
Abstract
Recent information demonstrated that BK virus reactivation is a dominant complication after kidney transplantation, which occurs because of immunosuppression. BK virus reactivation is the main reason of transplanted kidney losing. Immune response against BK virus is the major inhibitor of the virus reactivation. Therefore, improving our knowledge regarding the main parameters that fight against BK viruses can shed light on to direct new treatment strategies to suppress BK infection. Innate immunity consists of numerous cell systems and also soluble molecules, which not only suppress virus replication, but also activate adaptive immunity to eradicate the infection. Additionally, it appears that immune responses against reactivated BK virus are the main reasons for induction of BK virus-associated nephropathy (BKAN). Thus, improving our knowledge regarding the parameters and detailed mechanisms of innate immunity and also the status of innate immunity of the patients with BK virus reactivation and its complications can introduce new prospective strategies to either prevent or as therapy of the complication. Therefore, this review was aimed to collate the most recent data regarding the roles played by innate immunity against BK virus and also the status of innate immunity in the patients with reactivation BK virus and BKAN.
Introduction
BK
Extensively, the recurrence of BK virus infection is observed after allograft renal transplantation (28). As the virus induces nephritis and also ureteral stenosis in 1–10% of the patients under allograft renal transplantation (60), so, it has been considered as a major complication after renal transplantation and often results in graft rejection (40). Due to the fact that immunosuppression leads to reactivation of the BK virus, hence, it can be hypothesized that the pressure from immune responses can be considered as an important factor for suppression of BK activation (26).
Additionally, as mentioned in previous sentences, the reactivation of BK occurs in 1–10% of the transplanted patients (60), hence, it may also be hypothesized that the differences in genetic and immunological parameters can play key roles in the determination of infection outcome. Thus, improving our knowledge regarding the details of immune responses against BK virus can direct new treatment strategies to inhibit BK virus-related nephritis.
Innate immunity is an important arm of immune system, which plays critical roles in eradication and suppression of viruses (44). Several cell systems and also cellular and soluble molecules in innate immunity significantly participate in defense against viruses. Based on the important roles played by immune responses, especially innate immunity, in suppression of BK viruses and induction of BK virus-associated nephropathy (BKAN) and also the urgent necessities of new strategies to control BK infection in transplanted patients, it seems that improving our knowledge regarding the key parameters of innate immunity against BK and involving in the pathogenesis of BKAN can direct new strategies to treat BK virus reactivation.
Accordingly, the main aim of this review article was to describe recent information regarding the cellular and molecular mechanisms that are applied by innate immunity against BK virus and involved in the induction of BKAN and also the status of innate immunity in the patients with reactivated BK virus.
Evidence Acquisition
To collate the published information regarding the roles played by innate immunity during BK reactivation, a complete searching was performed using medical terms for BK virus, transplantation, innate immunity, nephritis, cytokines, inflammasomes, Toll-like receptors (TLRs), dendritic cells (DCs), natural killer (NK) cells, macrophages, neutrophils, basophils, eosinophils, mast cells, complement system, ficolins, lectins, pentraxins, melanoma differentiation-associated gene 5 (MDA5), retinoic acid-inducible gene I (RIG-1), stimulator of interferon (IFN) genes, (STING) and transcription factors (nuclear factor kappa light-chain-enhancer [NF-κB], activator protein-1 [AP-1], and IFN regulatory factors [IRFs]) from the famous international scientific databases such as PubMed, Medline, Web of Science, Web of Knowledge, Scopus, ProQuest, and Google Scholar. To improve our investigation, some advanced search using Boolean operators, controlled vocabulary (MeSH), truncations, limits, and field searching were performed.
BK Virus
BK virus is a human polyomavirus, which is prevalent in several ethnic populations in latent form (79). The virus can be reactivated following immunosuppression and leads to some complications such as nephropathy (79). The genome of BK virus consists of three major genetic regions, including early, late, and noncoding regions (56). The latest region is entitled NCCR and two different forms of the BK virus is defined based on the DNA sequences of the NCCR, archetype and rearranged variants (58). Archetype is separated from urine of either healthy people or diseased patients, but rearranged variants can most often be isolated from the serum of BK reactivated patients (57). BK virus can be detected in the 5–10% of immunocompetent adults, which is a sign of asymptomatic reactivation of the virus (40).
