
Abstract
Vitamin A deficiencies are common throughout the world and have a significant negative influence on immune protection against viral infections. Mouse models demonstrate that the production of IgA, a first line of defense against viruses at mucosal sites, is inhibited in the context of vitamin A deficiency. In vitro, the addition of vitamin A to activated B cells can enhance IgA expression, but downregulate IgE. Previous reports have demonstrated that vitamin A modifies cytokine patterns, and in so doing may influence antibody isotype expression by an indirect mechanism. However, we have now discovered hundreds of potential response elements among Sμ, Sɛ, and Sα switch sites within immunoglobulin heavy chain loci. These hotspots appear in both mouse and human loci and include targets for vitamin receptors and related proteins (e.g., estrogen receptors) in the nuclear receptor superfamily. Full response elements with direct repeats are relatively infrequent or absent in Sγ regions although half-sites are present. Based on these results, we pose a hypothesis that nuclear receptors have a direct effect on the immunoglobulin heavy chain class switch recombination event. We propose that vitamin A may alter S site accessibility to activation-induced deaminase and nonhomologous end-joining machinery, thereby influencing the isotype switch, antibody production, and protection against viral infections at mucosal sites.
V
The upregulation of cytokines is one method by which vitamins influence antibody isotype expression patterns (21,29,32,35 –37). For example, vitamin A can upregulate interleukin 6 (IL-6), a cytokine that supports maturation and stabilization of IgA-producing cells (29). However, we now hypothesize that vitamins may also have a direct influence on the B cell heavy chain class switch recombination (CSR) event. The CSR involves deletion of intervening sequences within the immunoglobulin heavy chain locus between a target S (e.g., Sα) and an upstream S (e.g., Sμ or Sγ2b) region during B cell maturation. S regions are each composed of a tandem array of repeating elements, and no two regions are alike (10). Coincident with the CSR is an increase of germline transcripts, initiated at promoters upstream of the targeted S and CH genes. Germline transcripts can form hybrids with DNA, visualized as R-loops (45). During the transcription process, RNA polymerase II molecules pause in S regions and proteins associated with paused polymerases can recruit activation-induced deaminase (AID) (2,24,27,39,44). Typically, AID drives the CSR process by converting cytosine to uracil in DNA (17,25). A potential next step is for uracil DNA glycosylase to excise uracil (26), rendering the abasic site vulnerable to cuts by apyrimidinic/apurinic endonucleases (33), followed by double-strand breaks and nonhomologous end joining (3).
To determine if potential response elements exist in the heavy chain locus, we searched for eight different RARE-like half-sites within published murine S regions, including direct and reverse complement sequences. As shown in Figure 1A, our search revealed an extraordinary number of juxtaposed half-sites in the Sα region. The most frequent distances between half-sites were 4 and 9 nucleotides (DR4 and DR9), the former most typical of heterodimeric, thyroid hormone, or liver X receptor (TR-RXR; LXR-RXR) binding motifs (38). Like Sα, Sμ, and Sɛ regions exhibited high-density response elements (Supplementary Figs. S1 and S2; Supplementary Data are available online at

Response element half-sites in the mouse Sα and Sγ2b regions. Response element half-sites are shown by color coding in the Sα
In a separate analysis, we examined human S regions. As was the case for mouse S regions, the frequency of juxtaposed half-sites was very high in human Sα (Sα1 and Sα2), Sμ, and Sɛ with numerous DR4 and DR9 patterns, but juxtaposed half-sites in Sγ sequences were relatively rare or absent (Supplementary Figs. S6–S13).
Because the half-site TGAGCT (reverse complement AGCTCA) was especially frequent, a full scan for this sequence in mouse and human heavy chain loci was conducted. Figure 2 illustrates the high density of AGCTCA sites in Sμ, Sα, and Sɛ regions for both human and mouse loci, and shows that when half-sites were observed in Sγ regions, they were often oriented as the reverse complement. Results support a hypothesis that retinoid receptors directly bind Sμ, Sα, and Sɛ regions and thereby promote or inhibit Sμ-Sα and Sμ-Sɛ joins. The half-sites in Sγ regions are rarely adjacent to one another, atypical of consensus RARE motifs, but might nonetheless support weak binding of nuclear receptors or binding of related proteins.
