidly than adenovirus-specific memory T cells, thus reflecting the differentiated status of RSV-specific CD8+ T cells in the lung (4).
Despite their differentiated status, T cells specific for the peptides STELNYNHLY and ISDYFHNTY were able to respond to challenge by secreting IFNγ, as were T cells specific for FLTEAIVHSV (Fig. 3A, C). Given that we were not able to prime HCMV responses from HCMV-seronegative donors in this short-term stimulation assay (Fig. 3B, D), we argue that such responses are recall responses, and thus the peptides are recognized as epitopes during RSV infection. Surprisingly, there were no positive IFNγ recall responses to the peptide FSECNALGSY (Fig. 3A), despite positive peptide-HLA multimer staining and the presence of FSECNALGSY-specific memory T cells (Fig. 2B, C). While these results may reflect the low frequency of T cells specific for FSECNALGSY, we cannot rule out that longer term stimulation of T cells with K15 antigen-presenting cells for peptide-HLA multimer staining may have primed naive T cells. Based on the negative IFNγ recall assays, we cannot confirm that FSECNALGSY is recognized as an epitope. Surprisingly, RSV-specific IFNγ responses were even lower than the previously described low responses to adenoviral peptide epitopes in healthy donors (data not shown) (8). It remains to be tested whether a therapeutic RSV peptide-based vaccine can boost memory T-cell responses to RSV and whether these confer protection.
Taken together, we identify one novel HLA-A*02-restricted and two novel HLA-A*01-restricted peptide epitopes from the RSV polymerase. These results lay the groundwork for the use of novel peptide epitopes in a peptide-based RSV vaccine designed to stimulate defined CD8+ T-cell responses.
Footnotes
Acknowledgments
This study was supported by the Interdisziplinäres Zentrum für Klinische Forschung Tübingen (DB, KMD). Use of human blood products was approved by the ethics commission of the Medical Faculty, Universitätsklinikum Tübingen. We thank the staff of the Blood Bank (Tübingen, Germany), Dario Campana (St Jude Children's Research Hospital, Memphis, TN), Tim Greten (NIH, Bethesda, MD), Klaus Hamprecht (Tübingen, Germany), Elizabeth Jaffee (Johns Hopkins University, MD), Peter Lang (Tübingen, Germany), and Wolfgang Schamel (Freiburg, Germany) for generously providing reagents.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.