It seems that archetype variant is the transmissible form of BK virus because it is separated from healthy individuals (15). Immune system is able to suppress BK virus reactivation and keep it in latent form, so, immunosuppression is associated with reactivation of BK virus that is seen in renal and bone marrow transplant (BMT) (59,73,75). BK virus reactivation during renal and BMT leads to nephropathy and hemorrhagic cystitis, respectively (34,59,73,75).
BK virus can be transmitted commonly through a respiratory route, but transmission through other routes, including fecal/urine, oral, blood transfusion, and transplantation have been reported by investigators (13). In several cases, primary infection is asymptomatic, but a mild respiratory illness may be manifested in some cases (13). The infection then disseminates to the urinary tract and replicates in the epithelial cells, kidney, and bladder (25).
In spite of all that, there are some publications which reported that other cell system like immune cells can be infected and the virus can be proliferated in the cells (63). In healthy people, BK virus can be replicated in low rate to shed the urine (25). BK virus can be divided into four genotypes rather than variants in the NCCR, termed genotypes I, II, III, and IV (47). The genotypes are defined according to the sequence variations in VP1 (7). The genotypes correspond to four different serotypes, BK, SB, As, and IV, respectively (7).
The genotype I is the most predominate genotype and its subgroup I/b2 is prevalent in the European and American populations, while subgroup I/c–2 is prevalent in the Asian population (11).
BK Virus Infection and Nephropathy
BKAN is an inflammatory interstitial nephropathy, which occurs 10–13 months after renal transplantation (41). However, development of new immunosuppressive drugs leads to decreased acute renal rejection rates, but results in increasing the incidence of BKAN (41). More than 10% of renal transplants are associated with BKAN and 90% of BKAN patients lose their graft (64). Some parameters increase the risk of BKAN and are as follows: older age, male gender, a history of BKAN, seronegative recipient, and seropositive donors (5,65). Entrance and replication of BK virus in epithelial, renal, and bladder cells result in cell dysfunction and progression of renal transplant loss (65).
Innate Immunity: Cell Systems, Extra/Intracellular Receptors, Intracellular Signaling, and Soluble Molecules
Innate immunity is the first line of immune responses against viral infection (16). Innate immunity consists of several cell systems and soluble molecules. Then main innate immunity cells are monocytes (macrophages in the tissues), DCs, NK cells, eosinophils, basophils, neutrophils, and mast cells (12). The soluble molecules in innate immunity are categorized into several families, including complementary system, pentraxins, lectins, ficolins, and some nonimmunologic molecules participating in inhibitory functions against microbes such as transferin and lactoferin (77).
The cells of innate immunity perform their immunological functions against virus through (i) phagocytosis of opsonized and non-opsonized virus, by macrophages, DCs, and neutrophils; (ii) antibody-dependent cellular cytotoxicity by NK cells, basophils, eosinophils, and mast cells; (iii) presenting viral antigens in adaptive immunity by major histocompatibility complex (MHCs), by macrophages and DCs (12,22). The innate immunity cells recognize viral pathogen-associated molecular patterns (PAMPs) through their extra/intracellular receptors (6). TLRs, c-type lectins, scavenger receptors, formyl-methionine receptors, and inflammasomes are the main extra/intracellular receptors in innate immunity (42,49,62).
There are some intracellular receptors in the cytoplasm of innate immune and nonimmune cells, which participate in viral PAMPs detection that are not in the categories, like MDA5, RIG-1, STING, and so on (45). Following recognition of BK virus by the extra/intracellular receptors, some known intracellular signaling pathways, such as mitogen-activated protein kinase (MAPK), TIR domain-containing adapter-inducing IFN-β (TRIF), and myeloid differentiation primary response gene 88 (MYD88)-dependent pathways, will be activated and subsequently lead to the activation of proinflammatory transcription factors like NF-κB, IRF3, IRF7, and AP-1 (72). The transcription factors will be translocated to the nucleus and transcript from proinflammatory molecules, including cytokines and costimulatory molecules, as well as homing and addressing molecules (72).