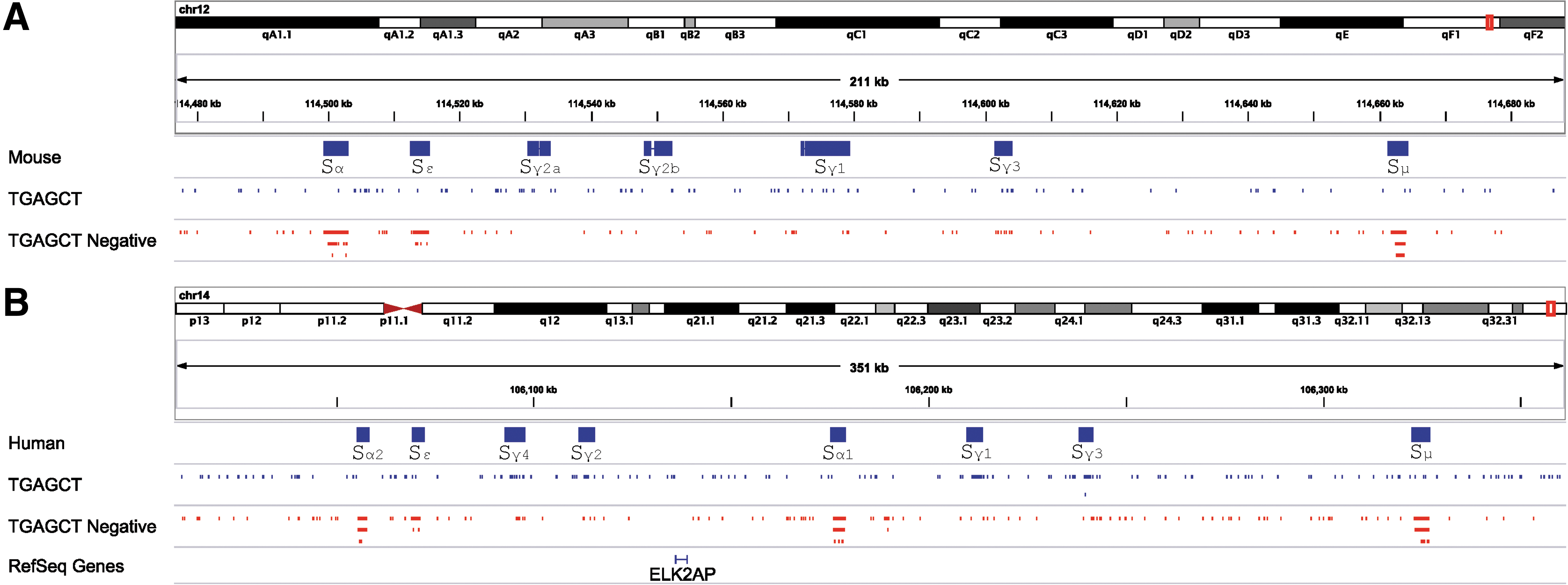
Clustered response elements in mouse and human heavy chain loci. Mouse and human immunoglobulin heavy chain switch regions were collected from NCBI Nucleotide database and aligned to mouse assembly mm9(NCBI37) or human assembly hg19(GRCh37) separately with BLAT (version 34 default parameters with maximum intron size 10 kb). We retained the best score alignment for each sequence. Alignment positions were loaded into Integrative Genomics Viewer (IGV, version 2.3). “TGAGCT” motif-matched tracks were created using the “find motif” tool in the IGV (28). Alignment positions are shown in
To seek potential binding sites for other nuclear receptors, we asked if potential estrogen response elements (ERE) were frequent in S regions of heavy chain loci. RARE and ERE can be juxtaposed to mediate multihormonal responsiveness in gene regulation, and estrogen is known to influence AID and CSR (12,16,23). A canonical DNA motif for the estrogen receptor is a palindrome with half-sites GGTCA and TGACC separated by three nucleotides (11). To allow for sequence variability, we searched for G/A G/A T/C T/C G/A NNN TGA C/G C in S regions. In so doing, we found that high-density motifs appeared in Sμ and Sα regions of human and mouse loci, but not in Sγ regions. For example, there were more than 70 motifs in Sα regions for both human and mouse, yet not a single motif in Sγ2 regions in either species.
Altogether, data encourage investigation of potential interactions between nuclear receptors and response element hotspots in immunoglobulin heavy chain S region loci. We note that the response elements observed in this study were positioned in S regions rather than in the known promoter regions for sterile transcripts. We, therefore, propose that nuclear receptors may influence CSR by mechanisms other than transcription upregulation (31). Possibly, the tripartite binding of ligands (e.g., vitamins), nuclear receptors, and DNA, alter chromatin and the subsequent recruitment of enzymes (e.g., HDAC3, and AID) to enhance deamination and strand breaks in S regions. We note that a hotspot for AID binding is AGCT (8), a sequence within the half-site TGAGCT, supportive of our hypothesis. Should nuclear receptors bind S regions and recruit AID, they would not act in isolation. Previous research has shown that S regions are bound by diverse macromolecular complexes comprising a plethora of factors (e.g., RNA polymerase II, SPT5, RNA exosome, 14-3-3 adapters, AID, E-box-binding proteins, NFκB, and histone-modifying enzymes), each of which may influence isotype expression patterns by activated B cell populations (2,14,24,39,40,43,44).
In conclusion, our results reveal a large number of potential response elements for vitamins and hormones in immunoglobulin S loci. Future research may determine if and how nuclear receptors bind (or are prohibited from binding) these regions. Results should provide a better understanding of CSR mechanisms, isotype expression patterns, and strategies with which protection against infectious diseases may be improved.
Footnotes
Acknowledgments
The research was funded in part by NIH NIAID R01 AI088729, NCI P30 CA21765, the American Lebanese Syrian Associated Charities (ALSAC), and the Intramural Research Program of the NIH, National Institute on Aging.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.