We will discuss the role and status of the innate immune cells, their receptors, and innate immunity soluble molecules in BK virus infection and also during BKAN in the next section.
Innate Immunity and BK Virus
Innate immunity cells and BK virus
As described in the previous section, innate immunity cells are the main arm of immune cells, which play key roles against viral infections. NK cells are the important immune cells which recognize viral infected cells through downregulation of class I MHCs, through inhibitory receptor, and upregulation of activator molecules like UL16-binding (ULB) protein, MHC class I polypeptide-related sequence A (MIC-A), and MHC class I polypeptide-related sequence B (MIC-B), through activating receptors, on the infected cells and kill or induce apoptosis in viral infected cells following activation (71).
According to the critical roles played by NK cells against viral infections, it has been proposed that the cells may significantly participate in the suppression of BK viral infections. Accordingly, a study by Trydzenskaya et al., revealed that the patients with BKAN carry low numbers of killer-cell immunoglobulin-like receptors (KIRs) as activating receptors, compared with the control group (kidney recipients with stable function and no BKAN) (78). They have also showed that KIR3DS1 was significantly decreased on the NK cells of patients with BKAN and also BK-reactivated patients when compared to the controls (78). Based on the study by Trydzenskaya et al., it can be concluded that NK cells play crucial roles in the recognition of BK viral infection and suppress replication of the virus.
It has been documented that NKG2D is an activating receptor on the NK cells that recognize a set of molecules, including ULB, MIC-A, and MIC-B, which are upregulated on the viral infected and tumor cells as well as under stress cells (71). An interesting study has been performed by Bauman et al., which demonstrated that the 3p′ miRNAs of BK virus through binding to the 3′UTR of UL16-binding protein 3 (ULBP3) leads to downregulation of ULBP3 (10). They have been revealed that downregulation of ULBP3 is not related to its mRNA levels, because its mRNAs did not differ among miRNAs transfected and nontransfected cells, but its protein levels were decreased after transfection. It was suggested that the 3p′ miRNAs of BK virus target translation of ULBP3 (10). Thus, it may be suggested that BK virus inhibits NK cells activation through suppressing the NKG2D/ULBP3 interaction.
In parallel with the results, another study showed that NK cells are the main cells in defense against BK virus (3). The mechanisms used by BK virus to suppress NK cell activation has been described in Figure 1. Collectively, according to the fact that BK virus targets the ligands of NK cells activating receptors on infected cells, it seems that NK cells play key roles in the healthy people to suppress BK virus through killing the source of the infection. Additionally, there are some documents which noted that NK cell activities against BK virus are associated with graft rejection. For instance, increased activities of NK cells in BKAN and BK viremia are associated with acute renal rejection (52).
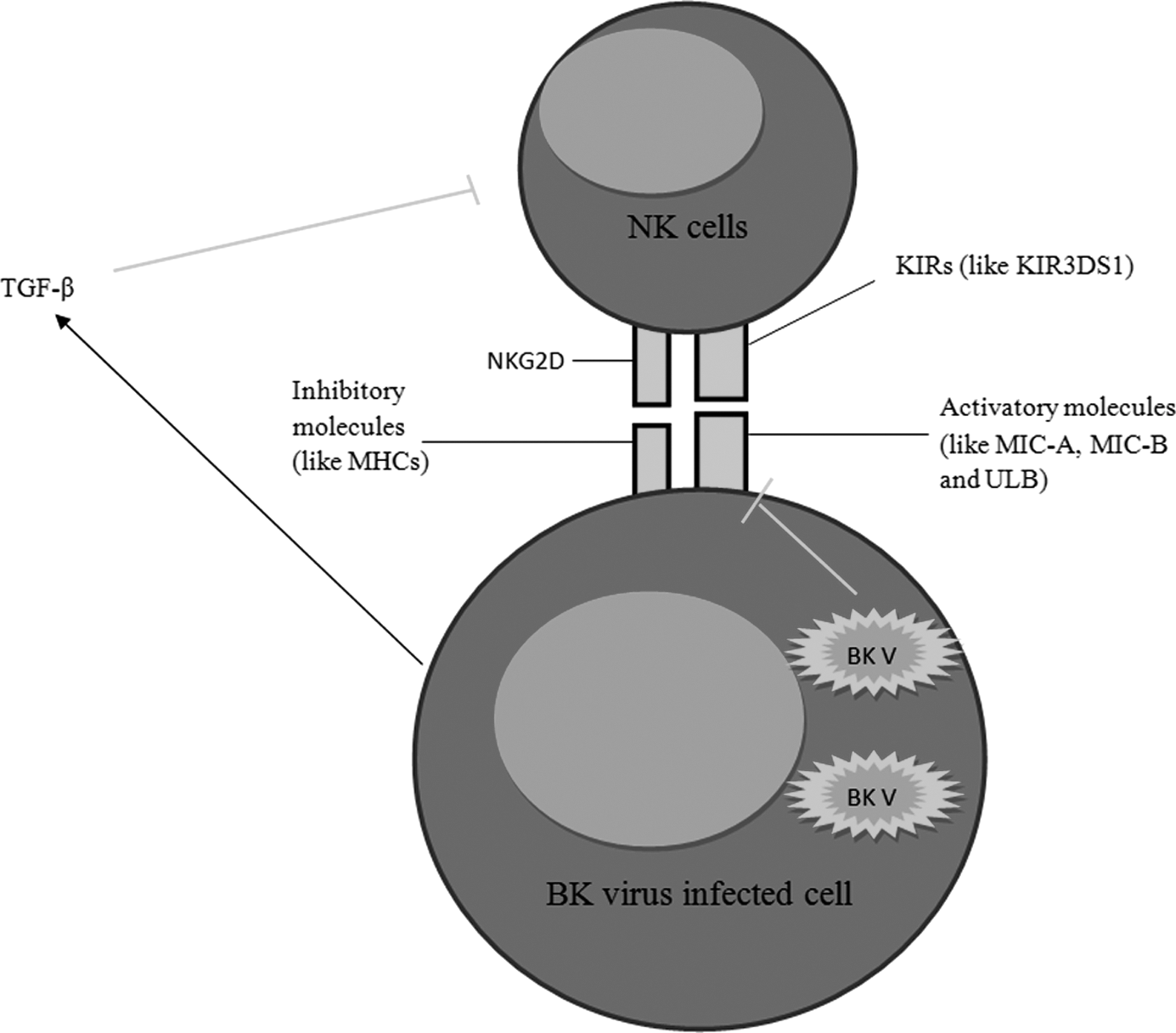
The mechanisms used by BK virus to suppress natural killer (NK) cells. As illustrated in the figure, BK virus suppresses the expression of UL16-binding protein 3 (ULBP3) and also induce the infected cells to produce tumor growth factor (TGF-β) to suppress NK cell functions.
It may be hypothesized that NK cells target the infected cells following the replication of BK virus at immunodeficiency conditions and, hence, induce graft rejection through either direct lysis of the infected target cells and losing their functions or release the allograft antigens through lysis of the infected cells and activation of adaptive immune system. Finally, according to studies, it appears that NK cells are the important parameters to suppress replication of BK virus, but after reactivation of BK virus it may induce severe complications.
It appears that macrophages and DCs not only contribute in defense against BK virus directly through phagocytosis of the virus particle, but also can modulate the disease in an indirect manner by present BK virus antigens and also production of various ranges of cytokines which are discussed in the next sections (29). Accordingly, a study by analysis of 1,189 urine sediments from 174 renal allograft recipients reported that the number of CD14-positive cells (macrophages and DCs) and the main attractor of the cells, monocyte chemoattractant protein-1 (MCP-1), were significantly associated with intragraft tubulointerstitial inflammation in the patients with BKAN (14).
DCs are the most important priming T cells, which present viral antigens to T cells through both class II (conventional) and class I (cross presentation) MHCs (53). DCs can migrate from infection site to the nearest peripheral lymphoid nodes and prime naive T cells. According to the important roles played by DCs, several studies have used DCs as vehicles for direct and indirect immunotherapy against viruses, including BK virus in immunodeficient patients (35,50). Major capsid protein VP1, derived from BK virus, induced mild DC maturation (32). Another study demonstrated that peripheral blood deficiency in renal transplanted patients is significantly associated with BK virus reactivation and is a predictor for the development of BKAN (81).
Neutrophils like macrophages and DCs are capable of phagocytosis of the virions (33). Geetha et al. reported that BK virus reactivation is seen more in the renal transplanted patients with anti-neutrophil cytoplasmic antibody-associated vasculitis, who suffered from neutrophil dysfunctions than renal transplanted patients with normal functions of neutrophils (33). It has also been reported that BK virus infection is inhibited by α-defensin human neutrophil protein 1 (HNP1) and human α-defensin 5 (HD5) through targeting the viral lifecycle. Additionally, human α-defensins also suppress BK virus infection through the virus aggregating and blocking their binding to host cells (27).
Nevertheless, some investigators believe that the existence of neutrophil component in the serum cannot be considered as a predictor for BKAN and BK virus reactivation following renal transplantation. For example, Rau et al. revealed that serum levels of neutrophil gelatinase-associated lipocalin are associated with neither BK nephropathy nor BK virus infection (66). Based on the results, it seems that the phagocytic cells, such as macrophages, DCs, and neutrophils, are the main arms of innate immunity to defense against BK virus, so the virus targets the cells to escape from immunosurveillance. However, the roles of macrophages, DCs, and neutrophils in the induction of BKAN are controversial and more studies are needed to complete our knowledge.
Eosinophils are another cell system in innate immunity, which prepare immunity against microbes especially parasites (68). The roles of eosinophils in defense against BK virus and their functions in the BKAN were not studied well. There is a study in this field that has been performed by Yamanaka et al. (84). They have reported that the presence of tubulitis in association with eosinophil infiltration is the main pathological feature of BKAN (84).
Accordingly, it appears that like other pathological features of eosinophils, its dangerous enzymes may participate in the induction of some pathological events during BKAN. Based on our investigations in research databases, there are no studies regarding the roles played by basophils and mast cells in immunity against BK virus and their roles in BKAN, hence, it appears that more studies may improve our knowledge regarding the roles played by eosinophils, basophils, and mast cells in innate immunity responses against BK virus infection and its complications. The roles played by innate immune cells in induction of BKAN and also reactivation of BK virus have been described in Figure 2.
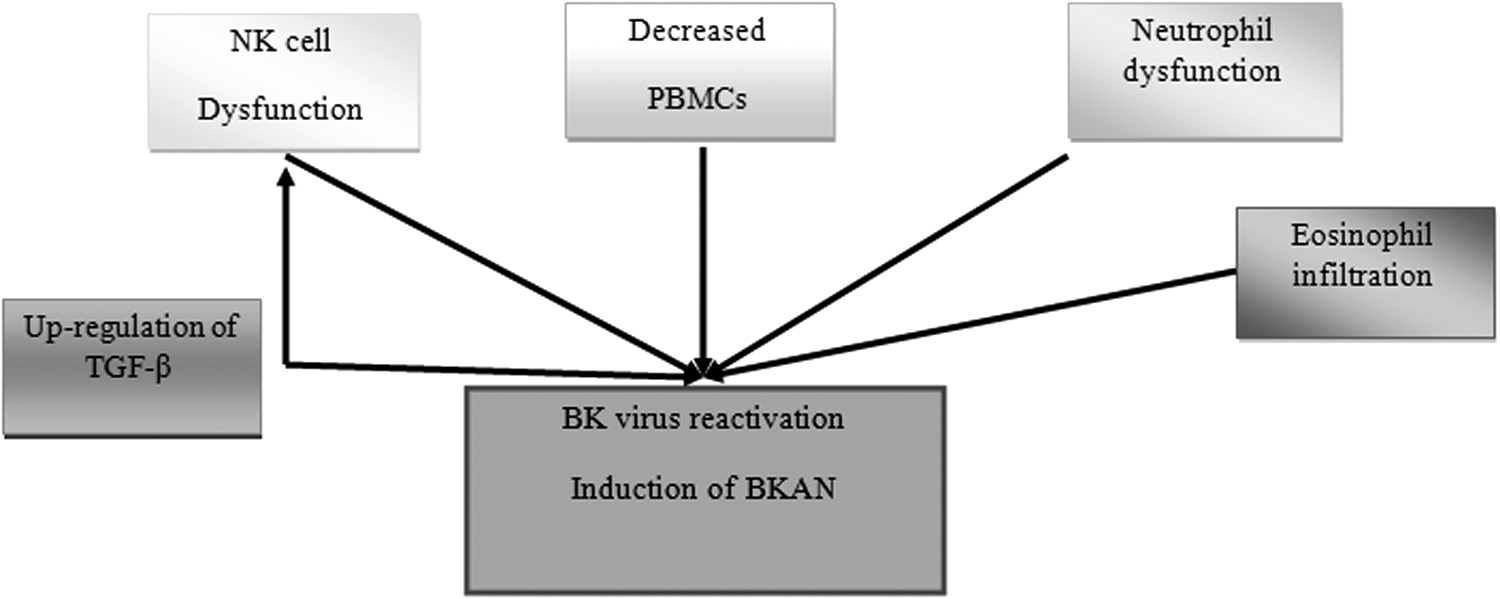
The roles of innate immunity cells on reactivation of BK virus as well as induction of BK virus-associated nephropathy (BKAN). As illustrated in the figure, the NK cell and neutrophil dysfunctions and also decreased peripheral blood mononuclear cells (PBMCs) may be the main reasons of BK virus reactivation. Eosinophil infiltration is a plausible reason of induction of BKAN.
Innate immunity sensors and BK virus
There are several viral sensors in innate immunity and also nonimmunity cells, which recognize BK viruses, and activate some intracellular signaling pathways. TLRs, inflammasomes, and viral nucleic acid sensors, like STING, MDA5, and RIG-1, are the main intracellular sensors (38,85). Among them, previous studies have evaluated the roles played by TLR3, MDA5, and RIG-1 in defense against BK virus and involved in BKAN only. Heutinck et al. reported that human tubular epithelial cells express TLR3, MDA5, and RIG-1, which are the sensors of dsRNA and inducers of immunity against viruses as well as inflammation (38). A study by Heutinck et al. revealed that the intracellular viral sensors are upregulated in the BK virus-infected human tubular epithelial cells (39). Another study demonstrated that activation of innate immunity mechanism is a main mechanism involved in BKAN (67).
Interestingly, the study demonstrated that mRNA levels of TLR3, but not RIG-1 were increased in BKAN (67). NF-κB and AP-1 are the transcription factors in downstream of several intracellular signaling pathways, including TLRs (44). Interestingly, it has been reported that BK virus has binding motifs for NF-κB and AP-1 in the promoter/enhancer region, which results in transcription from the BK virus early gene, the first event in viral lytic infection (17,18,36).
Accordingly, it may be concluded that TLR3 induces BKAN through either activation of inflammation or induction of viral lytic infection. As mentioned in previous sentences, the studies have evaluated the roles of TLR3, MDA5, and RIG-I in BK virus infection and the roles played by other sensors have yet to be clarified. But there is a study, which has been reported that expression levels of interleukin (IL)-1β, as the main target of inflammasome pathways, were not changed during BKAN (67). Based on the fact that inflammasome pathways lead to activation of IL-1β and have no affect on its expression (55), we are, therefore, unable to conclude that inflammasomes do not play key roles in the BKAN and more study are needed urgently.
Soluble molecules in innate immunity and BK virus
Soluble molecules are the main arms of innate immunity, which participate in viral elimination and also their complications by either directly targeting the viruses, like complementary system, or indirectly by opsonization of the particle (24). There are several studies regarding the roles of soluble molecules in the innate immunity against BK virus and induction of BKAN. Most of the studies were performed regarding the roles of complementary system.
Complementary system is activated through three pathways, including classical, alternative, and lectin-dependent pathways (30,83). In a study by Sharif et al. it has been reported that the deposition of C4b, a component of complementary system in classical and lectin-dependent pathways, is associated with BKAN in ABO system incompatible recipients (74). In another study, it was shown that diffuse tubular basement membrane C4d deposition was significantly associated with BKAN (9). Honsova et al. also confirmed the results (41). They have revealed that there is a relationship between BKAN and BK virus infection with activation of complementary system through classical and lectin-dependent pathways because a simultaneous C4d deposition was observed in the transplanted kidney and during BK virus reactivation, which led to BKAN (41).
According to the results, it appears that complementary system is an important arm of innate immunity, which participates in the induction of BKAN, may be a mechanism that is seen in super acute rejection, and results in losing the kidney following BK virus infection. It has not been demonstrated what the activator of complementary system is, specific antibody or recognition of BK virus component by lectins or C3b. Collectively, it seems that the complementary system not only disrupts the function of transplanted kidney cells, but also accelerates BKAN through induction of inflammation by complementary system anaphylatoxins like C3a and C5a that is reported in several complementary system-dependent hypersensitivities (76).
Lectins are the other important soluble molecules which recognize viral carbohydrates and activate innate immunity by activation of complementary system and also opsonized the viral particles (31). In the previous sentences, the importance of complementary system in induction of BKAN has been discussed, so, it seems that lectins also participate in the pathogenesis of innate immunity during BKAN through activation of a complementary system. Mannose-binding lectin (MBL) is the most known lectin, which recognizes the latest units of prokaryotic carbohydrates (mannose and fructose) and activates the complementary system and also plays as mediator of phagocytosis by binding prokaryotic antigens or virions to the MBL receptors on the phagocytic cell-like macrophages (82).
Some studies have evaluated the roles of lectins in response to BK virus and the pathogenesis of BKAN. Liman et al. demonstrated that there is no association between MBL deficiency and increased risk for BKAN (51). ULBP3 is a cell surface lectin which is upregulated by under stress cells like viral infected cells. As it has been discussed in the previous section, BK virus suppresses expression of the molecules by production of miRNAs, which target the coding mRNA (10). On the other hand, a study revealed that the A allele in MBL gene was associated with better immune responses against BK virus infection (19).
Due to the studies, it may be concluded that lectins, especially MBL, play key roles against BK virus infection and the virus targets lectins to survival, but their roles in the induction of BKAN is controversial and more studies can shed light on to clear the roles of lectins in the pathogenesis of BKAN. Based on our search in the main databases, there were no studies regarding the roles of pentraxins and ficolins in response against BK virus infection and during BKAN, but there were several studies regarding the status and roles played by innate immunity cytokines and chemokines during BK virus infection and its complications, BKAN, which are discussed in the next sentences.
Cytokines are the small glycoproteins produced in both immune systems, adaptive and innate immunity (54). The cytokines produced by innate immunity cells are categorized as innate immunity cytokines. They target either innate or adaptive immunity cells (8). The most important innate immunity cytokines are tumor necrosis factor-α (TNF-α), IL-1, IL-6, IL-8, IL-10, IL-12, IL-15, and IL-18 (8). IL-10 and tumor growth factor (TGF-β) are the cytokines which are produced by both adaptive and innate immunity cells (43,46). IL-10 and TGF-β are anti-inflammatory cytokines, while other cytokines induce inflammation in the local or systemic manners.
Some studies investigated the status of the cytokines and their plausible roles in response against BK virus or reactivation of the virus and induction of BKAN. For example, a study by Corallini et al. revealed that BK virus-transformed endothelial cells express TGF-β m-RNA more than normal murine endothelial cells (21). It has also been revealed that the stimulation of peripheral blood mononuclear cells (PBMCs), from patients with a history of BK virus infection, with peptides derived from BK virus led to the production of IL-10 and TGF-β (70). Another study showed that TGF-β increased the promoter activity of BK virus through effect on a small segment of the promoter, which is essential for stimulation in response to TGF-β (1). Based on the investigations, it may be hypothesized that TGF-β may participate in the reactivation of BK virus.
On the other hand, IL-12 is a proinflammatory cytokine that is produced by activated macrophages and DCs (23). IL-12 is the most important link between adaptive and innate immunity (23). It is an important cytokine for activation of several mechanisms against viral infections, including upregulation of costimulatory molecules, MHCs, and proinflammatory cytokines, such as IFN-γ. Non-interestingly, BK virus antigens induce expression of IL-12 by DCs and macrophages (20,32), while its roles in BKAN is yet to be clarified.
According to the key roles played by IL-12 in activation of adaptive immunity, including cytotoxic T cells (CTLs), it may be hypothesized that the cytokine may induce lysis of infected cells by CTLs in indirect format. Another study revealed that IFN-γ inhibits BK virus reactivation through upregulation of IL-12 in a positive feedback (2). IL-15, as a proinflammatory cytokine, is urgently needed for the development of NK cells. NK cells are the most important innate immunity cells against viral infections (71). BK virus infection in endothelial cells results in the downregulation of IL-15 (79), which may be a main immune escape mechanism applied by BK virus.
TNF-α is another proinflammatory cytokine and immunologists believe that it is the first cytokine that is produced in the site of infection (4). It induces expression of addressing molecules like selectins, integrins, and chemokines on the endothelial cells of blood vessels of the infected site as well as neutrophil release from bone morrow (4). TNF-α also significantly participates in several immunological disorders such as infectious shock (4). Previous studies have controversial results regarding the roles of TNF-α in defense against BK virus. For instance, Heutinck et al. reported that TNF-α is an important cytokine to orchestrate the control of BK virus infection during kidney transplantation (39). Another study showed that adaptive immunity cells (T helper cells) produced TNF-α in response to BK virus antigens (86). It has also been demonstrated that BK virus-like protein is able to induce expression of TNF-α by DCs (48).
In contrast to the studies, Wilson et al. revealed that TNF-α did not contribute to the control of mouse polyomavirus infection (80). Investigations also revealed that urine levels of secreted IL (sIL)-6R, IL-6, and sIL-1 receptor A (sIL-1RA) were increased in BK virus-infected patients in comparison to BK virus-negative patients with renal transplantation (29). Another study reported that BK virus-infected renal transplant recipients showed increased urine sIL-1RA, IL-6, and sIL-6R (3). A study by Sadeghi et al. revealed that sIL-1RA in the urine was increased during BLAN, which is a marker of inflammation (69).
According to the results presented here, it may be concluded that the proinflammatory cytokines not only are essential for inhibition of BK virus reactivation, but also participate in the pathogenesis of BNAK. They can also be considered as markers of BNAK during BK virus reactivation. BK virus also induces anti-inflammatory cytokines for either immune response suppression or increased expression of its proteins through interaction of the cytokine intracellular pathway molecules with BK virus gene promoters.
Conclusion
Based on the information collected in this review article, it may be concluded that although innate immunity significantly participates in defense against BK viruses, its roles in the induction of BKAN is also crucial. It appears that innate immunity responses against BK virus may lead to killing the infected cells and it may be a reason of kidney dysfunction during BK virus reactivation. Additionally, activation of some innate immunity-related transcription factor is another reason for reactivation of lytic phase of BK virus. Furthermore, it may be hypothesized that killing of infected cells by innate immunity may lead to the release of viral and host antigens and it may result in the induction of inflammation, which is an important part of BKAN.
Accordingly, it seems that new treatment strategies such as depletion of antineutrophil cytoplasmic antibody, upregulation of ULBP3, and using anti-TGF-β can be considered for the treatment of BK virus reactivation. Additionally, based on the roles played by innate immunity receptors, like TLR3, it may be hypothesized that using the receptor antagonists can be considered as future treatment strategies, although, it needs accurate animal/clinical trial studies using specific receptor antagonists and evaluation of their plausible side effects. Moreover, new molecular strategies for targeting proinflammatory molecules like using of micro-RNAs may be applicable to the treatment of BKAN.
Footnotes
Author Disclosure Statement
No competing financial interests exist.